|
هومو | |
|---|---|
 | |
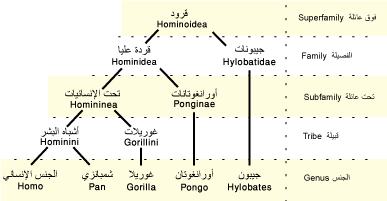
| المرتبة التصنيفية | جنس[1][2] |
| التصنيف العلمي | |
| فوق النطاق | حيويات |
| مملكة عليا | حقيقيات النوى |
| مملكة | حيوان |
| عويلم | ثنائيات التناظر |
| مملكة فرعية | ثانويات الفم |
| شعبة | حبليات |
| شعيبة | فقاريات |
| شعبة فرعية | فكيات |
| عمارة | رباعيات الأطراف |
| طائفة | ثدييات |
| طويئفة | وحشيات |
| صُنيف فرعي | مشيميات |
| رتبة ضخمة | وحشيات شمالية |
| رتبة كبرى | فوق رئيسيات |
| رتبة كبرى | أسلاف حقيقية |
| رتبة متوسطة | أشباه رئيسيات |
| رتبة | رئيسيات |
| رتيبة | نسناسيات بسيطة الأنف |
| تحت رتبة | سعالي |
| رتبة صغرى | نسناسيات نازلة الأنف |
| فصيلة عليا | قرد |
| فصيلة | قردة عليا |
| فُصيلة | أسرة الإنسانيات |
| الاسم العلمي | |
| Homo[1][2] كارولوس لينيوس ، 1758 |
|
الهومو، (Homo) هو جنس يندرج تحت الفصيلة الأَناسِيةّ (القردة العليا) في علم التصنيف الحيوي، ويتفرع منه الإنسان الحديث أوالأنسان العاقل والأنواع الحيوية المرتبطة به. عامة يعرف الجنس في علم الأحياء من حيث سلفه الموحد وسَيْغ تَرَاصّه وتباينه عن غيره من الأجناس وجنس الهومو يعني الجنس البشري لأن كلمة هومو تعني بشر باللغة اللاتينية. لم يتبق من هذا الجنس سوى نوع الإنسان المعاصر المعروف علميًا باسم الإنسان العاقل الحديث أما باقي أنواع هذا الجنس فقد انقرضت. كان البشر السالفون للإنسان العاقل أدمغة صغيرة في مطلع ظهور هذا الجنس، الذي من المحتمل يكون قد تطور من أسلاف عمارة الأسترالوبيثكوس (australopithecine) المندرجة تحت قبيلة أشباه البشر، بحوالي 2.3 إلي 2.4 مليون سنة [4][5] مع بداية ظهور الإنسان الماهر. طُرِحَت العديد من الأنواع الحيوية كجد أعلى مباشر لنسل الهومو (النسل الإنساني)، بما في ذلك الأسترالوبيثكس جارهي، والأسترالوبيثكس سيديبا، والأسترالوبيثكس الإفريقي، وأسترالوبيثكس أفارينيسيس.[6][7] هذه الأنواع الحيوية تمتلك سمات مورفولوجية (تَشَكُّلية أو تكوينية) تتوائم مع سمات جنس الهومو (الإنساني)، لكن ليس هناك توافق في الآراء بشأن أيهم أدى إلى ظهور جنس الهومو، وبافتراض أن ذلك السلف (أو الجد الأعلي) ليس نوع آخر غير مكتشف حتى الآن.
أبرز التطورات الفسيولوجية (الوظيفية) بين جنس الهومو والأنواع قُرْدُوحِيّة الجنوبية السابقة هي الزيادة في سعة القِحْف (الجمجمة)، من حوالي 450 سم3 (أو 27 بوصة مكعبة) في أ. جارهي إلى 600 سم3 (أو 37 بوصة مكعبة) في الإنسان الماهر. وبمرور مئات آلاف السنين تضاعفت سعة الجمجمة مرات أخرى من نحو 700 سم3 (الدماغ) حتى أصبحت في الإنسان العاقل نحو 1300 سم 3 ضمن الجنس الهومو (البشري) ، ويقترن بذلك تقدم القدرة على الكلام، بتشكل الحنجرة، من الإنسان الماهر وخلال الإنسان العامل أو الإنسان المنتصب وحتي إنسان هايدلبيرغ من حوالي 0.6 مليون سنة مضت. وتتداخل سعة الجمجمة لإنسان هايدلبيرغ مع النطاق المألوف في الإنسان الحديث.
هناك عدة سمات لجنس الهومو لا توجد في باقي الرئيسيات، وبعض هذه الصفات متواجدة لدى بعض الرئيسيات ولكن بصورة أقل بكثير مثل:
- حجم جمجمة كبير ويشمل حيزا كبيرا للمخ إذ يعتقد أنه نما بسبب ضعف وضيق عضلة اللثة
- المشي على اثنين وهي صفة يشترك في الإنسان مع الإنسان الحديث
- قدرة استيعاب واسعة وفهم للروابط والعلاقات بين الأشياء والتأثيرات المميزة للحركات والتغيرات المختلفة
- بناء أدوات معقدة واستخدام الادوات لصنع أدوات
- قدرة على الاتصال ولغة معقدة وميل طبيعي لتطوير لغة مشتركة مع بني جنسه وإن كانوا لا يشتركون في نفس اللغة.
تم حتى الآن اكتشاف عدد من الأنواع وإدراجها في تصنيف البشر كجنس وهي:
الإنسان الماهر، إنسان بحيرة رودولف، إنسان جيورجيا، الإنسان العامل، الإنسان المنتصب إنسان سيبرانو، الإنسان السالف، إنسان هايدلبيرغ، إنسان نياندرتال، هومو ناليدي، إنسان روديسيا، إنسان فلوريس، الإنسان العاقل القديم، الإنسان الحديث تشريحيا: (الإنسان العاقل الأول، الإنسان العاقل العاقل).
التسمية
في مجال العلوم البيولوجية، وخاصة الإنثروبولوجي، (علم الإنسان) وعلم الحفريات، الاسم الشائع لجميع أعضاء جنس الهومو هو بشر و"إنسان"، [8] وكلمة هومو هي كلمة لاتينية تعني "إنسان"، وجاءت لتعني "رجل" في اللاتينية الحديثة. وقد صيغ اسم الإنسان العاقل من قِبل كارولوس لينيوس [9] (1758).[10]
الأسماء والتصنيف
يُقصد بلفظ هومو (homō) اللاتيني «كائن بشري» أو «إنسان» بالمعنى العام لمصطلح «البشرية».[9] وقد تمت صياغة الاسم الثنائي «الإنسان العاقل» (Homo sapiens) على يد كارل لينيوس في عام 1758.[11] وظهرت أسماء الأنواع الأخرى لهذا الجنس بداية من النصف الثاني من القرن العشرين (إنسان نياندرتال (1864)، الإنسان المنتصب (1892))
حتى يومنا هذا، لم يصدر تعريف لجنس الإنسان (هومو) بشكل لائق.[12][13][14] ولأن السجل الأحفوري الإنساني كان بطيئا في الخروج من الأرض، فإن الحدود الفاصلة وتعريف جنس الهومو جرى تعريفهما بشكل سيء ويتعرضان للتغيير باستمرار. ولأنه لم يكن هناك سبب للاعتقاد في أن لهذا الجنس أفراد آخرون، فإن كارل لينيوس لم يكلف نفسه تعريف الهومو حينما صاغها لأول مرة للبشر في القرن الثامن عشر. وقد أتى اكتشاف النياندرتال بأول إضافة جديدة.
اتخذ جنس الهومو اسمه التصنيفي للإشارة إلى أن أفراد نوعه يمكن أن يصنفوا كبشر. وبمرور عقود القرن العشرين، قدمت الاكتشافات الأحفورية للأنواع قبل البشرية والبشرية البدائية، التي تعود إلى الفترة ما بين أواخر العصر الميوسيني ومطلع العصر البيلوسيني، تركيبة غنية للتصنيفات المتناظرة. وهناك جدال مستمر حول تمييز الهومو عن الأسترالوبيثكس –أو في الواقع تمييز الهومو عن البعام، حيث يجادل شطر من العلماء بأن هذين النوعين من الشمبانزي ينبغي تصنيفهما مع جنس البشر بدلا من البعام. لكن حتى لو تم ذلك، فإن تصنيف أحافير الهومو يتوفق مع الأدلة التالية: 1) ثنائية الحركة الموجودة لدى الإنسان الماهر، والموروثة عن الأسترالوبيثكس البدائي منذ 4 ملايين عام، كما تثبت آثار أقدام لايتولي؛ و2) ثقافة الأدوات البشرية التي كانت قد بدأت منذ 2.5 مليون عام.
منذ أواخر القرن التاسع عشر وحتى منتصف القرن العشرين، قدم عدد من الأسماء التصنيفية الجديدة التي تشتمل على أسماء عامة تم اقتراحها من أجل الأحافير الإنسانية الأولى؛ وقد اختلط معظمها منذ ذلك الوقت مع مصطلح الهومو كاعتراف بأن الإنسان المنتصب (الهومو إريكتوس) نوع مميز ومتفرد وواسع الانتشار جغرافيا بسبب الهجرات الأولى. وقد أطلقت العديد من الأسماء بوصفها «مرادفات تصنيفية» لمصطلح هومو، ومنها البثكانثروبس،[15] البروتانثروبس،[16] السينانثروبس،[17] السيفانثروبس،[18] الأفريكانثروبس،[19] التلانثروبس،[20] الأتلانثروبس،[21] والتشادانثروبس.[22]
لا يزال تصنيف جنس الهومو إلى أنواع وأنواع فرعية يعاني من معلومات غير كاملة ولا يزال مستواه متدنيا. وقد أدى ذلك إلى استخدام أسماء شائعة («النياندرتال» و«دينيسوفان») حتى في الأوراق البحثية، وذلك لتجنب الأسماء الثلاثية أو الالتباس في تصنيف المجموعات ضمن أصنوفة غير محددة –ومن أمثلة ذلك: هومو نياندرتال مقابل هومو سابينز نياندرتال، أو هومو جورجيكوس مقابل هومو إريكتوس جورجيكوس.[23] هناك بعض الأنواع المنقرضة المكتشفة حديثا ضمن جنس الهومو ولم يُجمع حتى الآن على اسم ثنائي لها (مثل إنسان دينيسوفان وجماعة كهف ريد دير).[24] ومنذ بداية العصر الهولوسيني، فمن المحتمل أن الإنسان العاقل (الهومو سابينز)، أي البشر الحاليين من الناحية التشريحية، كان النوع الوحيد المتواجد من الهومو.
كان جون إدوارد غراي (1825) من الداعمين الأوائل لتحديد الأصانيف بتخصيص قبائل وعائلات.[25] واقترح وود وريتشموند (2000) أن يتم تصنيف أشباه البشر كقبيلة شملت كل أنواع الإنسان وأسلاف البشر منذ الفترة التي تلي آخر سلف مشترك للبشر والشمبانزي؛ وأن تخصص الهومينينا كقبيلة فرعية من أشباه البشر (الهومينيني) بحيث تشمل جنس الهومو فحسب –وذلك لا يشمل أشباه البشر المنتصبين الأوائل بالعصر البليوسيني.[26]
تطوره
الأسترالوبيثكس
قدمت اقتراحات بأنواع عديدة، منها الأسترالوبيثكس غاري، الأسترالوبيثكس سيديبا، الأسترالوبيثكس أفريكانوس، والأسترالوبيثكس أفارسينسس، على أنها أسلاف مباشرة لنسل الهومو.[27][28] ولهذه الأنواع خصائص مورفولوجية متشابهة مع الهومو، إلا أنه لم يتم الإجماع على النوع الذي انحدر منه الهومو من بينها.
ومنذ عام 2010 على وجه الخصوص، أصبح تمييز الهومو عن الأسترالوبيثكس أكثر إثارة للجدل. فقد كان السائد أن ظهور الهومو تزامن مع أول استخدام للأدوات الحجرية (الصناعة الأولدوانية)، أي مع بداية العصر الحجري القديم السفلي. لكن في عام 2010، ظهر ما يبدو أنه دليل على أن استخدام الأدوات الحجرية كان منذ وجود الأسترالوبيثكس أفارينسس قبل حوالي 3.3 مليون عام مضت، أي ما يقرب من مليون عام قبل أول ظهور للهومو.[29] كما أن حفرية إل دي 350-1 ال، وهي جزء من الفك السفلي الأحفوري الذي يعود تاريخ إلى 2.8 مليون عام، التي اكتشفت في عام 2015 بولاية عفر بأثيوبيا، وصفت بأنها «تجمع الصفات البدائية الموجودة في الأسترالوبيثكس ومورفولوجيا مستمدة من الهومو اللاحق».[30] ويرجع بعض المؤلفين تطور الهومو إلى ما يقرب أو ما يزيد عن 3 مليون عام.[35] ويشك البعض فيما إذا كان ينبغي تصنيف الهومو هابيليس ضمن جنس الهومو، مقترحين بدلا من ذلك أن أصل الهومو والهومو إريكتوس يعود إلى 1.9 مليون عام تقريبا.[31]
تعتبر الزيادة في حجم باطن قحف الجمجمة التطور الأبرز في وظائف الأعضاء بين نوع الأسترالوبيثكس المبكر والهومو، فقد زادت من 460 سم مكعب لدى الأسترالوبيثكس غاري إلى 660 سم مكعب لدى الهومو هابيليس، وزادت لدى الهومو إريكتوس لتصبح 760 سم مكعب، وإلى 1250 سم مكعب لدى الهوموو هايديلبيرحنسيس، وإلى 1760 سم مكعب لدى الهومو نياندرتال. ومع ذلك، فإن الزيادة الثابتة في سعة الجمجمة موجودة بالفعل لدى الأسترالوبيثكينا ولا ينقطع بعد ظهور الهومو، ولذلك فإنه لا يعتبر معيارا موضوعيا في تفسير ظهور هذا الجنس.[32]
الإنسان الماهر
ظهر الإنسان الماهر قبل حاولي 2.1 مليون عام. قبل عام 2010، كانت هناك اقتراحات بألا يدرج الهومو هابيليس ضمن جنس الهومو، وأن يدرج بدلا من ذلك ضمن الأسترالوبيثكس.[33][34] والسبب الرئيسي لإدراج الهومو هابيليس ضمن جنس الهومو هو أن استخدامه للأدوات عفى عليه الزمن حينما اكتشف أن استخدام الأسترالوبيثكس للأدوات سبق استخدام الهومو هابيليس بمليون عام على الأقل.[29] وعلاوة على ذلك، فالاعتقاد الذي ساد طويلا هو أن الهومو هابيليس كان سلفا للهومو إريكتوس. وفي عام 2007، تم اكتشاف أن الهومو هابيليس والهومو إريكتوس تواجدا معا لمدة من الزمن، الأمر الذي يشير إلى أن الهومو إريكتوس لم يأتي مباشرة من نسل الهومو هابيليس، بل من سلف مشترك لهما.[35] بعد اكتشاف جمجمة دمانيسي 5 في عام 2013، أصبح هناك شك في أن الهومو إريكتوس الآسيوي متحدر من الهومو إرغاستر الأفريقي الذي يتحدر بدوره من الهومو هابيليس. لكن عوضا عن ذلك، فإن الظاهر أن الهومو إرغاستر والهومو إريكتوس أشكال متنوعة لنفس النوع، وهذا النوع قد يكون نشأ في أفريقيا أو في آسيا[36] وانتشر في نطاق عريض عبر أوراسيا (بما في ذلك أوروبا، إندونيسيا، الصين) منذ نصف مليون عام.[37]
الإنسان المنتصب
عادة ما يفترض أن الإنسان المنتصب تطور بشكل تجددي من الهومو هابيليس قبل مليوني عام مضت. وقد عزز هذا السيناريو باكتشاف الهومو إريكتوس جورجيكس، وحي حفريات لأفراد بدائيين من الهومو إريكتوس وجدت في القوقاز، وبدت عليها صفات انتقالية تتشابه مع الهومو هابيليس. ولدى اكتشاف أولى الأدلة على الهومو إريكتوس خارج أفريقي، أصبح معقولا أن يكون الهومو إريكتوس قد تطور في أوراسيا ثم هاجر إلى رجوعا إلى أفريقيا. وبناء على أحافير وجدت في كوبي فورا، شرق بحيرة توركانا بكينيا، جادل سبور وآخرون (2007) بأن الهومو هابيليس ربما يكون قد نجا للفترة التي تلي ظهور الهومو إريكتوس، ومن ثم فإن تطور الهومو إريكتوس لم يكن تجدديا، وتواجد الهومو إريكتوس جنبا إلى جنب مع الهومو هابيليس لنحو نصف مليون عام (في الفترة ما بين 1.9 إلى 1.4 مليون عام مضت)، وذلك أثناء العصر الكالاباري.[38]
وقد عثر في جنوب أفريقيا على نوع منفصل، الهومو غوتن، وافتُرض في عام 2010 أنه عاصر الهومو إريكتوس.
مراجع
- وصلة : التصنيف التسلسلي ضمن نظام المعلومات التصنيفية المتكامل — تاريخ الاطلاع: 19 سبتمبر 2013 — العنوان : Integrated Taxonomic Information System — تاريخ النشر: 1999
- وصلة : http://www.departments.bucknell.edu/biology/resources/msw3/browse.asp?s=y&id=12100794 — تاريخ الاطلاع: 18 سبتمبر 2015 — العنوان : Mammal Species of the World
- وصلة : http://www.departments.bucknell.edu/biology/resources/msw3/browse.asp?s=y&id=12100794 — المؤلف: فرانسيسكو أيالا — العنوان : Genera of the human lineage — المجلد: 100 — الصفحة: 7688 — العدد: 13 — نشر في: Proceedings of the National Academy of Sciences of the United States of America — https://dx.doi.org/10.1073/PNAS.0832372100 — https://pubmed.ncbi.nlm.nih.gov/12794185 — https://www.ncbi.nlm.nih.gov/pmc/articles/PMC164648
- Stringer, C.B. (1994). "Evolution of early humans". In Steve Jones, Robert Martin & David Pilbeam (eds.) (المحررون). The Cambridge Encyclopedia of Human Evolution. Cambridge: Cambridge University Press. صفحة 242. . Also (paperback)
- McHenry, H.M (2009). "Human Evolution". In Michael Ruse & Joseph Travis (المحررون). Evolution: The First Four Billion Years. Cambridge, Massachusetts: The Belknap Press of Harvard University Press. صفحة 265. .
- Pickering, R., Dirks, P. H., Jinnah, Z., De Ruiter, D. J., Churchill, S. E., Herries, A. I., ... & Berger, L. R. (2011). Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo" Science 333(6048), 1421-1423.
- Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S., & Suwa, G. (1999). Australopithecus garhi: a new species of early hominid from Ethiopia" Science 284(5414), 629-635.
- OpenStax College (30 May 2013). Biology. Houston, TX: Rice University. صفحة 13. . مؤرشف من الأصل في 30 مايو 2014. (Free PDF download available on web page)
- Note: In 1959, Linnaeus was designated as the عينة نمطية for إنسان (Stearn, W. T. 1959. "The background of Linnaeus's contributions to the nomenclature and methods of systematic biology", Systematic Zoology 8 (1): 4-22, p. 4) which means that following the القانون الدولي للتسمية الحيوانية, Homo sapiens was validly defined as the animal species to which Linnaeus belonged.
- Linné, Carl von (1758). Systema naturæ. Regnum animale (الطبعة 10). صفحات 18, 20. مؤرشف من الأصل في 19 أكتوبر 201919 نوفمبر 2012.
- The word "human" itself is from Latin humanus, an adjective formed on the root of homo, thought to derive from a لغة هندية أوروبية بدائية word for "earth" reconstructed as . dhghem The American Heritage Dictionary of the English Language: Fourth Edition. 2000. نسخة محفوظة 10 فبراير 2009 على موقع واي باك مشين.
- Schwartz, Jeffrey H.; Tattersall, Ian (28 August 2015). "Defining the genus Homo". Science. 349 (6251): 931–932. Bibcode:2015Sci...349..931S. doi:10.1126/science.aac6182. PMID 26315422. مؤرشف من الأصل في 26 أكتوبر 201502 نوفمبر 2015.
- Lents, Nathan (4 October 2014). "Homo naledi and the Problems with the Homo Genus". The Wildernist. مؤرشف من الأصل في 18 نوفمبر 201502 نوفمبر 2015.
- Wood, B.; Collard, M. (2 April 1999). "The human genus". Science. 284 (5411): 65–71. doi:10.1126/science.284.5411.65. PMID 10102822.
- "ape-man", from Pithecanthropus erectus (إنسان جاوة), Eugène Dubois, Pithecanthropus erectus : eine menschenähnliche Übergangsform aus Java (1894), identified with the Pithecanthropus alalus (i.e. "non-speaking ape-man") hypothesized earlier by إرنست هيكل
- "early man", Protanthropus primigenius إرنست هيكل, Systematische Phylogenie vol. 3 (1895), p. 625 - تصفح: نسخة محفوظة 06 نوفمبر 2018 على موقع واي باك مشين.
- "Sinic man", from Sinanthropus pekinensis (إنسان بكين), ديفيدسون بلاك (1927).
- "crooked man", from Cyphanthropus rhodesiensis (إنسان روديسيا) William Plane Pycraft (1928).
- "African man", used by T.F. Dreyer (1935) for the جمجمة فلورسباد he found in 1932 (also Homo florisbadensis or Homo helmei). Also the genus suggested for a number of archaic human skulls found at Lake Eyasi by Weinert (1938). Leaky, Journal of the East Africa Natural History Society' (1942), p. 43. نسخة محفوظة 06 نوفمبر 2018 على موقع واي باك مشين.
- "remote man"; from Telanthropus capensis (Broom and Robinson 1949), see (1961), p. 487. نسخة محفوظة 06 نوفمبر 2018 على موقع واي باك مشين.
- from Atlanthropus mauritanicus, name given to the species of fossils (three lower jaw bones and a parietal bone of a skull) discovered in 1954 to 1955 by كميل أرامبورغ in تغنيف, Algeria. Arambourg, C. (1955). "A recent discovery in human paleontology: Atlanthropus of ternifine (Algeria)". American Journal of Physical Anthropology. 13 (2): 191–201. doi:10.1002/ajpa.1330130203.
- Y. Coppens, "L'Hominien du Tchad", Actes V Congr. PPEC I (1965), 329f.; "Le Tchadanthropus", Anthropologia 70 (1966), 5–16.
- Alexandra Vivelo (August 25, 2013), Characterization of Unique Features of the Denisovan Exome - تصفح: نسخة محفوظة 2013-10-29 على موقع واي باك مشين.
- Barras, Colin (2012-03-14). "Chinese human fossils unlike any known species". New Scientist. مؤرشف من الأصل في 2 يوليو 201515 مارس 2012.
- J.E. Gray, "An outline of an attempt at the disposition of Mammalia into Tribes and Families, with a list of genera apparently appertaining to each Tribe", Annals of Philosophy', new series (1825), pp. 337–344.
- Brunet, M.; et al. (2002). "A new hominid from the upper Miocene of Chad, central Africa". Nature. 418 (6894): 145–151. doi:10.1038/nature00879. PMID 12110880.
- Pickering, R.; Dirks, P.H.; Jinnah, Z.; De Ruiter, D.J.; Churchill, S.E.; Herries, A.I.; Berger, L.R. (2011). "Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo". Science. 333 (6048): 1421–1423. Bibcode:2011Sci...333.1421P. doi:10.1126/science.1203697. PMID 21903808.
- Asfaw, B.; White, T.; Lovejoy, O.; Latimer, B.; Simpson, S.; Suwa, G. (1999). "Australopithecus garhi: a new species of early hominid from Ethiopia". Science. 284 (5414): 629–635. Bibcode:1999Sci...284..629A. doi:10.1126/science.284.5414.629. PMID 10213683.
- McPherron, S.P.; Alemseged, Z.; Marean, C.W.; Wynn, J.G.; Reed, D.; Geraads, D.; Bobe, R.; Bearat, H.A. (2010). "Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia". Nature. 466 (7308): 857–860. Bibcode:2010Natur.466..857M. doi:10.1038/nature09248. PMID 20703305. "The oldest direct evidence of stone tool manufacture comes from Gona (Ethiopia) and dates to between 2.6 and 2.5 million years (Myr) ago. [...] Here we report stone-tool-inflicted marks on bones found during recent survey work in Dikika, Ethiopia [... showing] unambiguous stone-tool cut marks for flesh removal [..., dated] to between 3.42 and 3.24 Myr ago [...] Our discovery extends by approximately 800,000 years the antiquity of stone tools and of stone-tool-assisted consumption of ungulates by hominins; furthermore, this behaviour can now be attributed to Australopithecus afarensis."
- Villmoare, Brian; Kimbel, William H.; Seyoum, Chalachew; Campisano, Christopher J.; DiMaggio, Erin N.; Rowan, John; Braun, David R.; Arrowsmith, J. Ramón; Reed, Kaye E. (2015-03-20). "Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia". Science. 347 (6228): 1352–1355. Bibcode:2015Sci...347.1352V. doi:10.1126/science.aaa1343. ISSN 0036-8075. PMID 25739410. مؤرشف من الأصل في 30 ديسمبر 2015. . See also: Erin N. DiMaggio; Campisano C.J.; Rowan J.; Dupont-Nivet G.; Deino A.L.; et al. (2015). "Late Pliocene fossiliferous sedimentary record and the environmental context of early Homo from Afar, Ethiopia". Science. 347 (6228): 1355–1359. Bibcode:2015Sci...347.1355D. doi:10.1126/science.aaa1415. PMID 25739409. مؤرشف من الأصل في 07 مارس 2015.
- Cela-Conde and Ayala (2003) recognize five genera within Hominina: Ardipithecus, Australopithecus (including Paranthropus), Homo (including Kenyanthropus), Praeanthropus (including Orrorin), and Sahelanthropus. Cela-Conde, C.J.; Ayala, F.J. (2003). "Genera of the human lineage". Proceedings of the National Academy of Sciences. 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. doi:10.1073/pnas.0832372100. PMC . PMID 12794185.
- "the adaptive coherence of Homo would be compromised if H. habilis is included in Homo. Thus, if these arguments are accepted the origins of the genus Homo are coincident in time and place with the emergence of H. erectus, not H. habilis" Bernard Wood, "Did early Homo migrate 'out of' or 'in to' Africa?", PNAS vol. 108, no.26 (28 June 2011), 10375–10376. نسخة محفوظة 18 أبريل 2018 على موقع واي باك مشين.
- Wood and Richmond; Richmond, BG (2000). "Human evolution: taxonomy and paleobiology". Journal of Anatomy. 197 (Pt 1): 19–60. doi:10.1046/j.1469-7580.2000.19710019.x. PMC . PMID 10999270. p. 41: "A recent reassessment of cladistic and functional evidence concluded that there are few, if any, grounds for retaining H. habilis in Homo, and recommended that the material be transferred (or, for some, returned) to Australopithecus (Wood & Collard, 1999)."
- Miller J.M.A. (2000). "Craniofacial variation in Homo habilis: an analysis of the evidence for multiple species". American Journal of Physical Anthropology. 112 (1): 103–128. doi:10.1002/(SICI)1096-8644(200005)112:1<103::AID-AJPA10>3.0.CO;2-6. PMID 10766947.
- F. Spoor; M.G. Leakey; P.N. Gathogo; F.H. Brown; S.C. Antón; I. McDougall; C. Kiarie; F.K. Manthi; L.N. Leakey (2007-08-09). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya". Nature. 448 (7154): 688–691. Bibcode:2007Natur.448..688S. doi:10.1038/nature05986. PMID 17687323. F. Spoor; M.G. Leakey; P.N. Gathogo; F.H. Brown; S.C. Antón; I. McDougall; C. Kiarie; F.K. Manthi; L.N. Leakey (2007-08-09). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya". Nature. 448 (7154): 688–691. Bibcode:2007Natur.448..688S. doi:10.1038/nature05986. PMID 17687323. "A partial maxilla assigned to H. habilis reliably demonstrates that this species survived until later than previously recognized, making an anagenetic relationship with H. erectus unlikely"
- Prins, Harald E.L.; Walrath, Dana; McBride, Bunny (2007). Evolution and prehistory: the human challenge. Wadsworth Publishing. صفحة 162. . مؤرشف من الأصل في 26 يناير 2020. .
- Augusti, Jordi; Lordkipanidze, David (June 2011). "How "African" was the early human dispersal out of Africa?". Quaternary Science Reviews. 30 (11–12): 1338–1342. Bibcode:2011QSRv...30.1338A. doi:10.1016/j.quascirev.2010.04.012.
- "A partial maxilla assigned to H. habilis reliably demonstrates that this species survived until later than previously recognized, making an anagenetic relationship with H. erectus unlikely. The discovery of a particularly small calvaria of H. erectus indicates that this taxon overlapped in size with H. habilis, and may have shown marked sexual dimorphism. The new fossils confirm the distinctiveness of H. habilis and H. erectus, independently of overall cranial size, and suggest that these two early taxa were living broadly sympatrically in the same lake basin for almost half a million years." Spoor, F; Leakey, M.G; Gathogo, P.N; Brown, F.H; Antón, S.C; McDougall, I; Kiarie, C; Manthi, F.K.; Leakey, L.N. (2007). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya". Nature. 448 (7154): 688–691. Bibcode:2007Natur.448..688S. doi:10.1038/nature05986. PMID 17687323.