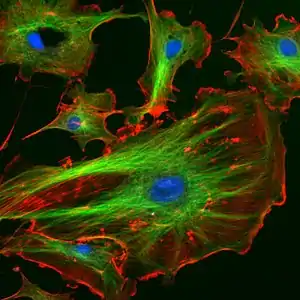
Le cytosquelette d'une cellule est l'ensemble organisé des polymères biologiques qui lui confèrent l'essentiel de ses propriétés architecturales et mécaniques. La référence terminologique au « squelette » des vertébrés est cependant trompeuse puisque :
- la plupart des composantes du cytosquelette sont renouvelées par polymérisation en permanence ;
- le cytosquelette est à l'origine de la plupart des forces exercées par la cellule pour se déplacer et se nourrir, ce en quoi il s'apparente plutôt à un ensemble de « muscles » ;
- les propriétés mécaniques du cytosquelette sont très variables suivant les composantes et les situations considérées ;
- l'activité du cytosquelette a des conséquences dépassant de loin les rôles purement mécaniques (mouvement et structure), par exemple : la régulation de l'expression génique, la différenciation cellulaire ou la prolifération.
Les cytosquelettes de tous les eucaryotes sont assez similaires (bien que des différences importantes existent entre les cellules animales et végétales), tandis que ceux récemment découverts chez les procaryotes semblent organisés de façon tout à fait différente.
Cytosquelette des eucaryotes
Composition et structure globale
Le cytosquelette est constitué de polymères biologiques de protéines, qu'on qualifie parfois de fibres étant donné leur taille importante à l'échelle cellulaire. On les classe en trois catégories :
- les filaments d'actine, formés d'actine (protéine dont il existe différents types). On retrouve aussi ces mêmes filaments en grande quantité dans les fibres musculaires. Leur diamètre avoisine 7 à 8 nm, et leur longueur de persistance d'environ 17 µm. Ce sont donc des filaments assez flexibles, puisqu'ils sont disposés en double hélice. Ils sont également orientés, du fait de l'asymétrie des monomères d'actine et de leur assemblage en hélice : l'une des extrémités (+) peut polymériser beaucoup plus vite que l'autre (-). On parle de filaments polarisés. Les filaments d'actine forment un réseau dynamique. La dynamique des filaments d'actine est permise par un phénomène de tapis roulant (dépolymérisation à l'extrémité (-) et polymérisation à l'extrémité (+) et par le clivage des filaments par certaines protéines (raccourcissant de ce fait les filaments et multipliant par deux les sites (+) de polymérisation).
- L'actine est associée à d'autres protéines (protéine de réticulation) comme la filamine et la fodrine, soit des protéines se chiffrant à 250 et 243 kDa respectivement, qui sont à l'origine du réseau tridimensionnel. De surcroît, la fodrine peut être à l'origine de l'apparition de maladies auto-immunes où la production d'auto-anticorps ciblant cette protéine est extrapolée chez les individus souffrant de la maladie de Sjogren. Le résultat de ces associations est, entre autres, l'augmentation de la viscosité du cytosol et le déclenchement de certaines voies signalétiques. Les filaments d'actines sont composés du polymère d'actine F (filament) et du monomère d'actine G (globulaire). Il existe un équilibre dynamique entre l'actine F et l'actine G. Le réseau d'actine est situé sous la membrane plasmique, le degré de polymérisation définit la forme globale de la cellule et la plasticité de la cellule qui est nécessaire pour les processus de migration, d'endocytose, de division, etc. ;
- les filaments intermédiaires. Ce sont les éléments les moins dynamiques du cytosquelette, mais leur étude se développe rapidement. Ils sont très importants pour la structure du noyau puisqu'ils sont les plus résistants. Eux sont non polarisés. Ils permettent l'ancrage des organites. Ils ont un diamètre pouvant se situer entre 8 et 10 nm, leur donnant ainsi une taille intermédiaire entre les microfilaments d'actine et les microtubules. On les trouve dans toutes les cellules eucaryotes mais pour certaines, on ne les trouve que chez les vertébrés. On distingue :
- les filaments à kératine qui sont caractérisés par de nombreux ponts disulfures ; on les trouve dans les cellules épidermiques des vertébrés, les cheveux, les poils, les ongles, etc.,
- les filaments à desmine qu'on retrouve dans les cellules musculaires des muscles lisses, striés et dans le muscle cardiaque,
- la lamina nucléaire présente dans le noyau appliquée contre la membrane interne du noyau ; c'est une couche protéique fibrillaire dont les protéines sont des lamines ;
- les microtubules sont les constituants les plus rigides du cytosquelette. Ce sont des structures cylindriques creuses dont la paroi est constituée de polymères de tubuline (dimères de tubuline α et β). Leur longueur de persistance est en effet de plusieurs millimètres, ce qui dépasse largement l'échelle de la cellule, pour un diamètre variant de 15 à 25 nm suivant les types de microtubules. Cette rigidité leur est conférée par une structure en tube due à l'assemblage particulier des monomères qui les composent. Les microtubules sont polarisés de la même façon que les filaments d'actine, mais la biochimie de polymérisation est différente. En particulier, il existe une instabilité dynamique qui peut conduire à un raccourcissement très brutal d'un microtubule, ce qui peut être à l'origine d'une force importante.
Les polymères sont organisés en réseaux, faisceaux ou câbles suivant les rôles qu'ils remplissent. Il est important de garder à l'esprit que la structure et la composition du cytosquelette sont hautement dynamiques. Ce haut niveau d'organisation et de labilité est rendu possible par la présence de centaines de protéines auxiliaires associées au cytosquelette :
- les protéines de pontage (couramment désignées par les termes anglais crosslinkers ou crosslinking proteins). Ce terme fait référence à la physique des polymères, où l'ajout de certains composants (comme le soufre pour le caoutchouc) conduit à la formation de pontages entre les chaînes et modifie complètement les propriétés physiques du matériau. C'est effectivement ce qui se passe, de façon encore plus spectaculaire, dans le cas du cytosquelette. En effet, la plupart des crosslinkers sont contrôlés par la cellule au moyen d'autres protéines de régulation, ce qui permet des réorganisations parfois très rapides du cytosquelette ;
- les protéines de branchement (parfois considérées comme un cas particulier des précédentes, mais importantes essentiellement dans le cas des filaments d'actine) ;
- les protéines capuchon et les protéines dépolymérisantes (capping, severing proteins) qui régulent les taux de polymérisation des filaments à leurs extrémités ;
- les protéines d'ancrage ;
- les moteurs moléculaires, tels les myosines, les kinésines ou les dynéines. Vaste champ d'investigation de la biologie cellulaire, les moteurs moléculaires sont fondamentaux pour l'ajustement des propriétés mécaniques et morphologiques de la cellule ainsi que pour le trafic vésiculaire.
Cytosquelette des végétaux
La forme de la cellule étant principalement définie par l'interaction pression osmotique/résistance de la paroi, on constate une moindre importance du cytosquelette : absence de filaments intermédiaires (sauf au niveau du noyau). Le réseau de microtubules est très développé et s'organise en une structure hélicoïdale se plaquant contre la membrane (liaison avec la synthèse des fibres de cellulose de la paroi). En effet la synthèse des fibres de cellulose de la paroi végétale est orientée grâce à des microtubules corticaux le long de la membrane plasmique.
Structures localisées
Principales fonctions
Le cytosquelette contribue à de nombreuses fonctions au sein de la cellule :
- la régulation de la forme de la cellule (exemples : diapédèse, sensibilité cellulaire aux paramètres mécaniques et topographiques du substrat environnant, matrice extracellulaire, autres cellules, biomatériaux...) ;
- l'ancrage aux membranes des cellules voisines ;
- la formation de protrusions ou d'involutions membranaires (importantes pour la phagocytose et pour la migration cellulaire : pseudopodes) ;
- le maintien de la structure interne, et en particulier des compartiments cellulaires ;
- le transport de protéines ou d'ARNm ;
- la séparation des chromosomes lors de la mitose ;
- la formation et la contraction de l'anneau mitotique permettant la séparation physique de deux cellules filles (cytodiérèse) ;
- la contraction des cellules musculaires.
Cette liste ne saurait être exhaustive, de nombreux rôles inconnus étant mis en évidence régulièrement par la recherche qui est très active dans ce domaine.
Propriétés mécaniques

Les différents types de filaments ont des propriétés mécaniques très différentes :
- les microtubules ont une capacité d'étirement importante mais une très faible résistance ;
- les microfilaments d'actine sont très résistants mais leur capacité de déformation est très réduite ;
- les filaments intermédiaires se déforment presque proportionnellement à la force exercée et le recouvrement important des dimères et des tétramères permet une très grande résistance aux contraintes mécaniques.
Cytosquelette des procaryotes
La présence d'un cytosquelette a été mise en évidence chez les procaryotes en 2006, grâce notamment aux travaux de Rut Carballido Lopez[1] et de son équipe. Ceux-ci ont découvert la protéine MreB (en), homologue à la protéine d'actine, et de structure similaire, localisée sous la membrane et semblant jouer un rôle important dans la structure et la forme cellulaire. La protéine FtsZ jouerait également un rôle dans la cytodiérèse des bactéries.
Notes et références
- ↑ (en) Rut Carballido-López, « The Bacterial Actin-Like Cytoskeleton », Microbiology and Molecular Biology Reviews, vol. 70, no 4, , p. 888-909 (DOI 10.1128/MMBR.00014-06)
Voir aussi
Articles connexes
- Complexe Arp2/3