
Un flagelle est un organite qui par ses mouvements permet la mobilité d'une cellule. Une même cellule peut en posséder plusieurs. Au plan fonctionnel, on peut considérer les flagelles comme des prolongements du cytoplasme. Ils sont capables de mouvements périodiques induisant le déplacement de la cellule dans un milieu liquide, ou du milieu liquide autour de la cellule.
On trouve des flagelles chez les procaryotes (archées et bactéries) comme chez les eucaryotes, mais fondés sur des principes génétiques, architecturaux et fonctionnels très différents.
- Chez les procaryotes, les flagelles sont des structures semi-rigides ancrées dans la membrane plasmique et dont l'embase est constituée par un moteur moléculaire qui assure sa rotation. Les flagelles des bactéries et des archées se ressemblent en apparence mais diffèrent profondément, si bien qu'on pense aujourd'hui qu'ils résultent d'une convergence évolutive plutôt que d'une homologie[1],[2].
- Chez les eucaryotes, les flagelles sont mus par un axonème, une structure du cytosquelette le long de laquelle les moteurs moléculaires sont périodiquement distribués.
Flagelles des bactéries
Chez les bactéries, l'expression de plus de 40 gènes est requise pour spécifier les protéines indispensables à l'assemblage séquentiel des flagelles, de la partie la plus interne vers la plus externe. La rotation du corps basal résulte d'une force proton-motrice produite par un moteur moléculaire homologue à l'ATPase H0H1 de la mitochondrie. Sur le plan cinématique, la rotation d'un tour du flagelle nécessite environ 1000 protons[3],[4]. La fréquence de rotation d'un flagelle bactérien est voisine de 40-60 Hz. Les flagelles bactériens sont relativement difficiles à observer en microscopie de fond clair, mais « facilement » observables en contraste de Nomarsky.
Il existe différents types de formation flagellaires ou ciliaires :
- un seul flagelle polaire = ciliature monotriche A (déplacement fléchant) ;
- plusieurs flagelles polaires = ciliature lophotriche B (déplacement fléchant + oscillant) ;
- un flagelle à chaque pôle = ciliature amphitriche C (déplacement oscillant) ;
- des cils/flagelles entourant la cellule = ciliature péritriche D (déplacement fléchant hélicoïdal).


Ultrastructure
Le flagelle « procaryote » est constitué de trois parties :
- le filament flagellaire : C'est la plus longue et la plus évidente, elle s'étend depuis la surface cellulaire : c'est cette partie qui est mise en évidence lors de colorations et observation en microscopie optique. C'est un cylindre creux et rigide constitué de nombreux monomères d'une seule protéine : la flagelline. Il existe cependant des flagelles complexes, composés de plusieurs protéines distinctes. Ces monomères semblent dans tous les cas emprunter le canal central pour être finalement assemblés à l'extrémité distale ;
- le crochet : Il est d'un diamètre supérieur au filament et est situé tout près de la surface cellulaire. Il fait la liaison entre le corps basal et le filament flagellaire. Son rôle est de transmettre le mouvement du corps basal au filament ;
- le corps basal : il est enfoui dans la cellule et sa structure diffère des bactéries Gram négatives et Gram positives. On peut le définir comme une petite tige centrale insérée dans un système d'anneaux, le tout enfoui au niveau de la membrane plasmique et la paroi.
- chez les Gram négatifs, on trouve quatre anneaux qui sont attachés à la tige centrale :
- deux anneaux extérieurs dont le rôle est de stabiliser et de fixer l'ensemble de la structure au corps cellulaire. L'anneau L est lié à la membrane externe et P est lié au peptidoglycane,
- deux anneaux internes M et S qui tournent, il s'agit en fait du moteur flagellaire, ce sont eux qui en tournant transmettent le mouvement au filament. M est lié à la membrane plasmique.
- chez les Gram négatifs, on trouve quatre anneaux qui sont attachés à la tige centrale :
Le flagelle est parfois recouvert d'une membrane, dérivée de la membrane externe dans certains cas, ou totalement originale dans d'autres cas.
- Chez les Gram positifs, on trouve seulement deux anneaux :
- un anneau externe lié au peptidoglycane ;
- un anneau interne, qui joue le rôle de moteur, il est lié à la membrane plasmique.
- Chez les Gram positifs, on trouve seulement deux anneaux :
Les anneaux internes (ou rotor) sont capables de tourner dans les deux sens, ce qui permet à la bactérie de changer de sens.
Les anneaux externes (ou stator) ne tournent pas, ils ont pour rôle de stabiliser l'ensemble de la structure flagellaire.
Flagelles des archées
Le flagelle des archées, appelé archaelle (en), est une structure unique dépourvue de canal central, qu'on trouve à la surface de nombreuses archées et qui leur confère une motilité natatoire. L'archaelle est constitué d'un filament hélicoïdal rigide attaché à la membrane cellulaire par un moteur moléculaire. Ce dernier, composé de protéines cytosoliques, membranaires et pseudo-périplasmiques, est responsable de l'assemblage du filament et, une fois assemblé, de sa rotation. La rotation du filament propulse les cellules archées en milieu liquide, à la manière d'une hélice de bateau.
L'archaelle est constitué d'archaellines, des protéines propres aux archées mais semblables aux pilines bactériennes de type IV, qui sont constituées de peptides signal de classe 3 et produites par une enzyme semblable aux prépiline peptidases de type IV. Les archaellines sont généralement modifiées par l'ajout de glycanes N-liés qui sont nécessaires pour un assemblage ou un fonctionnement correct[5].
Les travaux des années 1990 ont révélé de nombreuses différences entre les flagelles archéens et bactériens, notamment :
- la rotation des flagelles bactériens est alimentée par la force proton-motrice (un flux d'ions H+) ou occasionnellement par la force sodium-motrice (en) (un flux d'ions Na+). La rotation des flagelles archéens est alimentée par l'ATP[6] ;
- alors que les cellules bactériennes ont souvent de nombreux filaments flagellaires, dont chacun tourne indépendamment, le flagelle archéen est composé d'un faisceau de nombreux filaments qui tournent d'un seul bloc ;
- les flagelles bactériens se développent par l'ajout de sous-unités de flagelline à l'extrémité. Les flagelles archéens se développent par l'ajout de sous-unités à la base ;
- les flagelles bactériens sont plus épais que les archaelles, et le filament bactérien a un tube central creux suffisamment large pour que les sous-unités de flagelline puissent remonter à l'intérieur du filament et être ajoutées à la pointe. L'archaelle est trop mince pour cela (12–15 nm)[7] ;
- de nombreux composants des flagelles bactériens ont une similitude de séquence avec les composants des systèmes de sécrétion de type III, mais les composants des flagelles bactériens et des archaelles n'en ont aucune. Au lieu de cela, certains composants d'archaelles partagent une séquence et une similitude morphologique avec des composants de pili de type IV, qui sont assemblés par l'action de systèmes de sécrétion de type II (les nomenclature des systèmes de sécrétion de pili et de protéines ne sont pas cohérentes)[7].
Flagelles des eucaryotes
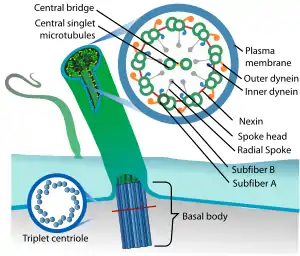
La structure des flagelles et des cils[alpha 1] des cellules eucaryotes est complètement différente de celles des procaryotes et nécessite l'expression de 250 à 400 gènes pour la seule mise en place de l'architecture axonémale dont le battement (< 70 Hz) est soit plan soit tridimensionnel. Un flagelle actif est une machine macromoléculaire permettant le mouvement, mais il est également aussi un couloir d'intense circulation de « trains » d'information, de substances énergétiques à acheminer et de déchets métaboliques à évacuer. Des techniques d'imagerie en microscopie électronique en 3D ont récemment montré qu'il contient sur toute sa longueur des doublets de microtubules permettant un transport intraflagellaire directionnel, jusqu'à la pointe ciliaire, tout en permettant les mouvements du cil. De premiers modèles mécaniste de la logistique du transport bidirectionnel intraflagellaire et de l'axonème (et de ses neuf doublets de microtubules) sont ainsi approchés[8].
Formation du flagelle lors de la spermiogenèse
Pour les spermatozoïdes humains, ils se forment à partir du centriole distal de la spermatide ronde. Ces deux centrioles migrent à la future partie postérieure de la cellule.
Le centriole le plus proche du noyau est dit proximal ou encore juxtanucléaire et le plus éloigné du noyau est dit distal.
Le flagelle du spermatozoïde des araignées
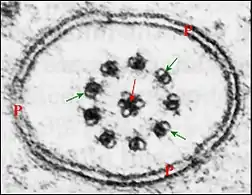
Chez les Araneae (Araignées), le flagelle du spermatozoïde se singularise par la présence de trois microtubules au lieu de deux dans sa partie centrale[9],[10],[11] dans des groupes variés : Pholcidae, Telemidae, Pisauridae
Notes et références
Notes
Références
- ↑ (en) « The archaeal flagellum: a different kind of prokaryotic motility structure », FEMS Microbiology Reviews, vol. 25, no 2, , p. 147–74 (DOI 10.1111/j.1574-6976.2001.tb00575.x).
- ↑ (en) « Archaeal flagellum », sur www.uniprot.org.
- ↑ (en) Frederick Carl Neidhardt et al., Escherichia coli and Salmonella : cellular and molecular biology, Washington, ASM Press, , 2822 p. (ISBN 1-55581-084-5 et 9781555810849, OCLC 34150023, lire en ligne), chap. 10 (« Flagella and motility »), p. 133-134.
- ↑ (en) David F. Blair, « Flagellar movement driven by proton translocation », FEBS Letters, vol. 545, no 1, , p. 86–95 (ISSN 1873-3468, DOI 10.1016/S0014-5793(03)00397-1, lire en ligne, consulté le ).
- ↑ (en) Pili and Flagella: Current Research and Future Trends, Caister Academic Press, (ISBN 978-1-904455-48-6).
- ↑ (en) Michael T. Madigan, Brock biology of microorganisms, NY, NY, , 70–71 p. (ISBN 9781292235103).
- 1 2 (en) « Assembly and function of the archaeal flagellum », Biochemical Society Transactions, vol. 39, no 1, , p. 64–9 (DOI 10.1042/BST0390064).
- ↑ Stepanek, Ludek & Pigino, Gaia (2016) Microtubule doublets are double-track railways for intraflagellar transport trains ; 06 mai 2016 ; Vol. 352, n°6286, pp. 721-724 ; DOI: 10.1126/science.aaf4594
- ↑ Lopez,A., avec L. Boissin, « La spermatide d’Holocnemus pluchei (Scop.)(Arachnida, Araneida, Pholcidae) : étude ultrastructurale. », Bull.Soc. zool. France, 101 (3), p. 423-431., .
- ↑ (en) Lopez,A. avec C.Juberthie et J.Kovoor, « Spermiogenesis and spermatophore in Telema tenella Simon (Araneae : Telemidae). An ultrastructural study. », Int. J.Invert.Reprod., 3, 181-191.,
- ↑ Lopez,A. avec J.C. Bonaric et L. Boissin, « Etude ultrastructurale de la spermiogénèse chezl’Araignée Pisaura mirabilis (Clerck, 1758) (Pisauridae). », Rev. Arachnol., 5 (2), p. 55-64., .
Voir aussi
Articles connexes
- Cil cellulaire
- Pilus (ou fimbriae)
- Évolution du flagelle
- Types cellulaires ou organismes unicellulaires possédant un flagelle :
- Spermatozoïdes
- Parasites responsables du mildiou
- Chytridiomycètes
- Certains E. coli
- Dinoflagellé (Microalgues)
- Caryomastigonte
- Unikonta / Bikonta