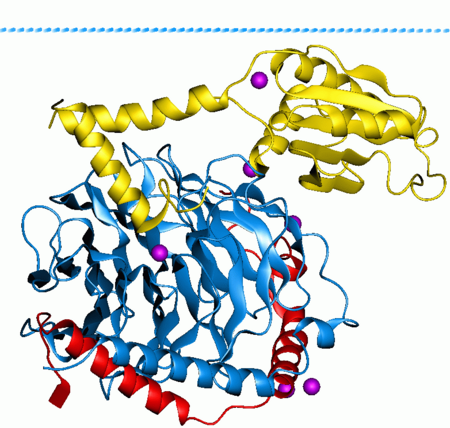
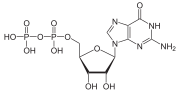

G بروتين (بالانجليزية: G protein) ، والمعروف أيضا باسم بروتين غوانين رابط - النيوكليوتيدات، هي عائلة من البروتينات التي تعمل كمفاتيح جزيئية داخل الخلايا، وتشارك في نقل الإشارات من مجموعة متنوعة من المحفزات خارج الخلية إلى داخلها.[1][2] و يُنظَّم نشاطها من قبل العوامل التي تتحكم في قدرتها على ربط وتحلل ثلاثي فوسفات الغوانوزين (GTP) إلى ثنائي فوسفات الغوانوزين (GDP). عندما تكون مرتبطة بغوانوسين ثلاثي الفوسفات، فإنها تكون 'فعالة'، وعندما تكون مرتبطة بغوانوسين ثنائي الفوسفات، فإنها تكون 'غير فعالة'. البروتينات G تنتمي إلى مجموعة أكبر من الانزيمات تسمى GTPases. هناك نوعان من G بروتين. النوع الأول يعمل كوحدات GTPases صغيرة، في حين أن الوظيفة الثانية كمركب الG بروتين الثلاثي المُغايِر (غيرمتجانس). تتكون الفئة الأخيرة من المركبات من وحدات ألفا (α) و بيتا (β) و غاما .(γ) بالإضافة إلى ذلك، فإن وحدات بيتا وغاما الفرعية يمكن أن تشكل مركب مثنوي مستقر المشار إليه باسم مركب بيتا - غاما. يتم تنشيط Gالبروتين الموجودة داخل الخلية من قبل مستقبلات G- بروتين المقترنة(GPCRs) التي تمتد على غشاء الخلية. ترتبط جزيئات التشوير بنطاق من ال GPCR الموجودة خارج الخلية، وبنطاق من مستقبلات البروتين المقترنة GPCRs) G) داخل الخلية، ثم تنشط بدورها بروتين G معين. وقد تبين أيضا أن بعض المستقبلات التي تقترن ب G بروتين غير المرتبطة بالبروتينات تكون "مقترنة مسبقا" مع G بروتين. ينشط ال G بروتين سلسلة من أحداث التشوير الأخرى التي تؤدي في النهاية إلى تغيير في وظيفة الخلية. مستقبلات البروتين المقترنة GPCRs) G) وال بروتين G تعمل معا بإرسال إشارات من العديد من الهرمونات والناقلات العصبية، وغيرها من عوامل التشوير. البروتينات G تنظم الإنزيمات الأيضية، وقنوات الأيونات، والبروتينات الناقله، وأجزاء أخرى من آلية الخلية، والسيطرة على النسخ، والحركة، والانقباض، والإفراز، والتي تنظم بدورها وظائف النظامية المتنوعة مثل التطور الجنيني والتعلم والذاكرة والتوازن.
تاريخيا
تم اكتشاف البروتينات G عندما قاما ألفريد جيلمان ومارتن رودبيل بالتحقيق في تحفيز الخلايا عن طريق الأدرينالين. ووجد الباحثون أنه عندما يرتبط الأدرينالين بالمستقبلات، فإن المستقبلات لا تحفز الإنزيمات (داخل الخلية) مباشرة. بدلا من ذلك، يحفز مستقبلات البروتين G، الذي يحفز الإنزيم. ومن الأمثلة على ذلك أدينيلات سيكليس (adenylate cyclase)، الذي ينتج المرسل الثاني الحلقي (AMP). لهذا الاكتشاف، فازوا بجائزة نوبل عام 1994 في علم وظائف الأعضاء أو الطب. وقد تم منح جوائز نوبل لكثير من جوانب الإشارات من قبل بروتينات G و مستقبلات G بروتين المقترنة (GPCRs) . وتشمل هذه مضادات المستقبلات، الناقلات العصبية، إعادة امتصاص النواقل العصبية، مستقبلات G بروتين المقترنة (GPCRs) ، وال Gبروتينات، المرسلات الثانويه، والانزيمات التي تؤدي إلى فسفرة البروتين ردا على cAMP، وما يترتب على ذلك من عمليات التمثيل الغذائي مثل انحلال الجليكوجين.
ومن الأمثلة البارزة (بالترتيب الزمني):
- جائزة نوبل في علم وظائف الأعضاء أو الطب لعام 1947 إلى كارل كوري، جيرتي كوري وبرناردو هوساي، لاكتشافهم كيفية تقسيم الجليكوجين إلى الجلوكوز وإعادة تهيئته في الجسم، لاستخدامها كمخزن ومصدر للطاقة. يتم تحفيز تحلل الجليكوجين من قبل العديد من الهرمونات والناقلات العصبية بما في ذلك الأدرينالين.
- جائزة نوبل عام 1970 في علم وظائف الأعضاء أو الطب إلى جوليوس أكسلرود، برنارد كاتس وأولف فون يولر لعملهم على تحرير عن وإعادة الناقلات العصبية. - جائزة نوبل في عام 1971 في علم وظائف الأعضاء أو الطب إلى إيرل سوثيرلاند لاكتشاف الدور الرئيسي للأدينيلات سيكليس (adenylate cyclase) ، التي تنتج الرسول الثاني الحلقي AMP.
- جائزة نوبل عام 1988 في علم وظائف الأعضاء أو الطب لجورج هيتشينغز، جيمس بلاك وجيرترود إليون "لاكتشافهم مبادئ هامة للعلاج من تعاطي المخدرات" تستهدف ال .GPCRs - جائزة نوبل عام 1992 في علم وظائف الأعضاء أو الطب لإدوين كريبس وإدمون فيشر لوصف كيف عكس الفسفرة يعمل كمفتاح لتفعيل البروتينات، وتنظيم العمليات الخلوية المختلفة بما في ذلك تحلل الجليكوجين.
- جائزة نوبل عام 1994 في علم وظائف الأعضاء أو الطب إلى ألفريد جيلمان ومارتن رودبيل لاكتشافهم " ال G بروتين ودور هذه البروتينات في نقل الإشارة في الخلايا". - جائزة نوبل عام 2000 في علم وظائف الأعضاء أو الطب لاريك كاندل، أرفيد كارلسون وبول غرينغارد، للبحث عن الناقلات العصبية مثل الدوبامين، التي تعمل عن طريق ال GPCRs.
- جائزة نوبل عام 2004 في علم وظائف الأعضاء أو الطب لريتشيل أكسيل وليندا باك لعملهما على مستقبلات ال G بروتين الشمية المقترنة. - جائزة نوبل في الكيمياء لعام 2012 لبريان كوبيلكا وروبرت ليفكويتز لعملهم على وظيفة ال GPCRs.
الوظائف
ال Gبروتين هي جزيئات هامة لنقل الإشارة في الخلايا. "اي خلل في مسارات إشارات مستقبلات البروتين المقترنةGPCRs يشترك في العديد من الأمراض مثل السكري والعمى والحساسية والاكتئاب والعيوب القلبية الوعائية وبعض أشكال السرطان. وتشير التقديرات إلى أن حوالي 30٪ من الاهداف الخلوية للعلاجات الدوائية الحديثة هي مستقبلات ال G بروتين المقترنة(GPCRs) ". الجينوم البشري يشفر ما يقرب من 800 مستقبلات البروتين G المقترنةGPCRs ، والتي تكشف فوتونات الضوء والهرمونات وعوامل النمو، والعقاقير، والأربطة الذاتية الأخرى. هناك ما يقرب من 150 من مستقبلات ال G بروتين المقترنةGPCRs وجدت في الجينوم البشري لديها وظائف غير معروفة حتى الآن. في حين يتم تنشيط ال G بروتين من قبل مستقبلات ال G بروتين المقترنة، ويتم تعطيلها من قبل البروتينات RGS "منظم إشارة ال G بروتين ". المستقبلات تعمل على تحفيزارتباط غوانوسين ثلاثي الفوسفات فتقوم بتفعيل ال G بروتين. البروتينات RGS تحفز تحليل غوانوسين ثلاثي الفوسفات (إنتاج غوانوسين ثنائي الفوسفات، وبالتالي يصبح ال G بروتين غير فعال.
أنواع اشارت الG بروتين
ال G بروتين يمكن أن يشير إلى اثنين من عائلات متميزة من البروتينات. ال G بروتين الثلاثية المُغايِرة )غيرمتجانسة) ، التي يشار إليها أحيانا بال G بروتين "الكبيرة"، يتم تنشيطها بواسطة مستقبلات ال G بروتين المقترنة (GPCRs),، وتتألف من وحدات ألفا (α) و بيتا (β) و غاما (γ). ال G بروتين الصغيرة (20-25kDa)تنتمي إلى فصيلة Ras من GTPases الصغيرة. هذه البروتينات متجانسة للوحدة الفرعية ألفا (α) الموجودة في المركبات الثلاثية المُغايِرة (غيرمتجانسة)، ولكنها في الواقع أحادية، تتكون من وحدة واحدة فقط. ومع ذلك، مثل أقربائهم الأكبر، كما أنها تربط غوانوسين ثلاثي الفوسفات وغوانوسين ثنائي الفوسفات وتشارك في نقل الإشارات.
ال G بروتين الثلاثي المُغايِر (غيرمتجانس)
ال G بروتين الثلاثي المُغايِر (غيرمتجانس) أنواع مختلفة من البروتينات ال G بروتين الثلاثية المُغايِرة (غيرمتجانسة) تشترك في آلية مشتركة. يتم تنشيطها في استجابة لتغير بتكوين في مستقبلات ال G بروتين المقترنة (GPCRs) ، لتعمل على تبادل ثنائي فسفات غوانوسين للثلاثي فسفات، والتفكك من أجل تفعيل البروتينات الأخرى في مسارنقل الإشارة. إلا أن الآليات المحددة تختلف بين أنواع البروتينات ال G بروتين الثلاثي المُغايِر (غيرمتجانس) أنواع مختلفة من البروتينات ال G بروتين الثلاثية المُغايِرة (غيرمتجانسة) تشترك في آلية مشتركة. يتم تنشيطها في استجابة لتغير بتكوين في مستقبلات ال G بروتين المقترنة (GPCRs) ، لتعمل على تبادل ثنائي فسفات غوانوسين للثلاثي فسفات، والتفكك من أجل تفعيل البروتينات الأخرى في مسارنقل الإشارة. إلا أن الآليات المحددة تختلف بين أنواع البروتينات
آلالية المشتركة

ترتبط البروتينات G المستقبلة مع السطح الداخلي للغشاء الخلية. وهي تتألف من Gα ووالوحدات الفرعية Gβγ المرتبطة ارتباطا وثيقا. هناك عدة فئات من الوحدات Gαالفرعية ومن بعض الأمثلة: G) Gsα المحفزة) , G) Giα المثبطة),G) Goα وغيرها) ,Gq/11α, G12/13α. أنها تتصرف بشكل مختلف في التعرف على الجزيء المؤثر، ولكنها تشترك بآلية تنشيط مماثلة.
التفعيل
عندما ينشط الربيطة مستقبلات البروتين المقترنة ب G، فإنه يؤدي إلى تغيير تشكلي في المستقبلات التي تسمح للمستقبل بالعمل كعامل تبادل نوكليوتيدات غوانين (GEF) الذي يعمل على تحويل غوانين ثنائي الفوسفات GDP إلى غوانين ثلاثي الفوسفات GTP وبالتالي تحول مستقبلات البروتين المقترنة (GPCRs)إلى " فعاله " . وحدات GTP (أو GDP) مرتبطة بالوحدة الفرعية Gα في وجهة النظر التقليدية لتنشيط ال GPCR الثلاثية المُغايِرة )غيرمتجانسة). يحفز هذا التبادل تفكك الوحدة الفرعية Gα (المرتبطة ب GTP) من ثنائي الوحدات βγ والمستقبل ككل. ومع ذلك، النماذج التي تشير إلى إعادة ترتيب الجزيئي، وإعادة التنظيم، وقبل تعقيد جزيئات المستجيب بدأت تقبل. يمكن لكل من Gα-GTP و Gβγ تنشيط مسارات متتالية مختلفة (أو مسارات مرسول ثانية) وبروتينات المستجيب، في حين أن المستقبلات قادرة على تنشيط بروتين G التالي
انهاء
سوف تحلل الوحدة الفرعية Gα في نهاية المطاف GTP إلى GDP من خلال النشاط الأنزيمي الأصيل، مما يسمح لها باعادة الارتباط مع Gβγ وبدء دورة جديدة. تعمل مجموعة من البروتينات التي تسمى منظم إشارة الG بروتين (RGSs))، بمثابة البروتينات المحفزة ل ,(GAPS) GTPase تعمل مجموعة من البروتينات التي تسمى منظم إشارة الG بروتين (RGSs))، بمثابة البروتينات المحفزة ل GAPS) GTPase) ، والتي هي محددة لوحدات Gα الفرعية. تعمل هذه البروتينات على تسريع التحلل من GTP إلى GDP، وبالتالي إنهاء إلاشارة المنقولة. في بعض الحالات، قد يكون المؤثر نفسه قد يمتلك نشاط GAP جوهري، والتي يمكن أن تساعد بعد ذلك على تعطيل المسار. وهذا صحيح في حالة Phospholipase C-beta، الذي يمتلك نشاط GAP داخل منطقة C-terminal. هذا هو شكل بديل من التنظيم للوحدة Gα. ال Gα GAPs ليس لديها مخلفات حفازة (تسلسل الأحماض الأمينية محددة) لتنشيط البروتين Gα. أنها تعمل بدلا من ذلك عن طريق خفض طاقة التنشيط المطلوبة لبدأ التفاعل .
آليات محددة
Gαs
Gαs ينشط المسار الذي يعتمد على ال cAMP من خلال تحفيز إنتاج AMP حلقي (cAMP) من ادونيسين ثلاثي الفوسفات (ATP). ويتم ذلك من خلال التحفيز المباشر لل انزيم adenylate cyclase الملزم بالغشاء. ثم يمكن لل cAMP بالعمل كمرسل ثاني الذي يكمل ليتفاعل ويحفز بروتين PKA) Kinase A . PKA) يمكن فسفرة عدد لا يحصى من الأهداف المصب. يتم استخدام المسار المعتمد على cAMP كمسار انقال الإشارة للعديد من الهرمونات بما فيها:
ADH - يعزز احتباس الماء عن طريق الكلى (التي أنشأتها خلايا V2 من الغدة النخامية الخلفية)
GHRH - يحفز انتاج واطلاق GH (خلايا سوماتوتروف (somatotroph) من الغدة النخامية الأمامية.
GHIH - يمنع تركيب وإطلاق GH (خلايا سوماتوتروف (somatotroph) من الغدة النخامية الأمامية)
CRH - يحفز انتاج وإطلاق عن ACTH (الغدة النخامية الأمامية)
ACTH - يحفزانتاج وإطلاق عن الكورتيزول (زونا فاسيكولاتا zona fasiculata من قشرة الغدة الكظرية في الغدة الكظرية).
TSH - يحفز انتاج والإفراج عن غالبية T4 (الغدة الدرقية).
LH - ينشط نضوج الجريبي والإباضة في النساء. أو إنتاج هرمون تستوستيرون ونطفة في الرجال
FSH - يحفز التنمية الجريبي في النساء. أو الحيوانات المنوية لدى الرجال
PTH - يزيد من مستويات الكالسيوم في الدم. ويتم ذلك من خلال مستقبلات هرمون الغدة الدرقية 1 (PTH1) في الكلى والعظام، أو عن طريق مستقبلات هرمون الغدة الدرقية 2 (PTH2) في الجهاز العصبي المركزي والدماغ، وكذلك العظام والكلى.
الكالسيتونين(Calcitonin) - يقلل من مستويات الكالسيوم في الدم (عن طريق مستقبلات الكالسيتونين في الأمعاء والعظام والكلى والدماغ)
الجلوكاجون(Glucagon) - يحفز انهيار الجليكوجين في الكبد
hCG - يعزز التمايز الخلوي، ومن المحتمل أن تشارك في موت الخلايا المبرمج.
الأدرينالين – يطلق من النخاع الكظري خلال حالة الصيام، عندما يكون الجسم تحت الإكراه الأيضي. أنه يحفز تحلل الجليكوجين، بالإضافة إلى إجراءات الجلوكاجون.
Gαi
Gαi يمنع إنتاج cAMP من ATP.يعمل الأنسولين من خلال Gi (مثبط) بروتينات المرسل الثانيويه .
Gαq/11
Gαq / 11 يحفز غشاء ملزمة C phospholipase بيتا، الذي يشق PIP2 إلى اثنين من المرسل الثاني، IP3 (Diacylglycerol) DAG يتم استخدام المسار المعتمدInositol Phospholipid كمسار تنبيغ إشارة للعديد من الهرمونات بما في ذلك:
- Vasopressin / AVP) AD): يحفز تخليق والإفراج عن السكرية Zona fasciculate من قشرة الغدة الكظرية في الكلى). يستحث تضيق الأوعية (خلايا V1 من الغدة النخامية الخلفية)
- TRH: يحفز التوليف والإفراج عن TSH (الغدة النخامية الأمامية).
- TSH : يحفز التوليف والإفراج عن كمية صغيرة من T4 (الغدة الدرقية).
- Angiotensin II: يحفز التوليف الألدوستيرون والإفراج عن (zona glomerulosa) من قشرة الغدة الكظرية في الكلى
. - GnRH: يستحث التوليف والإفراج عن FSH و LH (الغدة النخامية الأمامية).
Gα12/13
- Gα12 / 13 تشارك في إشارات عائلة Rho GTPase . هذا هو من خلال عائلة RhoGEF التي تنطوي على المجال RhoGEF من هياكل البروتينات. وتشارك هذه في السيطرة على الخلية الخلوية، وبالتالي في تنظيم هجرة الخلية.
Gβ
- Gβ مركبات ال Gβy لديها أيضاً وظائف نشطة ومن الأمثلة على ذلك اقتران وتفعيل قنوات الGبروتين المقترنة بالبوتاسيوم .
GTPases الصغيرة
GTPases الصغيرة أيضا تربط GTP وGDP وتشارك في نقل الإشارة. هذه البروتينات متماثلة إلى وحدات ألفا (α) الموجودة في المركبات الثلاثية غير المتجانسة، ولكنها موجودة كوحدات احادية. وهي بروتينات صغيرة (20 - كيلو دالتون إلى 25 كيلو دالتون) التي تربط بغوانوسين ثلاثي الفوسفات (GTP),
الربط بليبيد
من أجل ربط هذه البروتينات مع الطبقة الداخلية من غشاء البلازما، العديد من البروتينات G و GTPses الصغيرة مرتبطة مع ليبيدات بروابط تساهمية. الليبيد قد يكون حمض النخيل، حمض الميريستيك، أو مجموعة برينيل.
المراجع
- "معلومات عن بروتين G على موقع wikiskripta.eu". wikiskripta.eu. مؤرشف من الأصل في 13 ديسمبر 2019.
- "معلومات عن بروتين G على موقع thes.bncf.firenze.sbn.it". thes.bncf.firenze.sbn.it. مؤرشف من الأصل في 13 ديسمبر 2019.
روابط خارجية
- GTP-Binding Proteins في المَكتبة الوَطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).