(P1) le renversement de l'image par le système optique de l’œil dans le sens haut/bas et gauche/droite
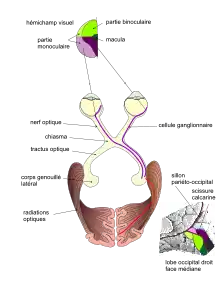
(P2) au niveau du chiasma, le croisement des fibres issues de l'hémirétine nasale et la projection du même côté des fibres issues de l'hémirétine temporale
(P3) au niveau des radiations optiques, les fibres représentant le quadrant supérieur arrivent sur la lèvre inférieure de la scissure calcarine (et inversement)
(P4) la fovéa, au centre du champ visuel, se projette dans la partie postérieure du cortex strié. La partie binoculaire de chaque hémichamp visuel se projette dans la région intermédiaire du cortex strié. La partie temporale, monoculaire de chaque champ visuel se projette sur la partie antérieure du cortex visuel.
Le système visuel humain est l'ensemble des organes participant à la perception visuelle humaine, de la rétine au système sensori-moteur. Son rôle est de percevoir et d'interpréter deux images en deux dimensions en une image en trois dimensions. Il est principalement constitué de l'œil (et plus particulièrement la rétine), des nerfs optiques, du chiasma optique, du tractus optique, du corps genouillé latéral, des radiations optiques et du cortex visuel.
Vision et nerf optique
En première approximation, l'œil peut être assimilé à un appareil photographique. La lumière diaphragmée par la pupille est déviée par la cornée et le cristallin puis projetée sur la rétine. L'image ainsi formée est une perspective conique inversée de l'espace environnant.
Il y a ensuite, au niveau de la rétine, transduction de l'information lumineuse en potentiels d'actions. L'image est analysée au sein même de la rétine en zones de contraste et le résultat de ce traitement est envoyé au reste du système visuel par le nerf optique. Chez les humains, le système visuel est le seul système sensoriel à être directement connecté, via le nerf optique, au cerveau, dû à la nécessité de traiter rapidement l'information visuelle.
La lumière pénètre dans l'œil, passant à travers la cornée et la pupille (contrôlée par l'iris) et est réfractée par le cristallin. Le cristallin inverse l'image qu'il projette sur la rétine.
La rétine contient un grand nombre de cellules « réceptrices » ainsi qu'une molécule nommée rhodopsine.
Deux voies visuelles, neuro-physiologiquement distinctes
Dans le cerveau humain, coexistent deux systèmes visuels transportant tous deux de l'information provenant de notre environnement, de la rétine vers corps genouillé latéral puis au cortex visuel primaire[1],[2],.
- la voie du système magno-cellulaire. Il est spécialisé dans trois types d'informations : le mouvement (on parle de « système analyseur de flux » visuel), la profondeur (liée à la vision 3D binoculaire) et la forme globale. Il retient les fréquences spatiales basses et mobilise les celules de la rétine périphérique (qui sont également plus sensibles que les autres dans la pénombre). Cette voie se prolonge ensuite par la « voie visuelle dorsale », qui répartit les informations visuelles ainsi acquises à plusieurs structures corticales et sous-corticales qui lui sont fortement connextées (cortex temporal, pariétal, pré-frontal et frontal, cervelet, structures thalamiques mésencéphaliques et pontiques)[3].
- la voie du système parvo-cellulaire (on parle de « système analyseur d’images »), qui véhicule les signaux permettant au cerveau de distinguer les couleurs et d'apprécier le détail des formes et des textures (fréquences spatiales hautes) ; cette voie mobilise les cellules du centre de la rétine. On a montré (sur le modèle animal) qu'à la naissance vette voie est moins mûre que celle du système analyseur de flux[3]. Cependant, lors des premiers mois de vie, ces deux voies améliorent ensuite rapidement leur performance[3].
Les informations transportées séparément par ces deux systèmes sont ensuite recombinées par le cerveau (maladroitement les premières années de la vie) puis permettent la perception spatiotemporelle necesaire à une planification de plus en plus sûre des actes[4].
Chiasma et tractus optique
Les nerfs optiques de chaque œil se rejoignent au niveau du chiasma optique, ce qui permet de redistribuer l'information visuelle selon qu'elle est dans la partie droite ou dans la partie gauche du champ visuel (chaque œil voit à gauche et à droite en même temps). L'hémichamp gauche va dans l'hémisphère droit et vice-versa (il y a une inversion).
La continuation des nerfs optiques après le chiasma s'appelle le tractus optique (en). Dans chaque hémisphère, il distribue l'information visuelle à différents noyaux :
- le corps géniculé latéral. Ce dernier envoie l'information qui va être analysée par le cortex visuel pour donner notamment la perception visuelle consciente ;
- le noyau suprachiasmatique de l'hypothalamus. Ce noyau contient l'horloge biologique. L'information de la présence ou de l'absence de lumière permet de synchroniser le cycle éveil/sommeil ;
- le colliculus supérieur ;
- le pulvinar.
Corps géniculé latéral
Le corps géniculé latéral (anciennement « genouillé externe » ou bien « corps genouillé latéral ») est un noyau relais, situé au niveau du thalamus, qui permet le transfert de l'information des yeux vers le cortex visuel. On a cru pendant un certain temps que c'était le simple et unique relais de la perception visuelle. En fait, ce n'est pas le seul noyau recevant de l'information visuelle (voir plus haut) et l'information qu'il reçoit ne vient pas que des yeux.
Chemins alternatifs de la vision
Le relais des fibres visuelles au niveau du corps géniculé latéral permet à environ 10 % de celles-ci de se projeter directement dans le colliculus supérieur. Des expériences (Weiskrantz, 1973) menées sur des sujets ayant subi une ablation chirurgicale du cortex visuel primaire ont montré que ceux-ci possèdent cependant une capacité visuelle sub-consciente, et qu'ils demeurent capables de localiser des objets, d'en déterminer l'orientation, éventuellement d'en percevoir la couleur, mais non de les comparer à des objets consciemment perçus (en cas d'ablation unilatérale). La sensation est donc inexploitable par le sujet (intuitive) et ne se manifeste pas, à l'exception d'objets en mouvement, de manière spontanée. Ce phénomène est nommé « blindsight » en anglais.
Des expériences complémentaires (Nakamura et Mishkin, 1980-1986) ont démontré qu'on pouvait provoquer la cécité consciente en isolant totalement le cortex visuel primaire du reste de l'encéphale, même si le tractus optique demeurait intact. La conscience visuelle résulterait donc d'un processus complexe de dialogue entre la perception corticale « classique » de la zone V1, les autres zones corticales V et les voies sous-corticales ou thalamiques « inconscientes » (Weiskrantz, 1999).
Deux zones du cortex préfrontal (Brodmann 9 et 46 droite) ont été identifiées dans le mécanisme de vision consciente. 9 pourrait correspondre à une zone de discrimination entre vision consciente et subconsciente, et 46 intégrerait et synthétiserait les perceptions visuelles corticales et thalamiques (Barbur, Sahraie, Weiskrantz, 1996).
Radiations optiques
Les fibres nerveuses qui relient le corps genouillé latéral au cortex visuel constituent la radiation optique d'un hémisphère. Une lésion d'une radiation optique conduit à la perte partielle ou totale de la vision dans l'hémisphère concerné (une anopsie). C'est donc par cette voie que se fait la perception visuelle consciente.
Cortex visuel
Le cortex visuel est le système le plus important en taille dans le cerveau humain et est responsable des traitements des images. Il effectue des traitements de bas niveau (orientation, contours) mais aussi de haut niveau (formes, mouvements).
Intégration
Une des interrogations les plus importantes autour du système visuel en 2019 chez l'homme, est le peu d'informations échangées entre l'œil et le système visuel du cerveau. Une grande partie de ce que nous voyons est « conjecturé » par le cerveau[5]. L'information provenant de l'œil passe par un goulot d'étranglement informationnel avant d'atteindre le cortex visuel du cerveau, qui traite fortement le signal épars[5].
Notes et références
- ↑ Jean-Michel Leuliot, « Les mécanismes de retransmission de la performance », dans Jean-Paul Callède, La recherche de performance, Maison des Sciences de l’Homme d’Aquitaine, , 33–41 p. (lire en ligne)
- ↑ Margaret Livingstone et David Hubel, « Segregation of Form, Color, Movement, and Depth: Anatomy, Physiology, and Perception », Science, vol. 240, no 4853, , p. 740–749 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.3283936, lire en ligne, consulté le )
- 1 2 3 Bruno Gepner, « "Malvoyance? du mouvement dans l'autisme infantile ?: Une nouvelle approche neuropsychopathologique développementale », La psychiatrie de l'enfant, vol. 44, no 1, , p. 77 (ISSN 0079-726X et 2102-5320, DOI 10.3917/psye.441.0077, lire en ligne, consulté le )
- ↑ Y. De Prost, Étude clinique et physiopathologique des érythèmes fessiers du nouveau-né et du nourrisson, vol. 13, (ISSN 0929-693X, DOI 10.1016/s0929-693x(06)80001-1, lire en ligne), p. 1
- 1 2 Kevin Hartnett, « A Mathematical Model Unlocks the Secrets of Vision », sur Quanta Magazine (consulté le )
Liens externes
- Ressources relatives à la santé :
- FMA
- Uberon
- (no + nn + nb) Store medisinske leksikon
- (cs + sk) WikiSkripta