Les cônes sont des photorécepteurs situés dans la rétine, transformant le signal électromagnétique de la lumière en signal nerveux permettant la vision photopique diurne. La vision scotopique nocturne est assurée par les bâtonnets.
Description
Distribution anatomique sur la rétine humaine
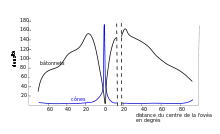
Les cônes sont entre 3[1] et 4[2] millions par œil chez l'Homme. Ils ne représentent que 5 % du total des photorécepteurs et sont principalement concentrés sur la fovéa, au centre de la rétine, dans le prolongement de l'axe optique. La partie centrale de la fovéa (ou « fovéola »), sur un rayon de 0,3 mm, ne contient que des cônes[3].
Cette région centrale est dotée d'une grande acuité visuelle. Chaque cône n'y est connecté qu'à une cellule bipolaire, elle-même liée à une seule cellule ganglionnaire. Cette région toute petite se projette ensuite dans le cortex sur une aire mille fois plus grande. Dès qu'on s'éloigne du centre, la densité des cônes diminue très rapidement, le degré de convergence avec les cellules ganglionnaires augmente et l'acuité s'en trouve corrélativement fortement réduite[4].
Cette propriété de restriction de l'acuité à une petite région rétinienne oblige à bouger des yeux sans arrêt pour percevoir clairement les objets intéressants.
Les deux, trois ou quatre types de cônes (selon l'espèce)

L'humain perçoit une immense variété de couleurs différentes. Pourtant il ne possède que trois types de cônes ayant une sensibilité plus grande à certaines radiations de longueurs d'onde comprises entre 400 et 700 nm : les cônes (B) sensibles aux radiations de basses longueurs d'onde ou cônes cyanolabes (437 nm), les cônes (V) sensibles aux radiations de moyennes longueurs d'onde ou cônes chlorolabes (533 nm), et les cônes (R) sensibles aux radiations de grandes longueurs d'onde ou cônes érythrolabes (564 nm) (ces derniers réagissant d'ailleurs principalement aux radiations provoquant la sensation jaune[5]). Dans la littérature scientifique anglo-saxonne, l'usage est de qualifier les cônes bleus de S (pour short), les cônes verts de M (pour medium) et rouges de L (pour long) en référence à la longueur d'onde au maximum de sensibilité. Ces maxima de sensibilité sont par ailleurs différents de plusieurs nanomètres d'un individu à l'autre[3].
Chaque type de cônes est sensible à des radiations appartenant à un domaine étendu de longueur d'onde, dans la mesure où sa réponse ne fait que refléter le nombre de photons qu'il capte, indépendamment de leur longueur d'onde (le cône rouge capte aussi bien des photons verts de 500 nm, jaunes de 560 nm ou rouges de 650 nm). Un photorécepteur n'est qu'un « compteur de photons », suivant la formule de Michel Imbert[6], chaque photon absorbé par le pigment produit le même effet. La longueur d'onde n'intervient qu'au niveau de la probabilité d'absorption suivant la sensibilité spectrale du pigment[N 1]. La perception des couleurs n'est possible qu'au niveau central par comparaison des signaux issus de deux classes de cônes.
De plus, la sensibilité spectrale, très proche, des cônes V et R fait qu'ils servent principalement à détecter la structure spatiale des images.
Chez l'Homme[7], les cônes B sont les moins nombreux (4 % – 5 %); puis viennent les cônes V et les cônes R (dans un rapport variable R/V allant de 1,1 à 4), avec des variations interindividuelles importantes. Les cônes forment une mosaïque avec chaque type disposé de manière aléatoire.
Les vertébrés de type poissons, reptiles et oiseaux ont une vision plus riche en couleurs grâce à leurs quatre pigments (répondant de manière optimale aux longueurs d'onde rouge, vert, bleu et ultra-violet, cette dernière couleur intervenant notamment dans les caractères sexuels secondaires importants dans la parade nuptiale). Par rapport aux mammifères, les oiseaux par exemple voient une plus grande proportion de couleurs naturelles comme non-spectrales[8] (pour rappel, une couleur est dite non-spectrale quand « les types de cônes non adjacents (sensibles à des parties largement séparées du spectre lumineux) sont principalement stimulés ; pour les humains, le violet est une lumière non-monochromatique qui apparait quand il y a stimulation au même moment des cônes sensibles au bleu et au rouge, deux cônes ayant des courbes de sensibilité spectrale non adjacentes) est une couleur non spectrale »[8]. Une étude récente (2020) basée sur des colibris sauvages formés à participer à des tests de vision des couleurs montre qu'ils distinguent une variété de couleurs non spectrales dont UV + rouge ; UV + vert ; violet et UV + jaune[8]. Ceci améliore leur capacité de se signaler par rapport aux éventuels partenaires sexuels et autres oiseaux, tout en facilitant leur recherche de nourriture[8].
Les mammifères non primates possèdent deux types de pigments bleu et rouge, la perte de deux pigments correspondant à leur mœurs de chasseurs nocturnes insectivores au Trias. Une duplication d'un gène fabriquant un pigment rétinien puis sa mutation auraient doté un ancêtre commun aux primates d'un troisième pigment sensible au rouge, permettant à cette espèce devenue diurne et omnivore de mieux distinguer la couleur des fruits sur le fond vert des feuillages[9].
Structure des cônes
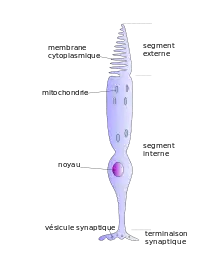
La fonction des cônes est de convertir l'énergie lumineuse en variation du potentiel électrique de membrane. Cette transduction du signal se fait dans les disques du segment externe grâce à un pigment nommé iodopsine, molécule composée d'une protéine de la classe des opsines et de rétinène (ou rétinal), un dérivé de vitamine A. Un photon, en tombant sur une molécule de rétinène, change sa conformation, en la faisant passer de l'état 11-cis en l'état tout-trans. Il s'ensuit une cascade de réactions qui, par l'intermédiaire de la stimulation d'une protéine G, aboutit à la fermeture des canaux sodiques et à l'hyperpolarisation de la membrane.
Ainsi, bizarrement, la lumière qui éclaire un cône ne produit pas sa dépolarisation mais son hyperpolarisation. À l'obscurité, le cône est dépolarisé[4], avec un potentiel de membrane d'environ −40 mV. Lorsque l'intensité lumineuse augmente, le potentiel de membrane est de plus en plus négatif, jusqu'à atteindre la saturation à −65 mV.
Ainsi, à l'obscurité, les photorécepteurs sont dépolarisés, un grand nombre de canaux calciques de la terminaison synaptique sont ouverts, entrainant une libération importante d'un neurotransmetteur, le glutamate, par la terminaison synaptique. Et lorsqu'on éclaire, l'inverse se produit : la lumière produit une diminution de la vitesse de libération des transmetteurs. Lorsque l'intensité du stimulus lumineux augmente graduellement, le taux de libération de glutamate diminue corrélativement.
Les différentes sensibilités
Chaque type de cône a une bande passante limitée, dans des longueurs d'onde correspondant à ce qui est perçu comme des couleurs. Ainsi les oiseaux et les reptiles, comme vraisemblablement les ancêtres des mammifères, distinguent quatre couleurs de base.
| Arbre phylogénétique simplifié | Type de vision | Longueurs d'onde |
|---|---|---|
|
─┬─ Tétrapode primitif |
Quadrichromie |
|
Des recherches actuelles[10],[11] tendent à prouver que chez un certain pourcentage d'hommes (10 %) et de femmes (50 %), il existerait un quatrième type de cônes sensibles aux oranges.
Maladies
Les daltoniens ont une défaillance du système liée à l'un de ces trois cônes. En effet, les cônes ne fonctionnent que grâce à la présence de « pigments photorécepteurs », sensibles à la lumière. L'absence de l'un de ces pigments provoque une défaillance du type de cône concerné (S, M ou L). Le cerveau doit donc se passer des informations envoyées par ce cône pour concevoir la vision.
L'achromatopsie congénitale est une maladie due à l'absence totale de pigments : seuls les bâtonnets fonctionnent, ne permettant à la personne qu'une vision en nuances de gris (et non en noir et blanc). Toutefois, cette maladie reste très rare, et n'affecte qu'une personne sur 33 000.
Finalement, certaines personnes peuvent souffrir d'une vision des couleurs faussée due aux faibles performances d'un pigment, qui dans leur cas détecte un spectre lumineux réduit.
Notes
- ↑ La réponse d'un cône est gouvernée par le principe d'univariance : alors que le stimulus lumineux peut varier selon deux dimensions, l'intensité et la longueur d'onde, la réponse d'un photorécepteur est limitée à une seule dimension, l'amplitude de son potentiel récepteur (sa polarisation graduée de −40 mV à −70 mV).
Références
- ↑ (en) Jonas JB, Schneider U, Naumann GO, « Count and density of human retinal photoreceptors », Graefes Arch Clin Exp Ophthalmol, vol. 230, no 6, , p. 505-10. (PMID 1427131).
- ↑ (en) Curcio CA, Sloan KR, Kalina RE, Hendrickson AE, « Human photoreceptor topography », J Comp Neurol, vol. 292, no 4, , p. 497-523. (PMID 2324310).
- 1 2 (en) Arne Valberg, Light Vision Color, Wiley, , 462 p..
- 1 2 Dale Purves, G-J Augustine, D. Fitzpatrick, W-C Hall, LaManta, McNamara, Williams, Neurosciences, De Boeck, , 811 p..
- ↑ On les appelle tout de même ainsi du fait que les longueurs d'onde rouges activent ces cônes sans activer les cônes verts.
- ↑ Michel Imbert, « La connaissance de la couleur », dans M. Elias, J. Lafait, La couleur : lumière, vision et matériaux, Belin, .
- ↑ Austin Roorda, Andrew B. Metha, Peter Lennie, David R. Williams, « Packing arrangement of the three cone classes in primate retina », Vision Research, vol. 41, , p. 1291-1306.
- 1 2 3 4 (en) Mary Caswell Stoddard, Harold N. Eyster, Benedict G. Hogan et Dylan H. Morris, « Wild hummingbirds discriminate nonspectral colors », Proceedings of the National Academy of Sciences, vol. 117, no 26, , p. 15112–15122 (ISSN 0027-8424 et 1091-6490, PMID 32541035, PMCID PMC7334476, DOI 10.1073/pnas.1919377117, lire en ligne, consulté le )
- ↑ (en) M. D. Eaton, « Human vision fails to distinguish widespread sexual dichromatism among sexually "monochromatic" birds », Proceedings of the National Academy of Sciences, vol. 102, no 31, , p. 10942-10946 (DOI 10.1073/pnas.0501891102).
- ↑ Backhaus, Kliegl & Werner « Color vision, perspectives from different disciplines » (De Gruyter, 1998), p.115-116, section 5.5.
- ↑ Pr. Mollon (université de Cambridge), Pr. Jordan (université de Newcastle) « Study of women heterozygous for colour difficiency » (Vision Research, 1993).