القشرة الحزامية (باللاتينية: Cortex cingularis) هي قسم من الدماغ يقع في الجانب الأنسي للقشرة الدماغية. تتضمن القشرة الحزامية كامل التلفيف الحزامي (باللاتينية: gyrus cinguli)، الذي يقع مباشرةً أعلى الجسم الثفني، ويستمر حتى الثلم الحزامي. تُعتبر القشرة الحزامية عادةً جزءًا من الفص الحوفي.
| قشرة حزامية | |
|---|---|
 السطح الإنسي لنصف الكرة المخية الأيسر، مع إبراز التلفيف الحزامي والتلم الحزامي.
| |
| تفاصيل | |
| نوع من | تلفيف[1] |
| معرفات | |
| ترمينولوجيا أناتوميكا | 14.1.09.231 |
| FMA | 62434 |
| UBERON ID | 0002967 |
| ن.ف.م.ط. | A08.186.211.577.330 |
| ن.ف.م.ط. | D006179 |
[ ] |
|


تتلقى المدخلات من المهاد والقشرة الحديثة، وتنقلها إلى القشرة الشمية الداخلية عبر الحزام. تعتبر جزءًا هامًا من الجهاز الحوفي المسؤول عن تشكيل العاطفة ومعالجتها، والتعلم، والذاكرة. تجعل توليفة تلك الوظائف الثلاث من التلفيف الحزامي مؤثرًا كبيرًا في ربط المخرجات التحفيزية للسلوك (مثلًا، يحفز نشاط معين استجابةً عاطفية إيجابية ما، فتؤدي في النتيجة النهائية إلى التعلم). يساهم هذا الدور بزيادة أهمية القشرة الحزامية في اضطرابات مثل الاكتئاب والفصام. ويلعب دورًا في الوظيفة التنفيذية والتحكم بالتنفس أيضًا.[2][3][4][5][6][7][8]

الاصطلاح
من اللاتينية (cingulātus) «مطوق أو محزم».
البنية
تقسم بناءً على التصميم الخلوي المخي إلى باحات برودمان 23، و24، و26، و29، و30، و31، و33. يُشار إلى الباحات 26، و29، و30 عادةً على أنها باحات خلف الشريط (شريط الجسم الثفني).

القشرة الحزامية الأمامية
- مقالة مفصلة: قشرة حزامية أمامية
تتوافق هذه القشرة مع باحات برودمان 24، و32، و33 و(إل إيه) «الباحات الحوفية العليا» الخاصة بتصميم كونستاتين فون ايكونومو وبايلي وفون بونين الخلوي. تستمر للأمام من خلال باحة الرُكبة (باحة برودمان 25)، التي تقع أسفل رُكبة الجسم الثفني. تعتبر لا محببة من ناحية التصميم الخلوي. ولها جزء تلفيفي وجزء ثلمي. قد تقسم القشرة الحزامية الأمامية بشكل إضافي في القشرة الحزامية الأمامية المجاورة للرُكبة (قرب الرُكبة) والقشرة الحزامية المتوسطة. تستقبل القشرة الحزامية الأمامية بشكل أساسي محاورها الواردة من نوى المهاد داخل الصفائح ونوى المهاد الناصفة (انظر المهاد). تستقبل النوى الأمامية أعصابًا حلمية مهادية واردة. تستقبل العصبونات الحلمية محاور من مِرفد الحصين. تعرف كافة أشكال الدائرة العصبية في الجهاز الحوفي باسم «دائرة بابيز». ترسل القشرة الحزامية الأمامية محاور إلى النوى الأمامية عبر الحزام إلى باحات برودمان الحوفية المختلفة. تشترك القشرة الحزامية الأمامية في عمليات اكتشاف الأخطاء والصراعات.[9]
القشرة الحزامية الخلفية
تتوافق هذه القشرة مع باحات برودمان 23 و31 و(إل بّي) الخاصة بتصميم فون ايكونومو وبايلي وفون بونين الخلوي. بنيتها الخلوية حبيبية. وتستمر إلى الخلف بالقشرة خلف الشريط (باحة 29). توجد الباحة 31 الحبيبية بشكل ظهري. تستقبل القشرة الحزامية الخلفية جزءًا كبيرًا من المحاور الواردة من النوى السطحية (أو النوى الظهرية الجانبية العليا بشكل خاطئ) للمهاد (انظر المهاد)، والتي تستقبل بذاتها محاور من مِرفد الحصين. وتكرر إلى حد ما دائرة بابيز. تستقبل أيضًا أعصابًا ورادة من مِرفد الحصين. حُددت القشرة الحزامية الخلفية منخفضة الاستقلاب (مع 18إف إف دي جي بّي إي تي) في مرض الزهايمر.
مدخلات التلفيف الحزامي الأمامي
كشفت تجربة تقفي الأثر العكسي التي أجريت على قرود المكاك اتصال النوى الأمامية البطنية (في إيه) والنوى الوحشية البطنية (في إل) للمهاد بالباحات المحركة الخاصة بالثلم الحزامي. تُقسم مناطق خلف الشريط (باحات برودمان 26، و29، و30) الخاصة بالتلفيف الحزامي إلى ثلاثة أجزاء: مثلًا، القشرة خلف الشريط الحبيبية إيه، والقشرة خلف الشريط الحبيبية بي، والقشرة خلف الشريط سيئة التحبب. يرسل تشكل الحصين إسقاطات كثيفة القشرة خلف الشريط الحبيبية إيه وبي وإسقاطات أقل إلى والقشرة خلف الشريط سيئة التحبب. يرسل خلف المرفد إسقاطات إلى القشرة خلف الشريط الحبيبية إيه وبي وإلى القشرة خلف الشريط سيئة التحبب. يرسل المرفد الظهري إسقاطات إلى القشرة خلف الشريط الحبيبية بي، بينما يرسل المرفد البطني إسقاطات إلى القشرة خلف الشريط الحبيبية إيه. ترسل الأقسام الذنبية من القشرة الشمية الداخلية إلى القشرة خلف الشريط سيئة التحبب.[10][11]
مخرجات القشرة الحزامية الأمامية
يرسل التلفيف الحزامي المنقاري (باحة برودمان 32) إسقاطات إلى التلفيف الصدغي المنقاري العلوي، والقشرة الجبهية الحجاجية المتوسطة والقشرة أمام الجبهية الوحشية. يرسل الحزام الأمامي البطني (باحة برودمان 24) إسقاطات إلى القشرة الجزيرية الأمامية، والقشرة قبل المحركة (باحة برودمان 6)، وباحة برودمان 8، والباحة المحيطة الأنفية، والقشرة الجبهية الحجاجية (باحة برودمان 12)، النوى الجانبية القاعدية الخاصة باللوزة الدماغية، والقسم المنقاري من الفصيص الجداري السفلي. أظهر حقن مزيج راصة جرثومة القمح وبيروكسيداز الفجل الحار ضمن التلفيف الحزامي الأمامي عند القطط وجود اتصالات متبادلة بين هذا التلفيف والجزء المنقاري للنواة المهادية الخلفية الوحشية والنهاية المنقارية للوسادة المهادية. يستقبل خلف المرفد إسقاطات من القشرة خلف الشريط سيئة التحبب والقشرة خلف الشريط الحبيبية إيه وبي. يستقبل مجاور المرفد إسقاطات من القشرة خلف الشريط سيئة التحبب والقشرة خلف الشريط الحبيبية إيه. تحصل الأقسام الذنبية والوحشية من القشرة السمعية الداخلية على إسقاطات من القشرة خلف الشريط سيئة التحبب، بينما يستقبل القسم الذنبي المتوسط من القشرة السمعية الداخلية إسقاطات من القشرة خلف الشريط الحبيبية إيه. ترسل القشرة خلف الشريط سيئة التحبب إسقاطات إلى القشرة المحيطة الأنفية. ترسل القشرة خلف الشريط الحبيبية إيه إسقاطات إلى أمام المرفد المنقاري.[12][13]
مخرجات التلفيف الحزامي الخلفي
ترسل القشرة الحزامية الخلفية (باحة برودمان 23) إسقاطات إلى القشرة أمام الجبهية الظهرية الوحشية (باحة برودمان 9)، والقشرة أمام الجبهية الأمامية (باحة برودمان 10)، والقشرة الجبهية الحجاجية (باحة برودمان 11)، والتلفيف المجاور للحصين، والقسم الخلفي من الفصيص الجداري السفلي، وأمام المرفد، والثلم الصدغي العلوي والمنطقة خلف الشريط.[14]
تتصل القشرة خلف الشريط والقسم الذنبي من القشرة الحزامية مع القشرة أمام الجبهية المنقارية عبر الحزم الحزامية عند قرود المكاك. وُجد اتصال متبادل بين القشرة الحزامية الخلفية البطنية مع القسم الذنبي من الفص الجداري الخلفي عند قرود المكاك الريسوسي. تتصل أيضًا القشرة الجدارية الخلفية المتوسطة مع القسم البطني الخلفي الفرعي من الثلم الحزامي.[15]
اتصالات أخرى
يتصل الحزام الأمامي مع الحزام الخلفي عند الأرانب على الأقل. يتصل التلفيف الحزامي الخلفي مع القشرة خلف الشريط، ويعتبر هذا الاتصال جزءًا من المرفد الظهري للجسم الثفني. ترسل التلافيف الحزامية الأمامية والخلفية والقشرة خلف الشريط إسقاطات إلى المرفد وأمام المرفد.[16]
الأهمية السريرية
الفصام
وجد تاكاشي وآخرون (2003) باستخدام عملية التصوير بالرنين المغناطيسي ثلاثي الأبعاد لقياس حجم التلفيف الحزامي الأمامي المنقاري (التلفيف الحزامي المجاور للرُكبة) أن التلفيف الحزامي الأمامي المنقاري مسيطر بشكل أكبر عند الإناث أكثر من الذكور (الأصحاء)، لكن لم يوجد هذا الاختلاف الجنسي عند الأشخاص المصابين بالفصام. لدى المصابين بالفصام حجم أصغر من التلفيف الحزامي المجاور للركبة منه عند عينات الأشخاص المرجعية.[17]
درس هازنيدار وآخرون (2004) معدل استقلاب الغلوكوز في التلفيف الحزامي الأمامي والخلفي عند المصابين بالفصام، واضطراب الشخصية فصامي النوع (إٍس بّي دي) وقارنوهم بمجموعة أشخاص مرجعية. كان معدل استقلاب الغلوكوز أقل في التلفيف الحزامي الأمامي الأيسر والتلفيف الحزامي الخلفي الأيمن عند المصابين بالفصام مقارنةً مع المجموعة المرجعية. وعلى الرغم من توقع إظهار المصابين باضطراب الشخصية فصامي النوع لمعدل استقلاب غلوكوز تقريبًا ما بين معدل الأفراد المصابين بالفصام والمجموعة المرجعية، كان لديهم في الواقع معدل استقلاب غلوكوز أعلى في التلفيف الحزامي الخلفي الأيسر. قُلل حجم التلفيف الحزامي الأمامي الأيسر عن المصابين بالفصام مقارنةً مع المجموعة المرجعية، لكن لم يكن هناك أي اختلاف بين المصابين باضطراب الشخصية فصامي النوع والمصابين بالفصام. تُظهر هذه النتائج أن الفصام واضطراب الشخصية فصامي النوع هما اضطرابين مختلفين عن بعضهما.[18]
أظهرت دراسة حجم المادة الرمادية والبيضاء في التلفيف الحزامي الأمامي عند المصابين بالفصام وأقارب الدرجة الأولى والثانية الأصحاء عدم وجود اختلاف ذو أهمية في حجم المادة البيضاء عند المصابين بالفصام وأقربائهم الأصحاء. ومع ذلك اكتُشف اختلاف ذو أهمية في حجم المادة الرمادية، إذ كان حجمها أقل عند المصابين بالفصام مقارنةً بأقارب الدرجة الثانية، دون أن تكون كذلك مقارنةً مع أقارب الدرجة الأولى. امتلك كلا الأفراد المصابين بالفصام وأقاربهم من الدرجة الأولى الأصحاء حجمًا أصغر للمادة الرمادية من أقارب الدرجة الثانية الأصحاء. يبدو أن الجينات هي المسؤولة عن انخفاض حجم المادة الرمادية عند المصابين بالفصام.[19]
أجرى فوجيوارا وآخرون (2007) تجربة ربطوا فيها حجم التلفيف الحزامي الأمامي عند المصابين بالفصام مع أدائهم الوظيفي في الإدراك الاجتماعي، والمرضيات النفسية والعواطف مع مجموعة مرجعية. كلما كان حجم التلفيف الحزامي الأمامي أصغر، كانت مستويات الوظيفة الاجتماعية أقل والمرضيات النفسية أعلى عند المصابين بالفصام. وُجد أن التلفيف الحزامي الأمامي أصغر بشكل ثنائي عند المصابين بالفصام مقارنةً مع مجموعة مرجعية. لا يوجد اختلاف في اختبارات القدرات العقلية (آي كيو) والقدرة البصرية الإدراكية الأساسية مع التنبيه الوجهي بين المصابين بالفصام والمجموعة المرجعية. [20]
ملخص
أظهر المصابون بالفصام اختلافًا في التلفيف الحزامي الأمامي مقارنةً مع مجموعة مرجعية. وُجد أن التلفيف الحزامي الأمامي أصغر عند المصابين بالفصام. وُجد أن حجم المادة الرمادية في التلفيف الحزامي الأمامي أقل عند المصابين بالفصام. تمتلك الإناث الأصحاء تلفيفًا حزاميًا أماميًا منقاريًا أكبر من الذكور، وهذا الاختلاف الجنسي في الحجم غير موجود عند المصابين بالفصام. كان معدل استقلاب الغلوكوز أقل في التلفيف الحزامي الأمامي الأيسر وفي التلفيف الحزامي الخلفي الأيمن.
بالإضافة للتغيرات في القشرة الحزامية، تظهر المزيد من البنى الدماغية المزيد من التغيرات عند المصابين بالفصام مقارنةً مع مجموعة مرجعية. وُجد أن الحصين عند المصابين بالفصام أصغر حجمًا مقارنةً مع مجموعة مرجعية من نفس العمر، وبشكل مشابه، وُجد أن المذنب والبَطامة أصغر حجمًا في دراسة طولانية عند المصابين بالفصام. وبينما يكون حجم المادة الرمادية أصغر، كان حجم البطين الجانبي والبطين الثالث أكبر عند المصابين بالفصام.[21][22][23]
التاريخ
تعني (Cingulum) حزامًا في اللاتينية. اختير هذا الاسم غالبًا بسبب إحاطة جزء كبير من هذه القشرة بالجسم الثفني. تُعتبر القشرة الحزامية جزءًا من «الفص الحوفي الكبير» لبروكا (1878) الذي يتألف من قسم حزامي علوي (فوق ثفني) وقسم حصيني سفلي (تحت ثفني). ينفصل الفص الحوفي عن المتبقي من القشرة عبر بروما لسببين: أولًا لأنه ليس ملففًا، وثانيًا لأن التلافيف موجهة بشكل مجاور سهميًا (بعكس التلفف المستعرض). يُلاحظ التلفف (تشكل التلافيف) مجاور السهمي عند الأنواع غير الرئيسيات. وبذلك يصنف الفص الحوفي على أنه «حيواني». وكما في الأجزاء الأخرى من القشرة، كانت وما زالت هناك اختلافات حول الحدود والتسمية. ميّز برودمان (1909) 24 باحة (حزامية أمامية) و23 باحة (خلفية) بناءً على تحببها. مؤخرًا، ضُمت جميعها إلى جزء من الفص الحوفي في المصطلحات التشريحية (1998) بعد نظام فون إيكونومو (1925).[24][25][26]
صور إضافية

السطح الإنسي لنصف الكرة المخية. مشهد إنسي. تشريح عميق.

السطح الإنسي لنصف الكرة المخية. مشهد إنسي. تشريح عميق.
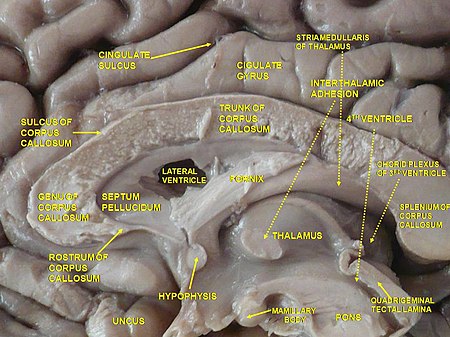
السطح الإنسي لنصف الكرة المخية. مشهد إنسي. تشريح عميق.
المراجع
- معرف النموذج التأسيسي في التشريح: 62434 — تاريخ الاطلاع: 1 أغسطس 2019
- Hadland, K. A.; Rushworth M.F.; et al. (2003). "The effect of cingulate lesions on social behaviour and emotion". Neuropsychologia. 41 (8): 919–931. doi:10.1016/s0028-3932(02)00325-1. PMID 12667528.
- "Cingulate binds learning". Trends Cogn Sci. 1 (1): 2. 1997. doi:10.1016/s1364-6613(97)85002-4. PMID 21223838.
- Kozlovskiy, S.; Vartanov A.; Pyasik M.; Nikonova E.; Velichkovsky B. (10 October 2013). "Anatomical Characteristics of Cingulate Cortex and Neuropsychological Memory Tests Performance". Procedia - Social and Behavioral Sciences. 86: 128–133. doi:10.1016/j.sbspro.2013.08.537.
- Kozlovskiy, S.A.; Vartanov A.V.; Nikonova E.Y.; Pyasik M.M.; Velichkovsky B.M. (2012). "The Cingulate Cortex and Human Memory Processes". Psychology in Russia: State of the Art. 5: 231–243. doi:10.11621/pir.2012.0014.
- Hayden, B. Y.; Platt, M. L. (2010). "Neurons in Anterior Cingulate Cortex Multiplex Information about Reward and Action". Journal of Neuroscience. 30 (9): 3339–3346. doi:10.1523/JNEUROSCI.4874-09.2010. PMC . PMID 20203193.
- Drevets, W. C.; Savitz, J.; Trimble, M. (2008). "The subgenual anterior cingulate cortex in mood disorders". CNS spectrums. 13 (8): 663–681. doi:10.1017/s1092852900013754. PMC . PMID 18704022.
- Adams, R.; David, A. S. (2007). "Patterns of anterior cingulate activation in schizophrenia: A selective review". Neuropsychiatric Disease and Treatment. 3 (1): 87–101. doi:10.2147/nedt.2007.3.1.87. PMC . PMID 19300540.
- Dorland's. Illustrated medical dictionary. Elsevier Saunders. صفحة 363. .
- McFarland, N. R.; Harber, S. N. (2000). "Convergent Inputs from Thalamic Motor Nuclei and Frontal Cortical Areas to the Dorsal Striatum in the Primate". The Journal of Neuroscience. 20 (10): 3798–3813.
- Wyass, J. M.; Van Groen, T. (1992). "Connections between the retrosplenial cortex and the hippocampal formation in the rat: A review". Hippocampus. 2 (1): 1–11. doi:10.1002/hipo.450020102. PMID 1308170.
- Pandya, D. N.; Hoesen, G. W.; Mesulam, M. -M. (1981). "Efferent connections of the cingulate gyrus in the rhesus monkey". Experimental Brain Research. 42–42 (3–4): 319–330. doi:10.1007/BF00237497.
- Fujii, M. (1983). "Fiber connections between the thalamic posterior lateral nucleus and the cingulate gyrus in the cat". Neuroscience Letters. 39 (2): 137–142. doi:10.1016/0304-3940(83)90066-6. PMID 6688863.
- Petrides, M; Pandya, DN (Oct 24, 2007). "Efferent association pathways from the rostral prefrontal cortex in the macaque monkey". The Journal of Neuroscience. 27 (43): 11573–86. doi:10.1523/JNEUROSCI.2419-07.2007. PMID 17959800.
- Cavada, C.; Goldman-Rakic, P. S. (1989). "Posterior parietal cortex in rhesus monkey: I. Parcellation of areas based on distinctive limbic and sensory corticocortical connections". The Journal of Comparative Neurology. 287 (4): 393–421. doi:10.1002/cne.902870402. PMID 2477405.
- Adey, W. R. (1951). "An Experimental Study of the Hippocampal Connexions of the Cingulate Cortex in the Rabbit". Brain. 74 (2): 233–247. doi:10.1093/brain/74.2.233. PMID 14858747.
- Takahashi, T.; Suzuki, M.; Kawasaki, Y.; Hagino, H.; Yamashita, I.; Nohara, S.; Nakamura, K.; Seto, H.; Kurachi, M. (2003). "Perigenual cingulate gyrus volume in patients with schizophrenia: A magnetic resonance imaging study". Biological Psychiatry. 53 (7): 593–600. doi:10.1016/S0006-3223(02)01483-X. PMID 12679237.
- Haznedar, M. M.; Buchsbaum, M. S.; Hazlett, E. A.; Shihabuddin, L.; New, A.; Siever, L. J. (2004). "Cingulate gyrus volume and metabolism in the schizophrenia spectrum". Schizophrenia Research. 71 (2–3): 249–262. doi:10.1016/j.schres.2004.02.025. PMID 15474896.
- Costain, G.; Ho, A.; Crawley, A. P.; Mikulis, D. J.; Brzustowicz, L. M.; Chow, E. W. C.; Bassett, A. S. (2010). "Reduced gray matter in the anterior cingulate gyrus in familial schizophrenia: A preliminary report". Schizophrenia Research. 122 (1–3): 81–84. doi:10.1016/j.schres.2010.06.014. PMC . PMID 20638248.
- Fujiwara, H.; Hirao, K.; Namiki, C.; Yamada, M.; Shimizu, M.; Fukuyama, H.; Hayashi, T.; Murai, T. (2007). "Anterior cingulate pathology and social cognition in schizophrenia: A study of gray matter, white matter and sulcal morphometry" ( كتاب إلكتروني PDF ). NeuroImage. 36 (4): 1236–1245. doi:10.1016/j.neuroimage.2007.03.068. PMID 17524666. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 24 سبتمبر 2019.
- Koolschijn, P. C. D. M. P.; Van Haren, N. E. M.; Cahn, W.; Schnack, H. G.; Janssen, J.; Klumpers, F.; Hulshoff Pol, H. E.; Kahn, R. S. (2010). "Hippocampal Volume Change in Schizophrenia". The Journal of Clinical Psychiatry. 71 (6): 737–744. doi:10.4088/JCP.08m04574yel. PMID 20492835.
- Mitelman, S. A.; Canfield, E. L.; Chu, K. W.; Brickman, A. M.; Shihabuddin, L.; Hazlett, E. A.; Buchsbaum, M. S. (2009). "Poor outcome in chronic schizophrenia is associated with progressive loss of volume of the putamen". Schizophrenia Research. 113 (2–3): 241–245. doi:10.1016/j.schres.2009.06.022. PMC . PMID 19616411.
- Kempton, M. J.; Stahl, D.; Williams, S. C. R.; Delisi, L. E. (2010). "Progressive lateral ventricular enlargement in schizophrenia: A meta-analysis of longitudinal MRI studies". Schizophrenia Research. 120 (1–3): 54–62. doi:10.1016/j.schres.2010.03.036. PMID 20537866.
- Broca, P (1878). "Anatomie comparee des circonvolutions cerebrales: Le grand lobe limbique et la scissure limbique dans la serie des mammifères". Revue d'Anthropologie. 1: 385–498
- "Terminologica Anatomica". www.unifr.ch (باللغة الإنجليزية). مؤرشف من الأصل في 16 مايو 201915 نوفمبر 2018.
- Economo, C., Koskinas, G.N. (1925). Die Cytoarchitektonik der Hirnrinde des erwachsenen Menschen. Wien: Springer Verlag.