-fr.svg.png.webp)
L'appareil de Golgi est un organite des cellules eucaryotes[1].
Il est un lieu majeur de transfert et de tri des molécules, ainsi que de synthèse des glycoprotéines et des sphingolipides.
Par sa position, entre le réticulum endoplasmique et la membrane plasmique, et sa fonction, il est l'intermédiaire entre la maturation et la sécrétion des protéines élaborées dans le réticulum, il régule le transport vésiculaire et poursuit et finalise les modifications post-traductionnelles des protéines comme les glycosylations, sulfatations, phosphorylations.
Il est composé de deux faces : la face cis, face d'entrée des protéines sécrétées par le réticulum et la face trans, d'où bourgeonnent les vésicules en direction des lysosomes, du milieu extracellulaire (exocytose) ou de la membrane cytoplasmique. Ainsi on note deux types de vésicules formées par les coatmères : les COPI (impliqués dans le transport rétrograde depuis le réseau Trans_Golgi vers le réseau Cis_Golgi, et du réseau Cis_Golgi vers le Réticulum endoplasmique), et les COPII (impliqués dans le transport antérograde des protéines du Réticulum endoplasmique rugueux vers le Cis_Golgi). On note aussi des vésicules à clathrine formées de l'association d'adaptines et de triskélions. Ces vésicules sont impliqués dans les processus d'endocytose, dans le transport des protéines depuis l'appareil de Golgi vers les lysosomes ou vers les vésicules de sécrétion.
Découverte
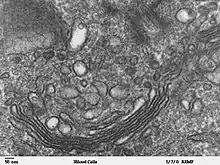
L'appareil de Golgi fut décrit par Camillo Golgi en 1898. Il observait alors un arrangement particulier de l'appareil réticulé interne. Il nomma cet arrangement en forme de croissant le dictyosome, qui sera plus tard rebaptisé à son nom, en raison de l'importance de ses travaux. Ses recherches à l'époque ont soulevé beaucoup de controverses. La coloration qu'il utilisait pour voir l'appareil réticulé était la coloration à l'argent. Cette méthode peu spécifique laissait croire à la communauté scientifique que les dictyosomes étaient simplement des artefacts.
Composition et structure
Structure
L'appareil de Golgi est un organite intracellulaire polymorphe constitué d'un ou plusieurs dictyosomes[2], de vésicules et de canalicules. Chez l'homme il en contient de 3 à 10. Ainsi, dans certaines cellules spécialisées (cellules sécrétrices), l'appareil de golgi contient de nombreux dictyosomes.
Un dictyosome est un empilement de saccules membranaires en forme d'assiette épaisse.
L'appareil de Golgi est formé d'un empilement de vésicules aplaties. Chaque dictyosome peut être divisé en trois régions fonctionnelles différentes :
- les saccules de la face cis ou face d'entrée. C'est ici qu'arrivent les vésicules en provenance du réticulum endoplasmique ;
- les saccules de la région médiane ;
- les saccules de la face trans ou face de sortie, où les vésicules destinées à la sécrétion ou à l'exocytose terminent leur parcours.
Composition
- Pour la membrane golgienne : les protéines constituent entre 60 et 65 % et les lipides de 35 à 40 %. C'est une composition intermédiaire entre le RE et la membrane plasmique riche en glycosyl transferase.
La répartition des protéines et l'activité enzymatique au sein de l'appareil de golgi est diverse :
Ainsi, les saccules trans sont riches en nucléoside diphosphatase, alors que le réseau trans est riche en phosphatase acide.
Les différentes parties de l'appareil de Golgi sont identifiées par des colorants
- le tétroxyde d'osmium colore les saccules cis ;
La méthode de la coloration immunohistochimique est également utilisée, durant laquelle on fait réagir par immunomarquage ou par fluorescence des anticorps ou des substrats spécifiques de certaines protéines :
- la dinucléoside diphosphatase dans les saccules trans ;
- la phosphatase acide dans le réseau trans golgien.
Fonctionnement
L'appareil de Golgi est le lieu où certaines protéines sont modifiées, notamment par glycosylation, après leur synthèse dans le réticulum au cours de la traduction des molécules d'ARNm- et de l'assemblage des protéoglycanes.
L'évolution des composés contenus dans le dictyosome s'effectue de la face cis (cis-golgi, contenant 70 % de protéines et 30 % de lipides) à la face trans (trans-golgi, contenant 60 % de protéines et 40 % de lipides) de celui-ci. Les protéines qui composent chacune des régions sont différentes et subissent des transformations différentes dans chaque compartiment. Il existe deux postulats quant au transport des protéines dans l'appareil de Golgi :
- Le modèle du transport vésiculaire : l'appareil de Golgi serait statique et le transport des protéines dans celui-ci se ferait par des vésicules. Les enzymes spécifiques de chaque compartiment seraient donc maintenues sur place alors que les protéines voyageraient via les vésicules qui bourgeonneraient d'un compartiment pour aller fusionner avec le suivant. Il existerait tout de même un flux rétrograde de vésicules qui s'assurerait de ramener toutes les protéines qui ont été capturées malencontreusement dans les vésicules COP I se dirigeant vers la face cis.
- Le modèle de la maturation des citernes : le Golgi serait une structure dynamique où les différents compartiments se transformeraient au fur et à mesure et migreraient progressivement vers la face trans. Ainsi, les vésicules du RER fusionneraient pour donner le premier compartiment (face cis) qui se transformerait à son tour au fur et à mesure qu'il avance pour devenir un compartiment médian et ainsi de suite jusqu'à se dissocier en diverses vésicules. Là aussi, un flux rétrograde de vésicules assurerait la spécificité de chaque compartiment en ramenant les enzymes spécifiques du compartiment qui vient de migrer.
Les vésicules produites par l'appareil de Golgi permettent l'assemblage des chaînes lourdes (H) et des chaînes légères (L) (dans le cas des anticorps) ainsi que l' "emballage" des immunoglobulines. De plus, ces vésicules de sécrétion permettent les transports des protéines vers la membrane cytoplasmique.
Fonction
L'un des rôles majeurs de l'appareil de Golgi est lié à des phénomènes d'exocytose. Il est le point de passage obligé et régulateur du trafic vésiculaire. Il régule le nombre de vésicules allant à la membrane et participe au renouvellement membranaire. Il entraîne des modifications post-traductionnelles des protéines :
- clivage des précurseurs polypeptidiques : maturation des protéines ;
- glycosylation (ajout de chaînes glucidiques) ;
- sulfatation ;
- phosphorylation ;
- ajout de chaîne d'acide gras.
Des modifications post-traductionnelles effectuées dans l'appareil de Golgi sont essentielles à l'adressage correct des protéines dans la cellule. Par exemple, dans le cis-golgi, il y a phosphorylation de certains résidus mannose de chaines oligosaccharidiques liées en N- sur les protéines qui aboutit à la présence de mannose-6-phosphate (il s'agit notamment des hydrolases lysosomales). Dans le trans-golgi, des récepteurs au mannose-6-phosphate concentrent les protéines à mannose-6-phosphate dans des vésicules spécifiques qui sont ensuite adressées aux lysosomes. Les protéines adressés aux lysosomes sont souvent des enzymes lytiques (hydrolases) dangereuses pour la cellule et son environnement, ainsi leur "signalement" par le mannose-6-phosphate dans l'appareil de Golgi évite qu'elles soient excrétées et détruisent les tissus.
Sécrétion des protéines
L'appareil de Golgi assure un contrôle précis sur les protéines qui le quittent. D'abord, il s'assure que les protéines nécessaires au fonctionnement du réticulum endoplasmique seront retournées par un flux rétrograde vers ce dernier. Ces protéines sont marquées par la séquence KDEL ou KKXX selon leur fonction. Le KDEL correspond aux protéines solubles et le KKXX aux protéines membranaires. Certaines protéines comportant la séquence KKXX servent de récepteurs au KDEL et assurent ainsi le flux rétrograde des protéines à KDEL de l'appareil de Golgi vers le réticulum endoplasmique.
Ensuite, il existe deux types de sécrétions pour les protéines dans l'appareil de Golgi. La sécrétion constitutive est généralement dirigée vers la membrane plasmique. Cette sécrétion ne nécessite aucun signal. La sécrétion contrôlée, comme son nom l'indique, nécessite un signal. Ce principe permet de contrôler l'exportation de ces protéines.
Transport golgien
Le transport antérograde des protéines du réticulum endoplasmique vers l'appareil de Golgi s'effectue par l'intermédiaire de vésicules dont le bourgeonnement s'effectue grâce aux protéine
2. Les transports transgolgiens, c'est-à-dire entre les saccules de l'appareil de Golgi, et les transports rétrogrades des vésicules de l'appareil de Golgi vers le réticulum endoplasmique, sont réalisés grâce aux vésicules qui bourgeonnent par l'intermédiaire protéines COP 1. Ces protéines sont des protéines de manteau (coat protein), et n'ont donc pas de rôle dans l'adressage ou le transport des vésicules, mais sont essentielles au bourgeonnement des vésicules au niveau de la membrane de l'appareil de Golgi. Le transport et surtout l'adressage de ces vésicules est assuré par les protéines de la famille SNARE. La sécrétion continue utilise des vésicules de FAPP, la sécrétion vers les rafts lipidiques elle, utilise des vésicules à cavéoline, tandis que la sécrétion régulée se fait par l'intermédiaire de vésicules recouvertes de clathrine.
Notes et références
Articles connexes
- Vacuome