
الشحميّات السفينجوليّة (بالانجليزية: Sphingolipid) هي فئة من فئات الدّهون التي تحتوي بشكل أساسيّ على أساس (دعامة)من القواعد السفينجوليّة، مجموعة من الكحوليّات الأليفاتية (سلسلة مفتوحة ) الأمينيّة التي تتضمّن سفينغوزين، تمّ اكتشافها في سبعينيات القرن الثامن عشر، عندما تمّ استخراج خلاصات (افرازات) الدماغ. سُميّت بذلك نسبة إلى أسطورة "أبو الهول العظيم" بسبب طبيعتها الغامضة.[1] وتلعب دوراً مهمهاً في عملية نقل الإشارات والتَعَرُّفٌ الخَلَوِيّ. الخلل في العمليات الايضية لشحميات السيفنجولية أو ما يُسمّى ب " الشُحامَاتٌ السفينغولية " أنّ لها تأثيرًا معيّنًا على الأنسجة العصبية.
ويطلق على الشحميّات السفنجولية التي تحتوي على مجموعة كربونيّة واحدة وذرة هيدروجين متّصلة ب " السيراميد ". وهناك بعض المجموعات الكربونيّة الشائعة التي ترتبط بالقواعد السفينجوليّة، مثل الفسفوكولاين والتي يُسفَر عنها ما يسمى بـ"سيفنجومالين "، و مُختلَف أنواع السكر الأحادي أو الثنائي الذي يُسفِر عن " السيريبروسيد (cerebrosides)" و " الجلوبوسيد "، ويُطلق عليها مجتمعةً " الشَّحْمِيَّاتُ السِّفِنْغُولِيَّةُ السُّكَّرِيَّة " الهيكل ( البُنيّة ) العام للشحميات السيفنجولية
الهيكل العام
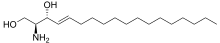
هي عبارة عن قواعد طويلة السلسلة، وتُعرف عادة بقواعد السفينغويد، وهي أول منتج غيرانتقالي في عملية الصناعة (دي نوفو) للشحميّات السفينغولية في كلا الخميرة والثدييات. تُسمّى هذه المركبات على وجه التحديد ب السفينغوزين النبايتة (فايتوسفينغوزين) و ثنائي هيدروسفينغوسين (أيضًا تُسمّى بسفينغانين، على الرّغم من أن هذا المصطلح غير مشهور)، هي عبارة عن مركّبات تحتوي على 18 كربون، مع وجودبعض المركبات التي تحتوي على 20كربون.[2] السيراميد والشحميّات السفينغوليّة السُّكّرية تمثل مشتقّات الأسل النيتروجينية من هذه المركبات. الجسم الهيكلي (السلسلة المفتوحة) لل سفينغوزين يرتبط بـ(رابطة تحتوي على ذرة الاكسجين) مع المجموعات الرأسيّة المشحونة غالباً، مثل إيثانولامين، أو السيرين، أو الكولين. الجسم الهيكليّ أيضا يرتبط برابطة أميديّة مع مجموعة الأسيل، مثل الدهنيّات.
الأنواع
البسيطة، وتُشكل الشحميّات السفنجوليّة البسيطة التي تتضمّن القواعد السيفنجولية والسيراميد، تمثل المنتجات الأوليّة في مسارات العمليّات البنائيّة للشحميّات السفنجوليّة. •القواعد السيفنجوليّة هي اللبنة الأساسيّة في بناء الشحميّات السيفنجوليّة[3][4]، ويعد " سيفنجوزين " و " ثنائي هايدروسيفنجوزين " القواعد السيفنجوليّة الأساسية في الثدييات، بينما في الخميرة فإنّ " ثنائي هايدروسيفنجوزين " و " فايتوسيفنجوزين " هما القواعد السيفنجوليّة الرئيسة. وقد يمكن فسفرة هذه القواعد الثلاثة. •السيراميد، كصنف عامّ، هي قواعد سيفنجوليةّ تحتوي على أسيتل النيتروجين، ولا تحتوي على أيّ مجموعة رأسيّة إضافيّة. •ثنائي هايدروسيرمايد، تُكوّن عن طريق أستلة (إضافة مجموعة اسيتل النيتروجين )القاعدة السيفنجولية " ثنائي هايدروسيفنجوزين "، وقد يتواجد في الثديّيات والخميرة. •يتمّ إنتاج السيراميد في الثدييات عن طريق إزالة الإشباع ل" ثنائي هايروسيراميد " بواسطة الأنزيم DES1. و يُعَدّ السيراميد جزيئًا نشيط حيويّا بشكل عالي؛ إذ يمكن أيضاً فسفرته ليكون ceramide-1-phosphate.(سيراميد أحادي الفسفات) •فايتوسيراميد، ويتم إنتاجه في الخميرة عن طريق عملية الهيدروكسلة (أضافة ال(OH ل " ثنائي هايروسيراميد" على الكربونة الرابعة.
المعقدة، وتُنتج من خلال إضافة مجموعة رأسية لـ:"السيراميد" أو "الفايتوسيراميد".
•سيفنغوميلين، تحتوي على جزيء الفسفوركولين أو الفسفورثانوألامين مرتبط مع المجموعة الهيدروكسية رقم واحد 1-hydroxy group في السيراميد برابطة إسترية.
•جلايكوسفينجوليبيد، وهي تمثل ارتباط جزيئات السكر بمجوعة الهيدروكسيل الاولى للسيراميد برابطة بيتا جلايكوسيد β-glycosidic .
•سيرِيبْرُوزيد، يحتوي على جزيء منفرد من الغلوكوز أوالغلاكتوز مرتبط على المجموعة الهيدروكسيّة رقم واحد.
•الكبريتيدات، وهو عبارة عن سيرابروزيد مُكبرت.
•غانغلوزيد، يحتوي على الأقلّ على ثلاث جزيئات سكر، واحدة منهم يجب أن تكون حمض السياليك.
•انيستول-سيراميد، مشتق من الفايتوسيراميد، يتم إنتاجه في الخميرة. يتضمّن على انيستول فسفورسيراميد، مانوز انيستول فسفورسيراميد ومانوز ثنائي انيستول فسفورسيراميد.
العمليّات الأيضيّة للشحميّات السيفنجولية في الثدييات تبدأ عملية إنشاء الشحميّات السيفنجولية بتكوين المركب (3_ كيتو ثنائي هايدروسيفنجوزين ) عن طريق الأنزيم " سيرين بالميتوترانسفيريز " ( SPLT )[5]. يُعتبَر كلا المركبان ال ( بالتيمول التميم ) و ال (سيرين) المواد المستهدفة لهذا التفاعل[6]، غير أنّ هنالك بعض الدراسات التي قامت بتأكيد فعاليّة الأنزيم نحو بعض الأنواع من الدّهون الأسيلية التميمة (Acyl-CoA )والحموض الأمينيّة البديلة[7]. وقد تمّت إعادة النظر في الآونة الأخيرة في التنوّع في القواعد السيفنجولية[8]. و المرحلة التالية من عملية إنشاء الشحميات السيفنجولية باختزال المركب (3_ كيتو ثنائي هايدروسيفنجوزين ) إلى ثنائي هايدرو سينفوجوزين الذي بدوره يتم أسيَلَته إلى ثنائي هايدرو سيراميد عن طريق الأنزيم ثنائي هايدرو_سيراميد سينثاز مثل لاس1ب أو لاس2ب، ثم تتمّ إزالة الإشباع عن المركب ثنائي هايدرو سيراميد[9] ليتكون المركب سيراميد[10].
قد يتعرض مركب السيراميد لأكثر من تفاعل، قد تتمّ فسفرته بواسطة الأنزيم سيراميد كيناز ليتكون المركب ( سيراميد_1_ فوسفات)، أو قد يرتبط بالغليكوزيل بواسطة الأنزيم غليكوز سيراميد سينثاز أو غلاكتوز سيراميد سنثاز، بالإضافة إلى ذلك قد يتمّ تحويله إلى سيفنجوميلين بإضافة المجموعة الرأسيّة فسفوكولين بواسطة سيفنجوميلين سينثاز، وفي هذه العمليّة يُنتج المركّب ثنائي الأسيل الجليسرول. أخيراً يُمكِن أن يتمّ تكسير مركب السيراميد ليكون سيفنجوزين بواسطة الأنزيم سيراميديز. و يمكن فسفرة السيفنجوزين لتكوين (سيفنجوزين_1_فوسفات)، وقد تتمّ إزالة الفسفرة عنه وإعادته إلى سيفنجوزين.[11]
أما بالنسبة عن مسارات تفكّك المركّبات الناتجة عن العمليّات الأيضيّة فقد تؤدي إلى رجوع المركب سيراميد. تتحلل الدهون الجلايكوسفنجولية المعقّدة إلى جلايكوسيراميد(تحتوي على سكر الغلوكوز) أو جلاكتوسيراميد(تحتوي على سكر الغلاكتوز)، وتتحلل هذه المركبات مرة أخرى بواسطة الأنزيم بيتا جلوكوسيديز وجلاكتوسيديز لتتم إعادة إنتاج السيراميد. وبطريقة مشابهة، يتكسّر سيفنجوميلين إلى سيراميد بواسطة سيفنجوميلينيز. الطريقة الوحيدة التي يُكوّن فيها الشحمات السيفنجولية مركبات غير الشحمات السيفنجولية هي بواسطة الأنزيم (سيفنجوزين_1_فوسفاتليز)، هذا يُنتج ايثانول أمين وهيكساديكنال.[12]

وظائف الشحميات السيفنجولية في الثدييات
أما عن وظيفة الشحميات السيفنجولية الأكثر شيوعا في الثدييات هو حماية سطح الخليّة من عوامل البيئة الضارّة، مِن خلال تشكيل وريقة خارجيّة مقاومة ميكانيكياً ومستقرّة كيميائياً على الطبقة الدّهنيّة المزدوجة للغشاء البلازميّ. وهناك بعض الدّهون السيفنجولية السكّريّة المُعقّدة التي تشارك في عمليّة التعرّف الخلويّ والإشعار الخلويّ. يعتمد التعرّف الخلويّ بشكل أساسيّ على الطبيعة الفيزيائيّة للشحميّات، بينما يعتمد الإشعار الخلويّ على مجموعة من التفاعلات المعينة بين الغليكان على الشحميات السيفنجوليّة ودهون مشابهة موجودة على الخلايا المجاورة والبروتينات. في الآونة الأخيرة، تم إثبات دور بعض الشحمياّت المستقلبة البسيطة مثل السيراميد وال (سيفنجوزين_1_فوسفات) باعتبارها وسيطا مهما في عملية الإستشعار الخلويّ(انتقال الاشارات) التسلسليّ، مشاركة في الاستماتة ( الموت الخلوي المبرمج )، التكاثر الخلوي، استجابات الإجهاد، النخر، الالتهابات، التهام الذات، الشيوخة والتمايز.[13][14][15][16][17][18][19] تتجمع الدّهون سيراميدية القاعدة ذاتياً جنبا إلى جنب في أغشية الخلية مكوّنة طورا منفصلا أقلّ مُيوعة من الدهون الفسفوريّة. هذه الشحميّات السيفنجولية ذات النطاق الصّغير أو ما يُسمّى ب " النطاق الدهني " تمّ اقتراحها في الأصل كنوع من البروتينات الغشائيّة ضمن الناقلات الغشائيّة في المسارات الخلويّة. في الوقت الحاضر، تتركّز معظم الأبحاث على عمليّات التنظيم في الإسشعار(انتقال الاشارات).[20]
تبدأ عملية إنتاج الشحميّات السيفنجولية في الشبكة الإندوبلازمية، وتكتمل في أجسام جولجي، تتمّ تقوية هذه الدّهون في الغشاء البلازمي والدخلول ( اندوسوم )؛ حيثُ تنفّذ العديد من مهامّها ووظائفها الخلويّة. يتم النقل عبر الحويصلات والناقلات الأحادية في العصارة الخلوية[21]. فعلياً، الشحميّات السيفنجولية غير موجودة في الشبكة الأندوبلازمية والميتوكندريا، ولكنّها تُشكّل حوالي (20-35) جزيء مولي من الدهون في الغشاء البلازمي.[22] ومن خلال التجارب الذي خضعت لها الحيوانات، فقد لوحظ أن تغذيتها بالشحميّات السيفنجولية يؤدي إلى منع تسرطن القولون، والتقليل من الكوليسترول قليل الكثافة (السيئ)، ويزيد من الكوليسترول عالي الكثافة (الجيد).
الشحميّات السيفنجوليّة في الخميرة
نتيجة للتعقيد الهائل في أنظمة الثدييات، فإنّ الخميرة نموذج مناسب للعمل عليها وتجربة مسارات جديدة؛ لأنّها كائنات وحيدة الخليّة، فمن السهل السيطرة على جيناتها مقارنة بخلايا الثدييات، ويمكن زراعة خلايا الخميرة باحداث تغيرات جينية غير قاتلة على جزيئاتها الذرية ليتمّ استنساخ خلايا ذات خواصّ مُعيّنة. أكثر أنواع الخمائر استخداماً هما خميرة البيرة (Saccharomyces cerevisiae) وخميرة سكيراء الجعة (Schizosaccharomyces pombe)، بالإضافة إلى الخميرة المَرَضيّة المبيضة البيضاء (Candida albicans). بالإضافة إلى أهميّة الوظيفة الهيكليّة للشحميّات السيفنجوليّة المُعقّدة، فإنّ القواعد السيفنجوليّة فايتوسيفنجوزين وثنائي هايدرو سيفنجوزين تلعب دوراً حيويًّا مهمًّا في خميرة البيرة (Saccharomyces cerevisiae ). و هذه التأثيرات تتضمّن تنظيم عملية التقام الخلوي، وعمليّة تحلّل البروتين المعتمدة على اليوبيكويتين ( وبالتالي تنظيم عمليّة امتصاص الموادّ الغذائيّة [23]، وديناميكية الهيكل الخلوي، الدورة الخلوية، الترجمة الجينية، تعديل البروتينات المتخصصة في الترجمة والاستجابة للإجهاد الحراري[24]. بالإضافة إلى معالجة العمليات الأيضية للشحميات السيفنجولية بواسطة (فسفوات ثنائي انيستول_4،5_بايفوسفات) المرسلة عبر Slm1p و Slm2p ، وقد تمّ أيضاً وصف الكالسينيورين مؤخراً[25]. وقد لوحظ ظهور التفاعلات المعتمدة على مراحل الركائز بين إنتاج الشحميات السيفنجوليّة المعقدة ودورة الأنزيم (فسفات ثنائي انيستول_4_فوسفات) بواسطة فسفور ثنائي انيستول كيناز (phosphatidylinositol kinase Stt4p )، و مفسفر الدهون ( lipid phosphatase Sac1p ).[26]
الشحميّات السيفنجوليّة في النباتات
تحتوي النباتات على مجموعة أكبر وأكثر تنوعًّا من الشحميّات السيفنجولية مقارنة بالحيوانات والخميرة الاضطرابات 9 هناك العديد من الاضطرابات التي قد تحصل في العمليّات الأيضيّة للشحميّات السيفنجولية، ويُطلق عليها الشحامات السيفنجولية. ومن أهمّها داء نيمان بيك، داء فابري، داء كرابه، مرض جوشر، داء تاي زاكس (الشحام السفينغولي الطفلي) ومرض حثل المادة البيضاء المتبدل اللون. هذه الأمراض عادة ما تنتقل وراثياً على الجينات الصبغيّة الجسديّة المتنحية، لكن مرض فابري على وجه خاصّ ينتقل على الجين X. قد تحدث هذه الاضطرابات في ما يقارب نسبته واحد لكل عشرة آلاف، وقد يزداد حدوثها عند مجتمعات معيّنة من السكّان مثل اليهود الأشكناز، وتُعد المُعالَجَة بالاِسْتِعاضَة الأنزيميّة وسيلة ملائمة لعلاج مرضيّ فابري وجوشر، والأشخاص المصابون بمثل هذا الخلل يعيشون بشكل جيّد حتى البلوغ، بينما هناك أنواع أخرى قد تكون قاتلة في عمر 1-5 سنوات، وقد يكون التقدّم بالمرض معتدلا لليافعين والبالغين.
المراجع
- Chun, J.; Hartung, H.P. (2010). "Mechanism of Action of Oral Fingolimod (FTY720) in Multiple Sclerosis". Clin. Neuropharmacol. 33 (2): 91–101. doi:10.1097/wnf.0b013e3181cbf825.
- Product page at Sigma Aldrich - تصفح: نسخة محفوظة 18 سبتمبر 2011 على موقع واي باك مشين.
- Dickson, Robert C (2008). "New insights into sphingolipid metabolism and function in budding yeast". J Lipid Res. 49: 909–921. doi:10.1194/jlr.R800003-JLR200.
- Bartke, N; Hannun, Y. (2009). "Bioactive sphingolipids: metabolism and function". J Lipid Res. 50: S91–6. doi:10.1194/jlr.R800080-JLR200.
- Merrill. "Characterization of serine palmitoyltransferase activity in Chinese hamster ovary cells." Biochim Biophys Acta (1983) 754(3):284-91.
- Merrill and Williams. "Utilization of different fatty acyl-CoA thioesters by serine palmitoyltransferase from rat brain". Journal of Lipid Research (1984) 25 (2): 185-188.
- Zitomer NC, Mitchell T, Voss KA, Bondy GS, Pruett ST, Garnier-Amblard EC, Liebeskind LS, Park H, Wang E, Sullards MC, Merrill AH Jr, Riley RT. "Ceramide Synthase Inhibition by Fumonisin B1 Causes Accumulation of 1-Deoxysphinganine: A Novel Category of Bioactive 1-Deoxysphingoid Bases And 1-Deoxydihydroceramides Biosynthesized By Mammalian Cell Lines And Animals". Journal of Biological Chemistry (2009) 284 (8): 4786-4795.
- Pruett et al. "Biodiversity of sphingoid bases ("sphingosines") and related amino alcohols". Journal of Lipid Research. (2008) 49:1621-1639.
- Pewzner-Jung et al. "When do Lasses (longevity assurrance genes) become CerS (ceramide synthases)?: insights into the regulation of ceramide synthesis". Journal of Biological Chemistry. (2006) 281, 25001-25005.
- Causeret et al. "Further characterization of rat dihydroceramide desaturase: tissue distribution, subcellular localization, and substrate specificity". Lipids. (2005) 35:1117-1125.
- Reviewed in Hannun and Obeid. "Principles of bioactive lipid signalling: lessons from sphingolipids". Nature Reviews Molecular Cell Biology. (2008) 9, 139-150.
- Bandhuvulua & Saba. "Sphingosine-1-phosphate lyase in immunity and cancer: silencing the siren". Trends in Molecular Medicine. (2007) 13:210-217.
- Hannun YA, Obeid LM (July 2002). "The Ceramide-centric universe of lipid-mediated cell regulation: stress encounters of the lipid kind". J. Biol. Chem. 277 (29): 25847–50. doi:10.1074/jbc.R200008200. PMID 12011103. مؤرشف من الأصل في 31 يناير 2009.
- Spiegel S, Milstien S (July 2002). "Sphingosine 1-phosphate, a key cell signaling molecule". J. Biol. Chem. 277 (29): 25851–4. doi:10.1074/jbc.R200007200. PMID 12011102. مؤرشف من الأصل في 31 يناير 2009.
- Lavieu, G; Scarlatti, F; Sala, G; Carpentier, S; Levade, T; Ghidoni, R; Botti, J; Codogno, P (2006). "Regulation of autophagy by sphingosine kinase 1 and its role in cell survival during nutrient starvation". J Biol Chem. 281 (13): 8518–27. doi:10.1074/jbc.M506182200.
- Venable, M. E.; Lee, J. Y.; Smyth, M. J.; Bielawska, A.; Obeid, L. M. (1995). "Role of ceramide in cellular senescence". J. Biol. Chem. 270: 30701–30708. doi:10.1074/jbc.270.51.30701. PMID 8530509.
- Hetz, C. A.; Hunn, M.; Rojas, P.; Torres, V.; Leyton, L.; Quest, A. F. (2002). "Caspase-dependent initiation of apoptosis and necrosis by the Fas receptor in lymphoid cells: onset of necrosis is associated with delayed ceramide increase". J. Cell Sci. 115: 4671–4683. doi:10.1242/jcs.00153.
- Snider, AJ; Orr Gandy, KA; Obeid, LM (2010). "Sphingosine kinase: Role in regulation of bioactive sphingolipid mediators in inflammation". Biochimie. 92 (6): 707–15. doi:10.1016/j.biochi.2010.02.008.
- Brown DA, London E (June 2000). "Structure and function of sphingolipid- and cholesterol-rich membrane rafts". J. Biol. Chem. 275 (23): 17221–4. doi:10.1074/jbc.R000005200. PMID 10770957. مؤرشف من الأصل في 01 يونيو 2009.
- Futerman AH (December 2006). "Intracellular trafficking of sphingolipids: relationship to biosynthesis". Biochim. Biophys. Acta. 1758 (12): 1885–92. doi:10.1016/j.bbamem.2006.08.004. PMID 16996025.
- van Meer G, Lisman Q (July 2002). "Sphingolipid transport: rafts and translocators". J. Biol. Chem. 277 (29): 25855–8. doi:10.1074/jbc.R200010200. PMID 12011105. مؤرشف من الأصل في 31 يناير 2009.
- "Sphingolipids in Food and the Emerging Importance of Sphingolipids to Nutrition". July 15, 2013. مؤرشف من الأصل في 09 ديسمبر 2019.
- Chung, N (2001). "Phytosphingosine as a specific inhibitor of growth and nutrient import in Saccharomyces cerevisiae". J Biol Chem. 276 (38): 35614–21. doi:10.1074/jbc.m105653200.
- Cowart; Obeid (2007). "Yeast sphingolipids: recent developments in understanding biosynthesis, regulation, and function". Biochim Biophys Acta. 1771 (3): 421–31. doi:10.1016/j.bbalip.2006.08.005.
- Dickson, RC (2008). J Lipid Res. 49 (5): 909–21.
- Brice; Cowart (2009). J Biol Chem.