الذاكرة هي العملية المعرفية حيث يتم التذكير بالخبرات والتعلم والاعتراف. تشكيل الذاكرة هو نتاج المرونة المخية، والتغيرات الهيكلية داخل المشابك التي تخلق ارتباطات بين المحفزات. يتم ترميز المحفزات ضمن أجزاء من الثانية. ومع ذلك، فإن الحفاظ على الذكريات على المدى الطويل يمكن أن يستغرق دقائق إضافية، أيام، أو حتى سنوات وتصبح ذاكرة مستقرة (أكثر مقاومة للتغيير أو التدخل). ولذلك، فإن تشكيل ذاكرة معينة يحدث بسرعة، ولكن تطور الذاكرة غالبا ما تكون عملية مستمرة.
وقد تبين أن عمليات الذاكرة تكون مستقرة عن طريق النوم الليلي وحتى القيلولة في النهار. ويلاحظ بعض مراحل النوم لتحسين ذاكرة الفرد، على الرغم من أن هذه المهمة محددة. عموما، يتم تعزيز الذكريات الإعلانية عن طريق النوم الموجة البطيئ، في حين يتم تعزيز الذكريات غير التصريح عن طريق نوم حركة العين السريعة (ريم) ، على الرغم من أن هناك بعض التناقضات بين النتائج التجريبية.

التاريخ
في عام 1801، افترض ديفيد هارتلي لأول مرة أن الحلم غير الروابط الكواكبية الارتباطية داخل الدماغ خلال تحلم فترات الأحلام. وكان الفكرة القائلة بأن النوم له تأثير تصالحي عقلي، وترسخ ودمج الذكريات والأفكار، كان مقبولا من الناحية الفكرية بحلول نهاية القرن التاسع عشر. في "بيتر وويندي"،[1] يا باري كتب "انها العادة الليلية لكل أم جيدة بعد نوم أطفالها الوقوع في عقولهم ووضع الأمور مباشرة صباح اليوم التالي، وإعادة حزم في أماكنها الصحيحة العديد من المقالات التي تجولت خلال النهار. ... عندما تستيقظ في الصباح، تم طي الشذوذ والمشاعر الشريرة التي ذهبت إلى السرير على صغيرها قد وضعت في الجزء السفلي من عقلك. وعلى القمة، بثت بشكل جميل، تنتشر أفكارك أجمل، وعلى استعداد لتضع على. "قصص بيتر بان تجري في عالم عقلي وتحتوي على العديد من التلميحات إلى جوانب علم النفس المعرفي، ووبعضها يسبق التحقيق العلمي الرسمي. .[2]
أجريت أول دراسة شبه متعددة المنهجية للنوم والذاكرة في عام 1924 من قبل جينكينز ودالبنباخ، لغرض اختبار نظرية اضمحلال الذاكرة هيرمان إيبينهاوس..وأظهرت نتائجها أن الاحتفاظ الذاكرة كان أفضل بكثير بعد فترة من النوم تعارض لنفس الفاصل الزمني مستيقظا. لم يكن حتى عام 1953 عندما تم تحديد النوم في النوم السريع حركة العين والنوم حركة العين السريعة غير أن الدراسات التي أجريت تركز على تأثير مراحل النوم محددة على الذاكرة.[3] كما السلوكية الخصائص من آثار النوم والذاكرة أصبحت على نحو متزايد لمزيد من التفهم ودعم الباحثين يتجهون إلى فهم بسيط لاسس عصبية النوم والذاكرة.
دورات النوم

النوم يتقدم في الدوري الأزياء من خلال خمس مراحل. أربعة من هذه المراحل مجتمعة حركة العين غير السريعة (NREM) النوم في حين أن آخر دورة حركة العين السريعة الفترة. دورة تستغرق حوالي 90-110 دقائق لإكمال. اليقظة هي وجدت من خلال EEG تدابير تتسم موجات بيتا التي هي أعلى في التردد وأقل في السعة وتميل إلى التحرك بشكل غير متسق بسبب الكمية الهائلة من المحفزات شخص لقاءات بينما مستيقظا.
- قبل النوم هو فترة من انخفاض الوعي الإدراكي حيث نشاط الدماغ تتميز موجات ألفا التي هي أكثر الإيقاعي، أعلى في السعة وانخفاض في وتيرة مقارنة مع موجات بيتا.
- المرحلة الأولى تتميز النوم الخفيف ويستمر حوالي 10 دقائق. موجات الدماغ تدريجيا الانتقال إلى موجات ثيتا.
- المرحلة الثانية أيضا يحتوي على موجات ثيتا; ومع ذلك، عشوائية رشقات نارية قصيرة من زيادة التردد يسمى النوم مغزل هي السمة المميزة لهذه المرحلة.
- المرحلة ثلاثة أربعة هي مشابهة جدا معا تعتبر "النوم العميق". في هذه المراحل نشاط الدماغ الانتقال إلى موجات دلتا التي هي أدنى في تواتر أعلى في السعة. اثنين من هذه المراحل مجتمعة تسمى أيضا نوم الموجة البطيئة (SWS).
- المرحلة الخامسة, REM, هي واحدة من الأكثر إثارة للاهتمام مراحل أنماط موجات المخ مماثلة لتلك التي شوهدت في استرخاء اليقظة. وهذا هو يشار إلى "النوم" و هي الفترة الأكثر الحلم يحدث. النوم REM هو يعتقد أيضا أن تلعب دورا في التنمية المعرفية من الرضع والأطفال كما أنها تنفق أكثر من ذلك بكثير من النوم في عيني فترات بدلا من البالغين.
مصطلحات الذاكرة
الاستقرار مقابل تعزيز
استقرار الذاكرة هو ترسيخ الذاكرة في المكان، وضعف يتم تأسيس اتصال. استقرار الإجرائية الذكريات حتى يمكن أن تحدث أثناء ساعات الاستيقاظ، مما يوحي بأن محددة غير معلن المهام المحسنة في غياب من النوم.[4] عندما الذكريات قال إلى تعزيز، ومع ذلك، فإن الاتصال هو تعزيز التدريب وكذلك توصيله إلى الأخرى ذات الصلة الذكريات مما يجعل استرجاع أكثر كفاءة. في حين أن استقرار غير معلن الذكريات يمكن أن تحدث أثناء حالة أرق, تعزيز هذه الحسية والحركية الذكريات وقد تبين أن تحدث أثناء النوم الليلي.
العمليات المعتمدة على الاستخدام مقابل العمليات المعتمدة على التجربة
نشاط الدماغ الذي يحدث أثناء النوم هو المقررة بطريقتين: استخدام-التبعية تجربة-التبعية.[5] استخدام تعتمد على نشاط الدماغ نتيجة العصبية الاستخدام التي وقعت خلال ساعات الاستيقاظ. أساسا هو العصبية تجديد النشاط الذي يحدث إذا كنت قد تعلمت شيئا جديدا أم لا.
الخبرة تعتمد على نشاط الدماغ نتيجة الوضع الجديد، البيئة، أو علم المهمة أو الوقائع التي حدثت في فترة ما قبل النوم بفترة. هذا هو نوع من نشاط الدماغ التي تدل على توطيد الذاكرة/تعزيز.
غالبا ما يكون من الصعب التمييز بين البلدين في المرحلة التجريبية لأن الإعداد وحده..... هذه البيئة الجديدة سيكون رأيت في النوم نشاط الدماغ جنبا إلى جنب مع المكتسبة حديثا المهمة. لتجنب هذا، معظم المجربون نصر المشاركين قضاء يوم واحد في التجريبية الشرط قبل أن يبدأ الاختبار لذا لا رواية واحدة تبدأ التجربة. هذا يضمن أن البيانات التي تم جمعها عن تجربة تعتمد على نشاط الدماغ هو محض من رواية المهمة.
توطيد
توطيد الذاكرة هي العملية التي تأخذ في البداية غير المستقرة التمثيل بترميز في أكثر قوي وفعال وناجع. في هذه الدولة الجديدة، ذاكرة أقل عرضة التدخل. هناك أساسا ثلاث مراحل توطيد الذاكرة وكل يعتقد أن يسهل النوم أو عدم النوم:
- الاستقرار هو ترميز الذاكرة الذي يستغرق سوى 6 ميلي ثانية.
- التعزيز هو عملية مستمرة من توطيد التي يمكن أن تحدث خلال دقائق, 7 ساعات، أيام ولكن ليس لفترة أطول. بعد النوم السلوكية الأنشطة يمكن أن تظهر تحسينات كبيرة في غياب الممارسة.
- التكامل أيضا يمكن أن يستغرق ساعات أو سنوات وهي عملية ربط مؤخرا ترميز الذكريات في الذاكرة القائمة الشبكات.
دعائم الاستقرار
للوحدة الذاكرة ينطوي على استرجاع بالفعل الموحدة الذاكرة (صريحة أو ضمنية) ، إلى قصيرة المدى أو الذاكرة العاملة. هنا يتم إحضارها إلى عطوب الدولة حيث المعلومات اللاحقة يمكن أن تتداخل مع ما هو حاليا في الذاكرة، وبالتالي تغيير الذاكرة. هذا هو المعروف باسم رجعي التدخل وهو هام للغاية القضية على المحكمة العين إفادات الشهود.[6]
التدريب البدني والحرمان من النوم بعد التدريب
الباحثين نهج دراسة النوم والذاكرة من زوايا مختلفة. بعض الدراسات لقياس تأثير الحرمان من النوم بعد رواية مهمة تدريس (موضوع يتعلم المهمة وهو محروم من النوم بعد ذلك). هذا ويشار إلى أنه بعد انتهاء التدريب الحرمان من النوم. على العكس من ذلك، تجارب أخرى أجريت لقياس آثار الحرمان من النوم قبل مهمة تم تدريسها (هذا الموضوع هو الحرمان من النوم ثم يتعلم مهمة). هذا ويشار إلى التدريب قبل النوم.
معالجة الذاكرة دون اتصال
هذا هو تجهيز ذكريات من الوعي واعية. بعد الانتهاء من قراءة شيء ما، على سبيل المثال هذه المقالة جدا عن النوم والذاكرة، الدماغ يستمر في معالجة المعلومات حتى يمكنك إنجاز المهام الأخرى مثل لعب لعبة كرة القدم. هذا "غير متصل" المعالجة كما يحدث في النوم الخاصة بك.
طرق قياس الذاكرة
التدابير السلوكية
- وهي النفس التي تأمر لافتا المهمة هي مهمة من الذاكرة حيث مشارك قدم مع عدد من الصور (أو الكلمات) التي رتبت على الشاشة. عدة محاكمات يتم عرض كل ترتيبات مختلفة والتي تحتوي على بعض الكلمات أو الصور. المهمة المشارك أن نشير إلى كلمة أو صورة لم تكن قد سبق وأشار إلى ذلك في محاكمات أخرى.[7]
- في حداثة التمييز مهمة المشاركين يتم عرض تجربتين من صورة العرض ثم محاكمة ثالثة تحتوي على خليط من الصور من المحاكمة. مهمتهم هي تحديد ما إذا كانت الصورة من أحدث عرض أو السابقة.
- في الطريق استرجاع مهمة التعلم المكاني يحدث فيها مشارك الجولات الافتراضية مكان معين (مثل المدينة أو المتاهة). يطلب من المشاركين بجولة تقريبا نفس الشيء في وقت لاحق أثناء تصوير الدماغ يستخدم لقياس النشاط.
- وهو تقرن كلمة النقابي المهمة يتكون من مرحلتين. خلال المرحلة الأولى (الاستحواذ) ، ردود إقران المنتسبين المهمة المستفادة تصبح الاستدعاء. في المرحلة الثانية (النقابي المرحلة) ، هذا الموضوع يتعلم الزوج كل رد منفصل التحفيز. على سبيل المثال، جديلة البصرية توفر معلومات عن ما هي الكلمات التي يجب أن يذكر بعد التحفيز والكلمات إزالتها.[8]
- في مرآة تتبع المهمة يطلب من المشاركين أن تتبع العديد من الأرقام بسرعة وبدقة قدر الإمكان مما إلا أنها يمكن أن نرى في المرآة. سجلت سرعة وكذلك مقدار ما تحيد عن الصورة الأصلية (دقة).
- في موريس المياه المتاهة المهمة الفئران تستخدم لاختبار التعلم المكاني في نوعين من الشروط: المكانية فراغي. في المكانية حالة منصة المخفية باستخدام الماء العكر وفي فراغي حالة منصة مرئية. المكاني حالة الفئران يجب أن تعتمد على الذاكرة المكانية للعثور على منصة في حين فراغي شرط يستخدم لأغراض المقارنة.

- في المسلسل وقت رد الفعل المهمة (SRT المهمة) مهمة حيث المواضيع الوجه شاشة الكمبيوتر حيث العديد من علامات يتم عرض هذا مكانيا ذات الصلة علامات على لوحة المفاتيح الخاصة بهم. المواضيع ويطلب الرد بسرعة وبدقة قدر الإمكان إلى ظهور التحفيز أدناه واحدة من علامات. المواضيع يمكن تدريبهم على المهمة سواء مع تعليمات واضحة (على سبيل المثال هناك تسلسل اللون المقدمة التي يجب أن يتعلمها) أو ضمنية منها (على سبيل المثال experimentor لا تذكر تسلسل اللون، ومن ثم ترك المواضيع التي نعتقد أنها تجري في اختبار السرعة). عند هذه المهمة المستخدمة في الدراسات النوم بعد وقت التأخير، مواضيع اختبار استبقاء.[9]
- في الوصول إلى فهم المهمة القوارض تعلمت المهرة forelimb المهمة. النوم تحسين سرعة الحركة مع المحافظة على الدقة. دون اتصال هذه التحسينات تم ربط كل من اعادتها من المهام ذات الصلة الفرق خلال حركة العين غير السريعة (NREM) النوم والزمانية التحولات التي أكثر بإحكام موتور القشرية الفرق إلى الحركات.[10]
- في neuroprosthetic المهمة القوارض المدربين لأداء بسيط الدماغ والآلة واجهة مهمة في نشاط مجموعة من السيارات القشرية الوحدات المستخدمة للسيطرة على ذراع ميكانيكية تعلق على تغذية صنبور. بعد نجاح التعلم المهمة المتعلقة الوحدات على وجه التحديد شهدت زيادة تأمين الاتساق إلى الموجة البطيئة النشاط (SWA) أثناء النوم. الوقت الذي يقضيه في سوا وتوقع المكاسب الأداء عند الاستيقاظ.[11]
- في منع التنصت المهمة يطلب من المشاركين كتابة سلسلة من خمسة أرقام مع المهيمنة أو اليد غير المسيطرة (المحددة في التجربة) ، مدة محددة من الوقت، تليها فترة راحة. عدد من هذه التجارب تحدث والكمبيوتر يسجل عدد متواليات الانتهاء من تقييم سرعة ومعدل الخطأ لتقييم دقة.
- وهو الاصبع التنصت اختبار يستخدم عادة عندما نقية الحركية المهمة المطلوبة. الاصبع التنصت يتطلب اختبار المواضيع باستمرار اضغط أربعة مفاتيح (عادة عددية مفاتيح) على لوحة المفاتيح مع nondominant اليد في تسلسل مثل 4-3-1-2-4 ، كمية معينة من الوقت. اختبار يتم عن طريق تحديد عدد من الأخطاء.[12]
تدابير التصوير العصبي
الأعصاب يمكن تصنيفها إلى فئتين، سواء المستخدمة في حالات متفاوتة اعتمادا على ما هو نوع المعلومات المطلوبة. الهيكلية التصوير تتعامل في الغالب مع بنية الدماغ (التصوير المقطعي) في حين وظيفية التصوير تتعامل بشكل كبير مع عمليات التمثيل الغذائي في ما يتعلق التشريحية عمل (التصوير المقطعي بالإصدار البوزيتروني, التصوير بالرنين المغناطيسي الوظيفي). في السنوات الأخيرة، العلاقة بين النوم والذاكرة العمليات قد ساعد على تطوير هذه تقنيات التصوير العصبي.[13]
التصوير المقطعي بالإصدار البوزيتروني (PET) في عرض وظيفي عمليات الدماغ (أو أجزاء أخرى من الجسم). أ البوزيترونالتي ينبعث منها النويدات المشعة يتم حقنها في مجرى الدم تنبعث أشعة جاما التي يتم الكشف عنها بواسطة التصوير الماسح الضوئي. تحليل الكمبيوتر ثم يسمح 3-الأبعاد إعادة بناء المنطقة في الدماغ أو الجسم جزء من الفائدة.
التصوير بالرنين المغناطيسي الوظيفي (الرنين المغناطيسي الوظيفي) هو نوع من تصوير الدماغ أن يقيس التغير الأكسجين في الدم بسبب نشاط الخلايا العصبية. البيانات الناتجة يمكن تصور كصورة من الدماغ مع الملونة تمثيل التنشيط.
التدابيرالجزيئية
على الرغم من أن هذا يمكن أن ينظر إليها على غرار تقنيات التصوير العصبي، الجزيئية تدابير تساعد على تعزيز مجالات التنشيط التي لولاها أن عويص إلى الأعصاب. واحدة من هذه التقنية التي الإيدز في كل الزمنية والبصرية القرار من الرنين المغناطيسي الوظيفي هو الدم الأكسجين على المستوى تعتمد (جريئة) استجابة. تغييرات جريئة استجابة يمكن أن ينظر إليه عندما يكون هناك مستويات مختلفة من التنشيط في المناطق المشتبه فيها من عمل. تزويد الطاقة إلى الدماغ في شكل الجلوكوز و الاكسجين (والتي يتم نقلها عن طريق الهيموغلوبين). إمدادات الدم باستمرار تنظيمها بحيث مجالات التنشيط تلقي كميات أكبر من الطاقة مقارنة مع المناطق التي هي أقل تفعيلها.[14] في التصوير المقطعي بالإصدار البوزيتروني ، استخدام النويدات المشعة (النظائر قصيرة مع نصف حياة) يسهل البصرية القرار. هذه النويدات المشعة تعلق السكر، الماء و النشادر بحيث سهلة الاستيعاب في تنشيط مناطق الدماغ هو إنجاز. مرة واحدة هذه المشعة استشفاف يتم حقنها في مجرى الدم ، والكفاءة موقع العمليات الكيميائية يمكن ملاحظتها باستخدام الحيوانات الأليفة.[15]
طرق قياس النوم
التدابير الكهربية
الأسلوب الرئيسي من قياس النوم في البشر هو دراسة النوم (باريس سان جيرمان). لهذا الأسلوب المشاركون في كثير من الأحيان يجب أن تأتي إلى المختبر حيث يمكن للباحثين استخدام باريس سان جيرمان قياس أشياء مثل إجمالي وقت النوم، النوم الكفاءة، خدمة بعد بداية النوم، والنوم تجزئة. باريس سان جيرمان يمكن رصد مختلف وظائف الجسم بما في ذلك نشاط الدماغ (المخ), حركة العين (electrooculography), حركة العضلات (الكهربائي) ، و إيقاع القلب (تخطيط القلب الكهربائي).
تخطيط كهربية الدماغ (EEG) هو الإجراء الذي يسجل النشاط الكهربائي على طول فروة الرأس. هذا الإجراء لا يمكن تسجيل نشاط الخلايا العصبية الفردية، ولكن بدلا من ذلك التدابير الشاملة متوسط النشاط الكهربائي في الدماغ.
Electrooculography (EOG) يقيس الفرق في الجهد الكهربائي بين الجبهة والجزء الخلفي من العين. هذا لا قياس استجابة الفرد للمؤثرات البصرية، ولكن بدلا من ذلك التدابير العامة حركة العين.
الكتروميوجرافي (فريق الإدارة البيئية) يسجل النشاط الكهربائي في العضلات والهيكل العظمي. جهاز يسمى electromyograph يقيس القدرة الكهربائية من خلايا العضلات لرصد حركة العضلات.
تخطيط القلب الكهربائي (ECG أو رسم القلب) التدابير الكهربائية الاستقطاب عضلات القلب باستخدام مختلف أقطاب كهربية وضعت بالقرب من الصدر والأطراف. هذا مقياس الاستقطاب يمكن استخدامها لمراقبة ضربات القلب.
التدابير السلوكية
Actigraphy هو مشترك مينيملي طريقة لقياس النوم. Actigraphy واحد فقط طريقة تسجيل الحركة. هذه الحركة يمكن تحليلها باستخدام مختلف actigraphic البرامج. على هذا النحو، actigraph غالبا ما يمكن ارتداؤها على غرار ساعة أو حول الخصر حزام. لأنه هو الحد الأدنى invase وغير مكلفة نسبيا، هذا الأسلوب يسمح التسجيلات خارج المختبر الإعداد لعدة أيام في وقت واحد. ولكن actigraphy في كثير من الأحيان أكثر التقديرات وقت النوم (دي سوزا 2003 Kanady 2011).
تنافس النظريات
معظم الدراسات تشير إلى العجز المحددة في التقريرية الذكريات التي تشكل ما قبل أو ما بعد الحرمان من النوم REM. على العكس من ذلك، العجز في غير معلن الذاكرة تحدث قبل أو بعد NREM الحرمان من النوم. هذه هي المرحلة المحددة تعزيز نظرية. هناك أيضا المقترحة المزدوج خطوة الذاكرة الفرضية مما يشير إلى أن الأمثل التعلم يحدث عندما تتبع الذاكرة في البداية معالجتها في SWS ثم النوم REM. دعم هذا هو مبين في العديد من التجارب حيث تحسين الذاكرة أكبر مع إما SWS أو النوم REM مقارنة مع الحرمان من النوم، ولكن الذاكرة هو أكثر دقة عند فترة النوم يحتوي على كل SWS و النوم REM.
الذاكرة التقريرية
التعريفي الذاكرة هي ذاكرة واعية الأحداث. هناك نوعان من الذاكرة التقريرية: العرضية والدلالي. العرضية الذاكرة على تذكر الخبرات في حين الدلالي الذاكرة في تذكر حقائق محددة.
الذاكرة الزمنية
الزمنية الذاكرة يتكون من تذكر عندما محدد الذاكرة قد حدث. في دراسة تم وضع المشاركين في 4 مجموعات ؛ اثنين من مجموعات التحكم إما بالنظر إلى الكافيين أو وهمي و اثنين من المجموعات التي كانت حرموا من النوم لمدة 36 ساعة إما بالنظر إلى الكافيين أو وهمي. المهمة المستخدمة لقياس الزمنية الذاكرة تتألف من التمييز بين الأخيرة وأقل الأخيرة الوجه العروض. مجموعة من اثني عشر وجوه غير مألوفة قدمت بالتتابع كل 10 ثوان. الذاتي أمر لافتا المهمة استخدامها بعد ذلك لمدة 5 دقائق لمنع بروفة للحفاظ على متعب المشاركين المحتلة. هذا المطلوب منهم وضع علامة على أي بنود جديدة ينظر (أما الأسماء أو الأشكال المجردة) التي قدمت على 12 ورقة. مجموعة ثانية قدم، تليها أخرى الذاتي أمر لافتا المهمة ثم تسلسل عشوائي من 48 وجوه إما تحتوي سابقا تقديم الوجوه الجديدة أو تلك التي تم عرضها على المشاركين. سئلوا إذا كانوا التعرف على الوجوه سواء كانوا من الأولى أو المجموعة الثانية. النتائج تشير إلى أن الحرمان من النوم لا تؤثر بشكل كبير على التعرف على الوجوه، ولكن لا تنتج ضعف كبير الزمنية الذاكرة (تمييز الوجه الذي ينتمي إلى أي مجموعة). الكافيين تم العثور على أن يكون لها تأثير أكبر على النوم محرومة مقارنة بمجموعة العلاج الوهمي والحرمان من النوم ولكن لا يزال أداؤها أسوأ من كل سيطرة الجماعات. الحرمان من النوم وجد أيضا إلى زيادة معتقدات صحيحة، خاصة إذا كانوا على خطأ. دراسات تصوير الدماغ من أولئك الذين حرموا من النوم وجدت أن أكبر انخفاض في معدل التمثيل الغذائي في قشرة الفص الجبهي.[16]
التعلم اللفظي
الدم الأكسجين على المستوى تعتمد (جريئة) الرنين المغناطيسي الوظيفي في دراسة دروموند et al. لقياس استجابة الدماغ على التعلم اللفظي بعد الحرمان من النوم. بالرنين المغناطيسي تسجيل نشاط الدماغ أثناء التعلم اللفظي مهمة من المشاركين إما العادي بعد ليلة من النوم أو أولئك الذين حرموا من 34.7 (± 1.2) ساعات من النوم. المهمة بالتناوب بين خط الأساس حالة من تحديد ما إذا كانت الأسماء العلوي أو السفلي القضية التجريبية حالة حفظ قائمة الأسماء. نتائج الدراسة تشير إلى أن الأداء هو أسوأ بكثير على حرية أذكر من قائمة الأسماء عندما حرموا من النوم (في المتوسط 2.8 ± 2 كلمات) مقارنة العادي بعد ليلة من النوم (4.7 ± 4 كلمات). حيث من مناطق الدماغ تنشيط اليسار الفص الجبهي من القشرة, القشرة أمام الحركيةو الفص الصدغي وجد أن يتم تفعيلها خلال مهمة في راحة دولة منفصلة في مناطق قشرة الفص الجبهي كانت أكثر تفعيلها خلال مهمة في النوم يحرم الدولة. وكذلك ثنائية الفص الجداري ، يسار الوسط التلفيف الجبهيو الداخلية التلفيف الجبهي تم العثور على تنشيط لأولئك الذين حرموا من النوم. الآثار المترتبة على هذه النتائج هي أن الدماغ يمكن في البداية تعويض عن آثار الحرمان من النوم مع الاحتفاظ جزئيا سليمة الأداء الذي ينخفض مع زيادة الوقت على المهمة. الأولي هذا التعويض قد تكون وجدت في الثنائية مناطق كل الجبهي والفص الجداري وتفعيل قشرة الفص الجبهي هو يرتبط بشكل كبير مع النعاس.
الأداء المعرفي
الدماغية التنشيط أثناء الأداء على ثلاثة المهام المعرفية (التعلم اللفظي، والحساب، وينقسم الانتباه) مقارنة بعد النوم الطبيعي و 35 ساعة من إجمالي الحرمان من النوم (TSD) في دراسة أجراها دروموند والبني. استخدام الرنين المغناطيسي الوظيفي قياس هذه الاختلافات في الدماغ. في التعلم اللفظي المهمة، بالرنين المغناطيسي إلى المناطق المعنية في كلا اللفظي التعلم والتلقين. وجدت النتائج أن كلا TSD العادي ليلة من النوم أظهرت استجابة هامة في قشرة الفص الجبهي وبعد TSD عرض ردا من مجالات إضافية والتي شملت أخرى الفص الجبهي المجالات الثنائية أدنى الفصيص الجداري ومتفوقة الفص الجداري. زيادة في النعاس أيضا يرتبط مع تفعيل اثنين بطني الفص الجبهي ومناطق ارتباط بين زيادة تفعيل ثنائية الفص الجداري (والتي تشمل مجالات اللغة) وانخفاض مستويات انخفاض القيمة على الحرة أذكر أيضا وجدت التالية TSD. في الحساب المهمة النوم الطبيعي أظهرت المتوقع تفعيل ثنائية الفص الجبهي والجداري الذاكرة العاملة المناطق ولكن بعد TSD فقط أظهرت التنشيط في اليسار متفوقة الجداري الفص الأيسر القشرة أمام الحركية في الاستجابة مع أي مناطق جديدة لتعويض (كما تم العثور في التعلم اللفظي). زيادة النعاس أيضا يرتبط مع التنشيط في بطني الفص الجبهي المنطقة، ولكن فقط في منطقة واحدة. مقسمة مهمة الاهتمام جنبا إلى جنب سواء اللفظي التعلم الحسابية المهمة. الرنين المغناطيسي الوظيفي إلى أن الدماغية رد بعد TSD هو مماثلة لتلك التي من التعلم اللفظي المهمة (على وجه التحديد قشرة الفص الجبهي الأيمن ثنائية الفص الجداري، و التلفيف الحزامي عرض أقوى رد). والمغزى من هذه النتيجة هو أن مناطق الدماغ إضافية تفعيلها بعد كل من التعلم اللفظي واهتمام تقسيم المهام التالية TSD تمثل الدماغية التعويضية استجابة تفتقر إلى النوم. على سبيل المثال، هناك انخفاض في رد الأيسر والفص الصدغي خلال كل المهام التي تشارك في مختلف مهام التعلم أثناء حالة راحة ولكن مشاركة السفلي الأيسر والفص الجداري في المدى القصير اللفظي ذاكرة التخزين التالية TSD يشير إلى أن هذه المنطقة قد تعوض. لا مجالات جديدة الحسابية المهمة قد تشير إلى أن ذلك يعتمد بشكل كبير على الذاكرة العاملة حتى التعويض غير ممكن، بالمقارنة مع المهام مثل التعلم اللفظي التي تعتمد أقل على الذاكرة العاملة.[17]
النوم البطيء (SWS)
النوم البطيء يحدث أثناء المراحل 3 و 4 من عملية النوم.[18] موجة بطيئة النشاط بنسبة تصل إلى 25% بعد التعلم الضمني والوقت الذي يقضيه في مرحلة النوم قد ثبت لتحسين أداء ضمنا المستفادة المهمة بعد النوم.
العيانية أنظمة الدماغ
مركز صدى الارتباط العصبي
استخدم الباحثون الفئران من أجل التحقيق في آثار من رواية اللمس الكائنات على التطور طويل الأجل الرئيسية القوارض الدماغ الأمامي الحلقات الأساسية في أنواع محددة من السلوك، بما في ذلك هياكل مثل الحصين, بوتامين, المخية الحديثة و المهاد.[19] الفئران تم رصد ولكن لا ازعجت عن 48-96 ساعات، مما يسمح طبيعية خدمة-دورات النوم تحدث. في بعض نقطة أربعة رواية اللمس وضعت الأشياء في أربع زوايا الفأر في قفص. كانوا جميعا مختلفة جدا عن بعضها البعض وكانت هناك مجموعه ساعة واحدة. نشاط الدماغ خلال هذه الساعة كان بمثابة الأساس أو قالب مقارنة. تحليل البيانات يعني أن العصبية الجمعيات خلال SWS ترتبط بشكل كبير أكثر مع قوالب من خلال ساعات الاستيقاظ أو النوم REM. وكذلك هذه بعد التعلم، بعد SWS أصداء استمرت 48 ساعة أطول بكثير من مدة رواية كائن التعلم (ساعة واحدة) ، مشيرا إلى التقوية طويلة الأجل. لمزيد من التحليل على الخلايا العصبية إلى الخلايا العصبية قاعدة تظهر أي مجموعة فرعية من الخلايا العصبية (الدماغ هيكل) أن تكون مسؤولة عن أصداء أو المضادة أصداء (أنماط من النشاط بشكل ملحوظ أكثر تختلف من رواية التحفيز قوالب). فرق آخر لاحظت أعلى علاقة القمم في SWS يتفق مع أدنى معدل إطلاق الخلايا العصبية في الدماغ الأمامي، مقابل أن REM من النوم والاستيقاظ حيث معدل اطلاق النار هو أعلى. فمن افترضنا أن هذا يرجع إلى تدخل من المحفزات واردة أثناء فترات الاستيقاظ. في SWS لا يوجد واردة المحفزات لذلك الرواية تجربة يمكن ردها دون انقطاع.
صدى الترابط الحصين العصبى
دراسة Peigneux et al., (2004) إلى أن إطلاق تسلسل في الحصين الفرق خلال التعلم المكاني هي أيضا نشطة أثناء النوم، مما يدل على أن ما بعد التدريب النوم لها دور في المعالجة المكانية الذكريات. وأجريت هذه الدراسة إلى إثبات أن نفس الحصين المناطق يتم تفعيلها في البشر خلال مسار التعلم في بلدة الظاهرية، يتم تنشيط أثناء اللاحقة نوم الموجة البطيئة (SWS).[20] من أجل رصد هذا التنشيط، المجربون تستخدم الحيوانات الأليفة بالأشعة و الرنين المغناطيسي الوظيفي استخدام تدفق الدم إلى المخ كعلامة من متشابك النشاط. إيجاد الإشارة إلى أن كمية الحصين التنشيط أثناء نوم الموجة البطيئة ارتباطا إيجابيا على تحسن على جولة افتراضية المهمة في اليوم التالي، مما يدل على أن الحصين النشاط أثناء النوم يرتبط مع تحسن في أداء الذاكرة. هذه النتائج تثبت أن التعلم تعتمد على التشكيل في الحصين النشاط أثناء النوم يدل على تجهيز تعلمت سابقا العرضية الذاكرة المكانية آثار. هذا التشكيل من الحصين يؤدي إلى البلاستيك تغييرات في الدماغ وبالتالي تحسن في الأداء. نتائج هذه الدراسة أظهرت أن الذاكرة المكانية آثار يتم معالجتها في البشر بينما هم في NREM sleep. أظهرت رد فعل من تشكيل الحصين خلال SWS بعد التعريفي الذاكرة المكانية المهمة. المجربون كما وجدت أن في البشر، هناك تجربة تعتمد التحويرات من النشاط خلال النوم NREM في المناطق الحصين، ولكن ليس أثناء النوم REM بعد التعلم. الأدلة من هذه الدراسة هو كبير إلى فرضية أن المعلومات المستفادة أثناء اليقظة، هو تغيير تعزيز بينما البشر في النوم.
انخفاض الأستيل كولين
في هذه الدراسة مجموعتين من المشاركين في الليل وازنت الدراسة. اثنين من المهام التي تم تعلمها من قبل جميع المشاركين بين 10:00-10:30 مساء. التعريفي المهمة كان إقران المنتسبين كلمة قائمة من 40 الألمانية لغويا كلمة ذات الصلة أزواج. غير مصرحة المهمة مرآة-تتبع المهمة. الساعة 11:00 مساء كل المشاركين في ساعتين ضخ إما فيزوستيغمين أو وهمي. فيزوستيغمين هو أستيل مثبط; هو الدواء الذي يحول دون انهيار المثبطة الناقل العصبي أستيل كولين ، مما يتيح لها أن تبقى نشطة لفترة أطول في نقاط الاشتباك العصبي. النوم المجموعة وضعت على السرير في حين أن مجموعة أخرى بقيت مستيقظا. اختبار كل المهام التي جرت في 2:45 صباحا، بعد 30 دقيقة من النوم الفريق قد استيقظ; النوم التي كانت غنية في نوم الموجة البطيئة (SWS). وأظهرت النتائج أن زيادة ACh سلبا أذكر الذاكرة (التعريفي المهمة) ، في حالة النوم بالمقارنة مع المشاركين إعطاء الدواء الوهمي. على وجه التحديد، أذكر بعد النوم المجموعة الثانية أظهرت زيادة قدرها 5.2 ± 0.8 الكلمات مقارنة مع زيادة قدرها 2.1 ± 0.6 الكلمات عندما تم منح المشاركين أستيل مثبط. على العكس من ذلك، ولا سرعة ولا دقة انخفضت في غير معلن مرآة مهمة عندما تم منح المشاركين فيزوستيغمين ولا أداء المهمة كان المتضررة في أعقاب المجموعات عندما physotigmine كانت تدار. هذا يشير إلى أن الغرض من منظمة العمل ضد الجوع قمع خلال SWS يسمح الحصين تعتمد على الذاكرة التقريرية التوحيد ؛ مستويات عالية من ACh خلال SWS كتل الذاكرة اعادتها على الحصين.
- ملاحظة: لم يكن هناك أي ارتباط بين مقدار SWS و مستوى يذكر. توطيد الذاكرة يمكن أن تتعطل، ومع ذلك، إذا كانت أجزاء كبيرة في SWS في عداد المفقودين.
الزيادات في محاور دوران النوم
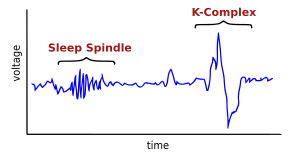
النوم مغزل قصيرة ومكثفة رشقات نارية من العصبية في المزامنة، تحدث في thalamo-القشرية الشبكات. هذه الذروة في وقت متأخر من الليل وهي تحديد خصائص المرحلة الثانية من النوم. النوم مغزل يعتقد أن تساعد في المعلومات توحيد أثناء النوم وقد ثبت أن زيادة بعد التدريب على السيارات المهمة.
دراسة باستخدام 49 الفئران إلى زيادة مغزل النوم أثناء نوم الموجة البطيئة التعليمية التالية. وقدم أدلة على أن زيادة التردد المغزل أثناء non-REM التالية يقترن المنتسبين السيارات-مهارة التعلم المهام. باستخدام EEG, النوم مغزل تم الكشف وأظهرت أن تكون موجودة فقط أثناء نوم الموجة البطيئة. بداية مع دراسة أولية الفئران خضعوا ست ساعات من رصد النوم بعد فترة من التعلم. وأظهرت النتائج أنه خلال الساعة الأولى بعد التعلم، كان هناك أكثر تأثير واضح على التعلم التضمين النوم المغزل الكثافة. إلا أن هذه الزيادة في المغزل الكثافة لا تعتمد على التدريب الشرط. وبعبارة أخرى، كانت هناك زيادة في مغزل بغض النظر عن كيف كانت الفئران المدربة. EEG أنماط أظهرت فرقا كبيرا في كثافة النوم مغزل مقارنة الكثافة السيطرة على مجموعة من الفئران، الذي لم يخضع أي تدريب قبل النوم مغزل تم قياس. هذا تأثير زيادة المغزل الكثافة استمر فقط لمدة ساعة الأولى في النوم التدريبية التالية، ثم اختفى في حدود الساعة الثانية إلى النوم.
مكافأة التعلم والذاكرة
في دراسة أجراها فيشر ولد عام 2009 ، [21] معرفة سابقة مكافأة مالية بعد انتهاء التدريب النوم ثبت أن تكون كبيرة تنبئ العام إصبع تسلسل التنصت الأداء. المواضيع قدمت مع إصبع تسلسل المهام التي يجب أن تتكرر في وقت لاحق. المواضيع قيل أنه سيكون هناك مكافأة عرضت للتحسين عند الاصبع التنصت تسلسل المهام. وهو مجموعة مراقبة لم يعط أي معرفة مكافأة. المواضيع فصلوا كذلك بالسماح النوم الفترة بين التدريب الأولي والاختبار النهائي مقابل مجموعة واحدة في حين أن مجموعة أخرى تواجه خدمة الاحتفاظ الفاصل. وخلص إلى أن المجموعة التي تلقت كل المعلومات عن مكافأة فضلا عن كونها قادرة على النوم عرض أعلى زيادة في الأداء على كل من الاصبع التنصت متواليات. المعرفة من المكافأة دون النوم والنوم دون معرفة الثواب سواء كانت تساهم بصورة كبيرة في تحسين الأداء. في جميع الحالات النوم تقرر أن يكون لها تأثير مفيد على الأداء العام بالمقارنة مع المجموعات التي خضعت اثني عشر ساعة في أعقاب فترة الاحتفاظ.
الذاكرة غير إعلانية
غير مصرحة الذاكرة هي الذاكرة المكتسبة من التجارب السابقة التي هي دون وعي تطبيق السيناريوهات اليومية. غير الذاكرة التقريرية ضروري لأداء تعلمت المهارات والعادات، على سبيل المثال الركض أو الطبخ المفضل وجبة. هناك ثلاثة أنواع من غير معلن الذكريات: الذاكرة الضمنية (اللاوعي والذاكرة، فتيلة) دور الذاكرة (الاشراط الكلاسيكي), و الذاكرة الإجرائية (تلقائي مهارة الذاكرة).
الحرمان من النوم
فسفرة ERK
خارج الخلية إشارة ذات الصلة تحركات، المعروف أيضا باسم الكلاسيكية خريطة كيناز، هي مجموعة من البروتين كيناز يقع في الخلايا العصبية. هذه البروتينات هي تفعيلها أو تعطيلها من خلال الفسفرة (إضافة مجموعة الفوسفات باستخدام ATP) ، ردا على الناقلات العصبية وعوامل النمو.[22] هذا يمكن أن يؤدي لاحقا البروتين البروتين التفاعلات إشارة transductions (الناقلات العصبية أو الهرمونات يحيل إلى الخلايا) ، والتي في نهاية المطاف يسيطر على جميع العمليات الخلوية بما في ذلك النسخ الجيني والخلايا دورات (مهم في التعلم والذاكرة). دراسة اختبار أربع مجموعات من الفئران في متاهة موريس المياه، مجموعتين في المكانية المهمة (منصة المخفية) مجموعتين في غير المكانية المهمة (مرئية منصة.) آثار ست ساعات من إجمالي الحرمان من النوم (TSD) تم تقييم المجموعة التجريبية (واحد المكانية مجموعة واحدة غير المكانية المجموعة) في كل المهام. بعد ست ساعات من TSD الفترة (أو النوم لفترة الضوابط) مجموعات من الفئران تم تدريبهم على أي مهمة ثم اختبار على مدار 24 ساعة في وقت لاحق. وبالإضافة إلى ذلك، فإن مستويات من إجمالي ERK الفسفرة (ERK 1 و ERK 2), البروتين الفوسفات 1 (الفقرة 1) ، MAPK الفوسفاتيز 2 (والأخيران كل من يشارك في dephosphorylation) تم تقييمها من قبل بقطع رؤوس أربعة مجموعات أخرى من الفئران (اثنين محروم من النوم واثنين من غير حرمان النوم), و إزالة hippocampuses بعد ست ساعات من TSD أو ساعتين بعد TSD (ثماني ساعات). وأظهرت النتائج أن TSD لم تنل من التعلم المكاني المهمة، ولكن ذلك لم يضعف الذاكرة. فيما يتعلق غير المكانية المهمة، التعلم مرة أخرى كان لا يختلف في TSD; ومع ذلك، الذاكرة في TSD الفريق كان في الواقع أفضل قليلا، وإن لم يكن بشكل كبير جدا. تحليل الحصين أظهرت أن TSD انخفاض كبير في مستويات إجمالي ERK الفسفرة بنحو 30%. TSD لم تؤثر على البروتينات في القشرة مما يدل على أن النقصان في ERK المستويات بسبب ضعف الإشارة في الحصين. وبالإضافة إلى ذلك، ولا الفقرة 1 أو MAPK الفوسفاتيز 2 مستويات متزايدة تشير إلى أن النقصان في ERK لم تكن بسبب dephosphorylation ولكن بدلا من ذلك نتيجة TSD. ولذلك يقترح أن TSD مكره له تأثير على العمليات الخلوية (ERK: النسخ الجيني الخ) ، الأساسية النوم التي تعتمد على الذاكرة اللدونة.
النوم REM
النوم REM تشتهر حية الإبداعات تشابه إلى الاحتمالات النواتج من الاستيقاظ شخص. هذه المرحلة من النوم تتميز العضلات ونى ونقص وسريعة ولكن منخفضة الجهد الكهربي EEG ، وكما يوحي اسمها، حركة العين السريعة. فمن الصعب أن تنسب الذاكرة المكاسب إلى مرحلة واحدة من النوم عندما قد يكون كامل دورة النوم هي المسؤولة عن تعزيز الذاكرة. الأبحاث الأخيرة التي أجرتها داتا et al.[23] استخدم تجنب المهمة تليها مرحلة ما بعد التدريب النوم REM الفترة لدراسة التغيرات في موجات P تؤثر على إعادة حصلت مؤخرا المحفزات. وقد تبين أن ليس فقط ف موجات متزايدة من خلال وظيفة التدريب النوم ولكن أيضا كثافة الأمواج. هذه النتائج قد يعني أن موجات P أثناء النوم REM قد تساعد على تنشيط مركز الحرجة والهياكل القشرية التعامل مع تعزيز الذاكرة. في Hennevin et al. الدراسة عام 1989 ، [24] في mesencephalic تشكيل شبكي (MRF) أعطيت الضوء التحفيز الكهربائيأثناء النوم REM ، والذي يعرف أن يكون لها تأثير مفيد على التعلم عند تطبيقها بعد التدريب. الفئران في تجربة تم تدريبهم على تشغيل في متاهة البحث عن المكافأة الغذائية. مجموعة واحدة من الفئران أعطيت غير الصحوة MRF التحفيز الكهربائية بعد كل من المتاهة التجارب مقارنة مع المجموعة الضابطة التي لم تتلق أي التحفيز الكهربائي. لوحظ أن حفز الفئران أداء أفضل بكثير فيما يتعلق خطأ الحد. هذه النتائج تعني أن الذاكرة الديناميكية العمليات التي تحدث أثناء التدريب وكذلك من خلال وظيفة التدريب النوم. دراسة أخرى من قبل Hennevin et al. (1998)[25] مكيفة الفئران الخوف من الضوضاء التي يرتبط اللاحقة القدم صدمة. الجزء مثيرة للاهتمام من التجربة هو أن الخوف الاستجابة إلى الضوضاء (تم قياسها في اللوزة) لوحظ عند الضوضاء قدمت أثناء النوم REM. هذا بالمقارنة مع مجموعة الزائفة مكيفة الفئران الذين لم يظهروا نفس amygdalar التنشيط من خلال وظيفة التدريب النوم. هذا يشير إلى أن العصبية تستجيب سابقا أبرز المحفزات يتم الاحتفاظ حتى أثناء النوم REM. ليس هناك نقص في الأبحاث التي أجريت على آثار النوم REM على عمل الدماغ ولكن الاتساق في النتائج هو ما اصاب البحوث التي أجريت مؤخرا. هناك أي ضمان بشأن ما وظائف REM النوم قد تؤدي أجسادنا وعقولنا ولكن البحوث الحديثة دائما التوسع واستيعاب الأفكار الجديدة إلى زيادة فهمنا من هذه العمليات.
موجات PGO
في الحيوانات، وظهور بونتو-geniculo-القذالي موجات (PGO موجات) هو ذات الصلة إلى أن من الاحتمالات النواتج حركات العين السريعة.[26] هذه الموجات هي الأكثر ملاحظته بوضوح خلال الانتقال من non-REM REM النوم. على الرغم من أن هذه phasic موجات لوحظ في كثير من أجزاء الحيوان الدماغ، هم الأكثر وضوحا في بونس، الركبي الوحشي والهيئات القذالي القشرة. Peigneux وآخرون، عام 2006 ، ذكرت أن النواة الركبية الجانبية و القذالي القشرة عرض مستويات أعلى من النشاط أثناء النوم REM من خلال اليقظة. هذا إضافة إلى نظرية التنشيط في هذه المجالات على غرار PGO موجة التنشيط في الحيوانات. جسري موجات شيوعا في الحيوانات كآلية تساعد على تيسير التعلم وتوطيد الذاكرة. تحسنا في أداء المهمة كان ينظر إلى نتيجة زيادة P موجات بين النوم REM الدورات.[27] في دراسة باستخدام وظيفة التعلم النوم REM الحرمان من آثار تحفيز موجة P مولد (الموجود في جسري السقيفة) من الفئران لوحظت. مجموعتين من الفئران خضع التعلم تجنب المهمة ثم سمح فترة النوم في حين أن مجموعة أخرى من الفئران التي حرمت من النوم. عند المقارنة بين مجموعتين النوم حرمان الفئران أظهرت عجز كبير في التعلم من بعد أن لم يطرأ النوم REM. في آخر الفئران المجموعة، ف مولد موجة وحفز باستخدام carbachol حقن الفئران ثم خضع الحرمان من النوم المرحلة. عند هذه الفئران مرة أخرى اختبار على التعلم فقد تبين أن تفعيل P مولد موجة أثناء النوم أدى إلى التعلم العادي يتحقق. وهذا من شأنه أن يشير إلى حقيقة أن تفعيل P موجات، حتى من دون النوم REM ، كان كافيا لتعزيز عمليات الذاكرة التي عادة لا يكون قد حدث.
ذاكرة الوجه الضمنية
الوجوه هي جزء مهم من الحياة الاجتماعية. أن تكون قادرة على التعرف والرد والعمل تجاه شخص يتطلب فاقد الوعي عمليات ترميز واسترجاع الذاكرة. تتم معالجة محفزات الوجه في التلفيف المغزلي (منطقة المخ القذالي الصدغي) وهذه المعالجة هي وظيفة ضمنية تمثل شكلا نموذجيا من الذاكرة الضمنية .[28] وقد رأى النوم REM لتكون أكثر فائدة لعمليات الذاكرة فيسوسباتيال الضمنية، بدلا من النوم الموجة البطيئة التي هي حاسمة لتوحيد الذاكرة صريح. ومن المعروف أن النوم REM لتجارب بصرية، والتي قد غالبا ما تشمل تصورات مفصلة من كونستانانس الإنسان. وقد استخدمت مهمة الاعتراف لقياس الألفة مع تسلسل يظهر سابقا من وجوه بعد فترة لاحقة من النوم REM. وقد شوهد أن التلفيف المغزلي كان نشطا أثناء التدريب، وفترة النوم REM، ومهمة الاعتراف أيضا. ومن المفترض أن آليات المخ خلال النوم REM، فضلا عن فتيلة التكرار النقي، يمكن أن تمثل الاعتراف الضمني من وجوه المعروضة سابقا.
وقد أظهرت الأبحاث السابقة النوم الريمي لإعادة تنشيط التجميعات العصبية القشرية بعد التدريب على مهمة وقت رد الفعل المتسلسل (SRT)، وبعبارة أخرى النوم ريم يعيد المعالجة التي حدثت في حين تعلم احد مهمة ضمنية في ساعات الاستيقاظ السابقة[29] ومع ذلك، فإن عناصر التحكم لم تكتمل مهمة سرت، وبالتالي لا يمكن للباحثين أن يفترضوا إعادة تنشيط بعض الشبكات لتكون نتيجة تسلسل / قواعد اللغة المستفادة ضمنا لأنها يمكن أن تكون ببساطة بسبب المعالجة فيسوموتور الابتدائية التي تم الحصول عليها في كلتا المجموعتين. للإجابة على هذا السؤال تم إعادة تشكيل التجربة وأضيفت مجموعة أخرى الذين شاركوا أيضا في مهمة سرت. لم يشهدوا أي تسلسل لمهمة سرت (مجموعة عشوائية)، في حين أن المجموعة التجريبية لم تجربة تسلسل (المجموعة الاحتمالية)، على الرغم من دون وعي واعية. نتائج المسح الضوئي بيت تشير إلى أن كونيوس الثنائية كانت أكثر نشاطا بشكل ملحوظ خلال ممارسة سرت فضلا عن النوم ريم بعد التدريب في المجموعة بروبابيليستيك من مجموعة عشوائية.. وبالإضافة إلى ذلك، تم زيادة هذا التنشيط بشكل ملحوظ خلال النوم ريم مقابل مهمة سرت. وهذا يشير إلى أن مناطق معينة من الدماغ تشارك على وجه التحديد في مرحلة ما بعد معالجة المعلومات المتسلسلة. ويدعم ذلك أيضا حقيقة أن مرفق التنوع البيولوجي الإقليمي (rCBF) خلال فترة ما بعد التدريب في ريم يتم تشكيله بمستوى تعليمي عالي المستوى، ولكن ليس منخفضا تم الحصول عليه قبل النوم. لذلك، يتم تضمين مناطق الدماغ التي تشارك في عملية التعلم من قبل كل من هيكل متتابعة من المواد المستفادة (زيادة تفعيل في كونيوس)، وكمية التعلم عالية الترتيب(rCBF).
REM الحرمان من النوم وعصبية العوامل
تم تقييم آثار الحرمان من النوم ريم (رسد) على العوامل العصبية، وتحديدا عامل نمو الأعصاب (نغف) والعامل العصبي المستمدة من الدماغ (بدف)، في عام 2000 من قبل سي إت إل. نيوروتروفينز هي البروتينات الموجودة في الدماغ والمحيطات التي تساعد في بقاء، وتعمل وتوليد الخلايا العصبية. وهذا هو عنصر مهم في عملية اللدونة متشابك، الأساس الأساسي الكيميائية في تشكيل الذكريات. .[30] ساي وآخرون، أقطاب إدراجها في جماجم سبعة أزواج من الفئران لقياس الكهربائي (EEG)، وإدخال السلك في عضلات الرقبة من الفئران لقياس كهربية (EMG)، وهي تقنية تستخدم لقياس كمية من نشاط العضلات. شهدت نصف الفئران وREM من النوم فترة الحرمان لمدة ست ساعات، في حين شهد النصف الآخر فترة النوم لمدة ست ساعات، تحتوي على جميع دورات النوم. وأظهرت النتائج أنه في الفئران في المجموعة الحرمان REM من النوم أظهرت انخفاض مستوى عامل التغذية العصبية المشتقة من الدماغ في المخيخ (التنسيق والتعلم الحركي) والدماغ (الحسي والصعود السيارات المسار)، على العكس الحصين (الذاكرة على المدى الطويل والمكانية الملاحة)، أظهرت انخفاض في مستويات عامل نمو الأعصاب. وقد ثبت أن البروتين بدف ضروري للتعلم الإجرائي (شكل من الذاكرة غير إعلانية). منذ التعلم الإجرائي وقد عرضت أيضا توطيد وتعزيز تحت النوم REM، يقترح أن انخفاض قيمة مهام التعلم الإجرائية ويرجع ذلك إلى نقص البروتينات BDNF في المخيخ وجذع الدماغ خلال RSD. .في ما يتعلق بغاز نغف، فإن الدماغ الأمامي القاعدي (إنتاج وتوزيع أش في الدماغ)، وبشكل أكثر تحديدا منطقة الحاجز الإنسي، يرسل كوليني (مثار في الحصين) و غابينرجيك (مثبط) الناقلات العصبية من خلال الألياف إلى الخلايا المستهدفة الحصين. هذه الخلايا المستهدفة ثم تفرز نغف الذي يلعب دورا رئيسيا في الحالة الفسيولوجية للحصين ووظائفها. وقد لوحظ أن النوم ريم يزيد من إفراز نغف، ولذلك فقد اقترح أن خلال النشاط رسد الكوليني يقلل مما أدى إلى انخفاض في نغف وانخفاض في التعلم الإجرائي..

إعادة تنظيم نظام المخ المجهري
ووكر وفحص ستيكغولد أنه بعد اكتساب الذاكرة الأولية، ينظم النوم تمثيل الذاكرة على مستوى النظم الكلية المخ. وكانت تجربتهم تتألف من مجموعتين؛ تم تدريس مجموعة النوم ليلا كتلة تسلسل الحركية مهمة التنصت في الليل، وضعت للنوم ثم إعادة اختبارها بعد 12 ساعة. وقد تم تدريس مجموعة اليوم بعد نفس المهمة في الصباح واختبارها بعد 12 ساعة مع عدم وجود النوم التدخل. تم استخدام فمري لقياس نشاط الدماغ أثناء إعادة الاختبار. وأظهرت النتائج بشكل ملحوظ أخطاء أقل / تسلسل في مجموعة النوم ليلا مقارنة مع مجموعة أعقاب اليوم. وأشارت النتائج فمري لمجموعة النوم ليلا زيادة التنشيط في القشرة الحركية الأولية الحق / M1 / الجيب الفص الجبهي (كونترا الجانبية إلى اليد كانت كتلة التنصت مع)، الحق الأمامي الفص الجبهي الإنسي، الحصين الأيمن (الذاكرة على المدى الطويل، المكانية الذاكرة)، المخطط البطني الأيمن (الحديبة الشمية، نواة أكومبنز)، وكذلك مناطق المخيخ (الفصيصات V1، V11). في مجموعة النهار، أظهرت الرنين المغناطيسي الوظيفي "انخفض" تنشيط إشارة ثنائيا في القشور الجداري (يدمج طرائق متعددة)، بالإضافة إلى القشرة القاعدية اليسرى (تنظيم التوازن)، الأيسر الزماني القطب (معظم الأمامي من القشرة الزمنية)، و اليسار، أدنى، فرونتو-بولار، كورتيكس. وقد أظهرت التحقيقات السابقة أن زيادات إشارة تشير إلى اللدونة الدماغ. زيادة النشاط إشارة ينظر في M1 بعد النوم يتوافق مع زيادة النشاط في هذا المجال ينظر خلال الممارسة؛ ومع ذلك، يجب على الفرد ممارسة لفترات أطول مما كان عليه أن ينام من أجل الحصول على نفس المستوى من زيادات إشارة M1. ولذلك، يقترح أن النوم يعزز تمثيل القشرية من المهام الحركية عن طريق توسيع نظام الدماغ، كما يتضح من زيادة النشاط إشارة.
الذاكرة العاملة
تعتبر مساحة العمل العقلية تمكين التخزين المؤقت واسترجاع المعلومات، الذاكرة العاملة أمر بالغ الأهمية لحل المشاكل وتحليل حالات مختلفة. سعة الذاكرة العاملة هي مقياس لعدد من وظائف المعالجة الذهنية واحد قادر على أداء متتالي. ويمكن تحقيق زيادة في سعة الذاكرة العاملة في واحدة مع استراتيجية تعرف باسم تشونكينغ. أريتك وآخرون. .[31] أجرى تسلسل إصبع التنصت التجربة التي تم عرض الموضوعات النقاط الملونة في تسلسل على شاشة المقابلة للأزرار على لوحة المفاتيح الخاصة بهم. عندما تم عرض لون كان الموضوع للرد عن طريق الضغط على اللون المناسب على لوحة المفاتيح. تم فصل المواضيع إلى ثلاث مجموعات. المجموعة الأولى تدرب باستمرار مع عدم وجود فترات من النوم. تم تدريب المجموعة الثانية وإعادة اختبارها على مدى عشر ساعات من اليقظة تليها ثماني ساعات من النوم والاختبار النهائي. تم تدريب المجموعة الثالثة في الساعة العاشرة مساء، يليها النوم لمدة ثماني ساعات. ثم تم اختبار هذه المجموعة في صباح اليوم التالي ومرة أخرى في وقت لاحق في نفس اليوم. أظهرت النتائج أن اليقظة كانت مؤشرا غير مهم لتحسين الأداء، ما لم تليها فترة من النوم. المجموعات التي سمح لها فترة ما بعد التدريب التدريب، بغض النظر عن وقته في إشارة إلى التدريب، كان مفيدا للتحسين على تسلسل التنصت الاصبع. وبلغ متوسط سعة الذاكرة العاملة الأولية للمجموعات ثلاث إلى أربع وحدات. في المجموعتين الثانية والثالثة تم زيادة سعة الذاكرة العاملة إلى متوسط 5-6 وحدات. واقترح أن التحسينات التي تعتمد على النوم قد تسهم في التحسين العام في قدرة الذاكرة العاملة مما يؤدي إلى تحسين الذكاء السوائل.
الحرمان من النوم
الحرمان من النوم، سواء كان الحرمان من النوم الكلي أو الحرمان من النوم الجزئي، يمكن أن يضعف الذاكرة العاملة في تدابير الذاكرة، وسرعة المعالجة المعرفية، والانتباه والتبديل المهمة. كاسيمنت إت آل. وجدت أنه عندما طلب من الموضوعات التعرف على الأرقام المعروضة على الشاشة عن طريق كتابتها على لوحة المفاتيح، وسرعة الذاكرة العاملة من الأشخاص الذين كان النوم يقتصر على أربع ساعات في الليل (حوالي 50٪ من كمية النوم العادية) كانت أبطأ 58٪ من مجموعات السيطرة التي سمح لهم ثماني ساعات كاملة من النوم. .[32]
المرونة
ان نموذج تبادل المعلومات ومعالجتها بالمخ متغير باستمرار، ومرونة. ولكي يتمكن الدماغ من دمج تجارب جديدة في مخطط مكرر، يجب أن يخضع لتعديلات محددة لتوحيد واستيعاب جميع المعلومات الجديدة.. ويمكن وصف المرونة كما تغير في القوة بين اثنين من الخلايا العصبية ذات الصلة. ويرى أكثر وضوحا العصبية في حالات الحرمان من النوم ريم أثناء نضج المخ. أظهرت قياسات الدماغ الإقليمية في نوم الريم حديثي الولادة المحرومين الفئران انخفاض حجم كبير في مجالات مثل القشرة المخية وجذع الدماغ. وقد حرمت الفئران خلال فترات حرجة بعد الولادة، وبالتالي لوحظ انخفاض حجم التشريحية. [33] باستخدام مهمة السعي (تستخدم لاختبار قدرات فيسوموتور) في تركيبة مع الرنين المغناطيسي الوظيفي، ماكيت وآخرون، 2003، [34] أن الزيادات في التنشيط شوهدت في حقل العين التكميلي ونواة المسنن الأيمن من الأشخاص الذين سمح لهم بالنوم بالمقارنة مع الأشخاص المحرومين من النوم. كما لوحظ أن التلم الصدغي العلوي الأيمن له مستويات تنشيط أعلى. عندما تم تحليل التوصيلية الوظيفية وجد أن النواة المسننة كانت أكثر ارتباطا بوظائف التلم الصدغي العلوي. وتشير النتائج إلى أن الأداء في مهمة البحث يعتمد على قدرة الشخص على فهم أنماط الحركة المناسبة من أجل استجمام الحركات المثلى. تم العثور على الحرمان من النوم لقطع العمليات البطيئة التي تؤدي إلى تعلم هذه المهارة الإجرائية وتغيير تغييرات الاتصال التي كان من الطبيعي أن ينظر إليها بعد ليلة من الراحة. وقد تم دراسة المرونة العصبية بدقة على مدى العقود القليلة الماضية، وأظهرت النتائج أن التغيرات الهامة التي تحدث في المناطق المعالجة القشرية لدينا القدرة على تعديل اطلاق الخلايا العصبية إلى كل من المحفزات الجديدة والخبرة السابقة.
تنظيم الناقلات العصبية
التغييرات في كمية من الناقل العصبي معينة وكذلك كيف تستجيب محطة ما بعد التشابك لهذا التغيير هي الآليات الكامنة من المرونة المخية. خلال النوم هناك تغييرات ملحوظة في الناقلات العصبية في جميع أنحاء المخ. أسيتيلكولين هو ناقل عصبي مثير أن ينظر إلى زيادة إلى مستويات الاستيقاظ القريب خلال النوم REM في حين بالمقارنة مع مستويات أقل خلال النوم موجة بطيئة.[35] وقد أظهرت الأدلة أن أداء نظام الذاكرة تعتمد الحصين (الذاكرة العرضية وذاكرة السيرة الذاتية) يتأثر مباشرة من التغييرات كوليني في جميع أنحاء دورة النوم بعد النوم. سوف مستويات عالية من أش تعزيز المعلومات التي تحققت أثناء اليقظة ليتم تخزينها في الحصين. ويتم ذلك عن طريق قمع الاتصالات الإثارة السابقة في حين تسهيل الترميز دون تدخل من المعلومات المخزنة سابقا. خلال النوم NREM، وخاصة النوم الموجة البطيئ، وانخفاض مستويات أش يسبب الإفراج عن هذا القمع والسماح للانتعاش التلقائي من الخلايا العصبية قرن آمون مما أدى إلى تسهيل توحيد الذاكرة..[36]
التعبير الجيني
في الآونة الأخيرة، تم العثور على ما يقرب من مائة الجينات التي التعبير عن الدماغ خلال فترات النوم.[37] م العثور على عدد مماثل من الجينات لتعزيز التعبير الجيني خلال اليقظة. هذه المجموعات من الجينات ترتبط بمجموعات وظيفية مختلفة والتي قد تعزز العمليات الخلوية المختلفة. الجينات التي أعرب عنها أثناء اليقظة قد تؤدي واجبات عديدة بما في ذلك تخصيص الطاقة، العصبي متشابك العصبي، والنشاط النسخي عالية وتثبيط متشابك في تعلم معلومات جديدة. كان هناك زيادة في النوم ذات الصلة في العمليات التي تنطوي على تركيب وصيانة المشبك. وتشمل هذه العمليات الاتجار الغشائي، إعادة تدوير الحويصلة متشابك، تشكيل البروتين الميالين الهيكلي، والكولسترول وتوليف البروتين. في دراسة مختلفة وجد أن هناك زيادة في النوم ذات الصلة في كالمودولين تعتمد على كيناز البروتين الرابع الذي شارك على وجه التحديد في الاكتئاب متشابك وفي توطيد الذاكرة على المدى الطويل.[38] هذه النتائج تشجع على العلاقة بين النوم وجوانب مختلفة من اللدونة العصبية.
جداول النوم البديلة
تعلم المهارات الحركية
تم النظر في تأثير القيلولة خلال النهار من قبل ووكر وستيكغولد (2005). أعطيت المجموعة التجريبية قيلولة بعد ظهر اليوم 60-90 دقيقة (دورة كاملة واحدة)، بعد أن تعلمت مهمة المهارات الحركية ذلك الصباح، في حين لم تحصل مجموعة السيطرة على أي قيلولة. تحسنت مجموعة قيلولة 16٪ عند اختبارها بعد قيلولة، في حين أن مجموعة عدم قيلولة لم تحدث أي تحسينات كبيرة. ومع ذلك، يبدو أن كل حتى خارج بعد أن نفس الليالي النوم؛ تحسنت مجموعة عدم قيلولة 24٪ وحسن مجموعة قيلولة فقط 7٪ أكثر لما مجموعه 23٪، متطابقة تقريبا. وفيما يتعلق التعلم المهارات الحركية، ويبدو أن القيلولة تسريع فقط تعزيز المهارات، وليس زيادة كمية التحسين..[39]
تعلم مهارات المرئيات
مثل الكثير من المهارات الحركية التعلم، وزيادة المهارات اللفظية التعلم بعد فترة قيلولة النهار. وقد أظهر الباحثون مدنيك وزملاؤه أنه إذا تم تدريس مهمة المهارات البصرية [العثور على مهمة] في الصباح واختبارها بشكل متكرر على مدار اليوم، فإن الأفراد سيصبحون أسوأ في هذه المهمة. الأفراد الذين سمح لهم قيلولة 30-60 دقيقة يبدو لكسب الاستقرار من المهارة كما لم يحدث أي تدهور. إذا سمح قيلولة 60-90 دقيقة (النوم REM ونوم موجة بطيئ)، وعرض الأفراد تعزيز. على عكس المهمة الحركية، لم يتم قمع تعزيز خلال النوم الليلي إذا كان الفرد قد نابد في وقت سابق. في حالة التعلم عن المهارات البصرية، وقد تبين أن القيلولة لمنع تدهور اليقظة وحتى تعزيز التعلم فوق وما بعد تعزيز يحدث في النوم الليلي..[40]
عمال المناوبة
وقد عرف العمال الذين يعملون طوال الليل بأنهم لديهم حوادث أكثر بكثير بالمقارنة مع العمال في النهار..[41] ويمكن أن يعزى ذلك إلى عدة عوامل، منها عدد أقل من الموظفين والإرهاق؛ ومع ذلك، فإن جزءا من المشكلة قد يكون العمال الفقراء الذاكرة العاملة وضعف مهارات الأداء بسبب ضعف توطيد الذاكرة. كل من المهام المستفادة ضمنا والمهام المستفادة صراحة تحسن بنسبة 20٪ تقريبا بعد ليلة كاملة من النوم..دون ليلة كافية النوم بين تعلم مهمة جديدة وأداء تلك المهمة، فشل الأداء في التحسن. لن يؤدي العمال الذين لم يعطوا كمية كافية من النوم، وخاصة في مرحلة NREM، بين التعلم وأداء المهمة، فضلا عن العمال الذين يحافظون على روتين النوم العادي..
النوم والشيخوخة
وغالبا ما يصبح النوم غير منظم في كبار السن ويمكن أن يؤدي إلى تفاقم انخفاض الذاكرة الموجودة مسبقا.
كبار السن الأصحاء
العلاقة الإيجابية بين النوم والذاكرة تنهار مع الشيخوخة. بشكل عام، كبار السن يعانون من انخفاض كفاءة النوم..[42] مقدار الوقت وكثافة النوم REM و SWS تنخفض مع التقدم في السن.[43][44][45] وبالتالي، فمن الشائع أن كبار السن لا يحصلون على زيادة في الذاكرة بعد فترة من الراحة.[46]
لمكافحة هذا، تم اختبار دونيبيزيل في المرضى المسنين الاصحاء حيث تبين أن زيادة الوقت الذي يقضيه في النوم REM وتحسين استعادة الذاكرة في اليوم التالي [47]
مرض الزهايمر
ان تجربة المرضى الذين يعانون من مرض الزهايمر والمزيد من اضطراب النوم في كبار السن الأصحاء. وقد أظهرت الدراسات أنه في المرضى الذين يعانون من مرض الزهايمر، انخفاض سريع في محاور الدوران. كما تبين عن أن كثافة محاور الدوران في الليلة السابقة لاختبار الذاكرة ترتبط بشكل إيجابي مع دقة استدعاء فورية.. كما تم الإبلاغ عن علاقة إيجابية بين الوقت الذي يقضيه في SWS واليوم التالي استدعاء ذاكرة السيرة الذاتية في مرضى الزهايمر.[48]
.المراجع
- Barrie, James (1911). Peter and Wendy. Hodder and Stoughton.
- Ridley, Rosalind (2016). Peter Pan and the Mind of J. M .Barrie. An Exploration of Cognition and Consciousness. Cambridge Scholars Publishing. .
- Walker, M.P.; Stickgold, R.; Alsop, D.; Gaab, N.; Schlaug, G. (2005). "Sleep-dependent motor memory plasticity in the human brain". Neuroscience. 133 (4): 911–7. doi:10.1016/j.neuroscience.2005.04.007. PMID 15964485.
- Walker, Matthew P.; Stickgold, Robert (2006). "Sleep, Memory, and Plasticity". Annual Review of Psychology. 57 (1): 139–66. doi:10.1146/annurev.psych.56.091103.070307. PMID 16318592.
- Maquet, P. (2001). "The Role of Sleep in Learning and Memory". Science. Science. 294 (5544): 1048–52. Bibcode:2001Sci...294.1048M. doi:10.1126/science.1062856. PMID 11691982.
- Stickgold, Robert; Walker, Matthew P. (2007). "Sleep-dependent memory consolidation and reconsolidation". Sleep Medicine. 8 (4): 331–43. doi:10.1016/j.sleep.2007.03.011. PMID 17470412.
- Gillin, J. Christian; Drummond, Sean P. A.; Brown, Gregory G.; Stricker, John L.; Wong, Eric C.; Buxton, Richard B. (2000). "Altered brain response to verbal learning following sleep deprivation". Nature. 403 (6770): 655–7. Bibcode:2000Natur.403..655D. doi:10.1038/35001068. PMID 10688201.
- Gais, Steffen; Born, Jan (2004). "Low acetylcholine during slow-wave sleep is critical for declarative memory consolidation". Proceedings of the National Academy of Sciences. 101 (7): 2140–2144. Bibcode:2004PNAS..101.2140G. doi:10.1073/pnas.0305404101. JSTOR 3371408.
- Robertson, Edwin M.; Pascual-Leone, Alvaro; Press, Daniel Z. (2004). "Awareness Modifies the Skill-Learning Benefits of Sleep". Current Biology. 14 (3): 208–12. doi:10.1016/j.cub.2004.01.027. PMID 14761652.
- Ramanthan, Dhakshin S.; Gulati, Tanuj S.; Ganguly, Karunesh (2014). "Sleep-Dependent Reactivation of Ensembles in Motor Cortex Promotes Skill Consolidation". PLoS Biology. 13: e1002263. doi:10.1371/journal.pbio.1002263. PMID 26382320. مؤرشف من الأصل في 29 سبتمبر 2018.
- Gulati, Tanuj; Ramanathan, Dhakshin S.; Wong, Chelsea C.; Ganguly, Karunesh (2014). "Reactivation of emergent task-related ensembles during slow-wave sleep after neuroprosthetic learning". Nature Neuroscience. 17 (8): 1107–1113. doi:10.1038/nn.3759. PMID 24997761. مؤرشف من الأصل في 26 أبريل 2020.
- Walker, Matthew P.; Brakefield, Tiffany; Morgan, Alexandra; Hobson, J.Allan; Stickgold, Robert (2002). "Practice with Sleep Makes Perfect". Neuron. 35 (1): 205–11. doi:10.1016/S0896-6273(02)00746-8. PMID 12123620.
- Maquet, Pierre; Laureys, Steven; Peigneux, Philippe; Fuchs, Sonia; Petiau, Christophe; Phillips, Christophe; Aerts, Joel; Del Fiore, Guy; et al. (2000). "Experience-dependent changes in cerebral activation during human REM sleep". Nature Neuroscience. 3 (8): 831–6. doi:10.1038/77744. PMID 10903578.
- Siesjo, Bo K. (1978). Brain Energy Metabolism. Wiley. صفحة 612. .
- Rennie, Michael J. (2007). "An introduction to the use of tracers in nutrition and metabolism". Proceedings of the Nutrition Society. 58 (4): 935–44. doi:10.1017/S002966519900124X. PMID 10817161.
- Harrison, Yvonne; Horne, James A. (2000). "Sleep Loss and Temporal Memory". The Quarterly Journal of Experimental Psychology Section A. 53 (1): 271–279. doi:10.1080/713755870.
- Drummond, S; Brown, GG (2001). "The Effects of Total Sleep Deprivation on Cerebral Responses to Cognitive Performance". Neuropsychopharmacology. 25 (5): S68–73. doi:10.1016/S0893-133X(01)00325-6. PMID 11682277.
- Huber, Reto; Felice Ghilardi, M.; Massimini, Marcello; Tononi, Giulio (2004). "Local sleep and learning". Nature. Nature. 430 (6995): 78–81. Bibcode:2004Natur.430...78H. doi:10.1038/nature02663. PMID 15184907.
- Ribeiro, Sidarta; Gervasoni, Damien; Soares, Ernesto S.; Zhou, Yi; Lin, Shih-Chieh; Pantoja, Janaina; Lavine, Michael; Nicolelis, Miguel A. L. (2004). "Long-Lasting Novelty-Induced Neuronal Reverberation during Slow-Wave Sleep in Multiple Forebrain Areas". PLoS Biology. PLoS Biology. 2 (1): e24. doi:10.1371/journal.pbio.0020024. PMID 14737198.
- Peigneux, Philippe; Laureys, Steven; Fuchs, Sonia; Collette, Fabienne; Perrin, Fabien; Reggers, Jean; Phillips, Christophe; Degueldre, Christian; et al. (2004). "Are Spatial Memories Strengthened in the Human Hippocampus during Slow Wave Sleep?". Neuron. 44 (3): 535–45. doi:10.1016/j.neuron.2004.10.007. PMID 15504332.
- Fischer, Stefan; Born, Jan (2009). "Anticipated reward enhances offline learning during sleep". Journal of Experimental Psychology: Learning, Memory, and Cognition. 35 (6): 1586–1593. doi:10.1037/a0017256.
- Guan, Zhiwei; Peng, Xuwen; Fang, Jidong (2004). "Sleep deprivation impairs spatial memory and decreases extracellular signal-regulated kinase phosphorylation in the hippocampus". Brain Research. Brain Research. 1018 (1): 38–47. doi:10.1016/j.brainres.2004.05.032. PMID 15262203.
- Datta, Subimal (2000). "Avoidance Task Training Potentiates Phasic Pontine-Wave Density in the Rat: A Mechanism for Sleep-Dependent Plasticity". The Journal of Neuroscience. 20 (22): 8607–13. PMID 11069969. مؤرشف من الأصل في 26 أبريل 2020.
- Hennevin, Elizabeth; Hars, Bernard; Bloch, Vincent (1989). "Improvement of learning by mesencephalic reticular stimulation during postlearning paradoxical sleep". Behavioral and Neural Biology. 51 (3): 291–306. doi:10.1016/S0163-1047(89)90948-5. PMID 2730495.
- Hennevin, Elizabeth; Maho, Catherine; Hars, Bernard (1998). "Neuronal plasticity induced by fear conditioning is expressed during paradoxical sleep: Evidence from simultaneous recordings in the lateral amygdala and the medial geniculate in rats". Behavioral Neuroscience. 112 (4): 839–862. doi:10.1037/0735-7044.112.4.839.
- Dang-Vu, T. T.; Desseilles, M.; Peigneux, P.; Maquet, P. (2006). "A role for sleep in brain plasticity". Developmental Neurorehabilitation. 9 (2): 98–118. doi:10.1080/13638490500138702.
- Datta, Subimal; Mavanji, Vijayakumar; Ulloor, Jagadish; Patterson, Elissa H. (2004). "Activation of Phasic Pontine-Wave Generator Prevents Rapid Eye Movement Sleep Deprivation-Induced Learning Impairment in the Rat: A Mechanism for Sleep-Dependent Plasticity". The Journal of Neuroscience. 24 (6): 1416–27. doi:10.1523/JNEUROSCI.4111-03.2004. PMID 14960614. مؤرشف من الأصل في 26 أبريل 2020.
- Wagner, Ullrich; Hallschmid, Manfred; Verleger, Rolf; Born, Jan (2003). "Signs of REM sleep dependent enhancement of implicit face memory: A repetition priming study". Biological Psychology. 62 (3): 197–210. doi:10.1016/S0301-0511(02)00125-4. PMID 12633978.
- Hatanaka, Hiroshi; Nihonmatsu, Itsuko; Tsukui, Hiroko (1988). "Nerve growth factor promotes survival of cultured magnocellular cholinergic neurons from nucleus basalis of Meynert in postnatal rats". Neuroscience Letters. 90 (1–2): 63–8. doi:10.1016/0304-3940(88)90787-2. PMID 2842704.
- Sei, Hiroyoshi; Saitoh, Daisuke; Yamamoto, Kei; Morita, Kyoji; Morita, Yusuke (2000). "Differential effect of short-term REM sleep deprivation on NGF and BDNF protein levels in the rat brain". Brain Research. 877 (2): 387–90. doi:10.1016/S0006-8993(00)02708-6. PMID 10986357.
- Kuriyama, K.; Mishima, K.; Suzuki, H.; Aritake, S.; Uchiyama, M. (2008). "Sleep Accelerates the Improvement in Working Memory Performance". Journal of Neuroscience. Journal of Neuroscience. 28 (40): 10145–50. doi:10.1523/JNEUROSCI.2039-08.2008. PMID 18829972.
- Casement, Melynda D.; Broussard, Josiane L.; Mullington, Janet M.; Press, Daniel Z. (2006). "The contribution of sleep to improvements in working memory scanning speed: A study of prolonged sleep restriction". Biological Psychology. 72 (2): 208–12. doi:10.1016/j.biopsycho.2005.11.002. PMID 16384630.
- Mirmiran, M.; Scholtens, J.; Van De Poll, N.E.; Uylings, H.B.M.; Van Der Gugten, J.; Boer, G.J. (1983). "Effects of experimental suppression of active (REM) sleep during early development upon adult brain and behavior in the rat". Developmental Brain Research. 7 (2–3): 277–86. doi:10.1016/0165-3806(83)90184-0. PMID 6850353.
- Maquet, Pierre; Schwartz, Sophie; Passingham, Richard; Frith, Christopher (2003). "Sleep-Related Consolidation of a Visuomotor Skill: Brain Mechanisms as Assessed by Functional Magnetic Resonance Imaging". The Journal of Neuroscience. 23 (4): 1432–40. PMID 12598632. مؤرشف من الأصل في 26 أبريل 2020.
- McCormick, DA (1992-01-01). "Cellular mechanisms underlying cholinergic and noradrenergic modulation of neuronal firing mode in the cat and guinea pig dorsal lateral geniculate nucleus". The Journal of Neuroscience. 12 (1): 278–89. PMID 1309574. مؤرشف من الأصل في 26 أبريل 2020.
- Hasselmo, Michael E. (1999). "Neuromodulation: Acetylcholine and memory consolidation". Trends in Cognitive Sciences. 3 (9): 351–359. doi:10.1016/S1364-6613(99)01365-0. PMID 10461198.
- Cirelli, Chiara; Gutierrez, Christina M.; Tononi, Giulio (2004). "Extensive and Divergent Effects of Sleep and Wakefulness on Brain Gene Expression". Neuron. 41 (1): 35–43. doi:10.1016/S0896-6273(03)00814-6. PMID 14715133.
- Ahn, Sohyun; Ginty, David D.; Linden, David J. (1999). "A Late Phase of Cerebellar Long-Term Depression Requires Activation of CaMKIV and CREB". Neuron. 23 (3): 559–68. doi:10.1016/S0896-6273(00)80808-9. PMID 10433267.
- Walker, Matthew P.; Stickgold, Robert (2005). "It's Practice, with Sleep, that Makes Perfect: Implications of Sleep-Dependent Learning and Plasticity for Skill Performance". Clinics in Sports Medicine. 24 (2): 301–17, ix. doi:10.1016/j.csm.2004.11.002. PMID 15892925.
- Mednick, Sara; Nakayama, Ken; Stickgold, Robert (2003). "Sleep-dependent learning: A nap is as good as a night". Nature Neuroscience. 6 (7): 697–8. doi:10.1038/nn1078. PMID 12819785.
- Folkard, S.; Tucker, P (2003). "Shift work, safety and productivity". Occupational Medicine. 53 (2): 95–101. doi:10.1093/occmed/kqg047. PMID 12637593.
- Morin, Charles; Gramling, Sandy (1989). "Sleep patterns and aging: Comparison of older adults with and without insomnia complaints". Psychology and Aging. 4 (3): 290–294. doi:10.1037/0882-7974.4.3.290. PMID 2803622.
- Rauchs, Géraldine; Schabus, Manuel; Parapatics, Silvia; Bertran, Francoise; Clochon, Patrice; Hot, Pascal; Denise, Pierre; Desgranges, Beatrice; Eustache, Francis (2008). "Is there a link between sleep changes and memory in Alzheimer's disease?". Learning and Memory. 19 (11): 1159–1162. doi:10.1097/WNR.0b013e32830867c4. PMID 18596620.
- Petit, Dominique; Gagnon, Jean-François; Fantini, Maria; Ferini-Strambi, Luigi; Montplaisir, Jacques (2004). "Sleep and quantitative EEG in neurodegenerative disorders". Journal of Psychosomatic Research. 56 (5): 487–496. doi:10.1016/j.jpsychores.2004.02.001. PMID 15172204.
- Darchia, Nato; Campbell, Ian; Feinberg, Irwin (2003). "Rapid eye movement density is reduced in the normal elderly". Sleep. 26 (8): 973–977. PMID 14746377.
- Scullin, Michael (2013). "Sleep, Memory, and Aging: The Link Between Slow-Wave Sleep and Episodic Memory Changes from Younger to Older Adults". Psychology and Aging. 28 (1): 105–114. doi:10.1037/a0028830. PMID 22708533.
- Schredl, M; Weber, B; Leins, M; Heuser, I (2001). "Donepezil-induced REM sleep augmentation enhances memory performance in elderly, healthy persons". Experimental Gerontology. 36 (2): 353–361. doi:10.1016/s0531-5565(00)00206-0. PMID 11226748.
- Rauchs, G; Piolino, P; Bertran, F; de La Sayette, V; Viader, F; Eustache, F; Desgranges, B (2013). "Retrieval of recent autobiographical memories is associated with slow-wave sleep in early AD". Frontiers in Behavioral Neuroscience. 7 (114): 1–11. doi:10.3389/fnbeh.2013.00114. PMID 24065896.