
| Domaine | Eukaryota |
|---|---|
| Règne | Chromalveolata |
| Division | Alveolata |
| Embranchement | Apicomplexa |
| Classe | Aconoidasida |
| Ordre | Haemosporida |
| Famille | Plasmodiidae |
Espèces de rang inférieur
- Voir le texte
Plasmodium est un genre de protozoaires — des organismes unicellulaires — parasites, dont cinq espèces causent le paludisme chez l'être humain.
Propagation
Le parasite est transmis à l’homme par une piqûre d’anophèle (genre de moustique majoritairement présent en régions chaudes). Chez l’être humain et les autres animaux, il infecte en premier lieu les hépatocytes et ensuite, au cours du cycle parasitaire, les érythrocytes. D'autres espèces de Plasmodium ont été retrouvées chez divers animaux endothermes comme les singes, les oiseaux mais aussi ectothermes comme les lézards.
Histoire de la connaissance épidémiologique du plasmodium
- En 1880, Alphonse Laveran découvre Plasmodium falciparum[1]. L'année suivante il publie Nature parasitaire des accidents de l'impaludisme dans lequel il décrit les formes principales sous lesquelles se présente l'hématozoaire[2].
- En 1882, il se rend à Rome pour étudier les paludéens de la campagne romaine. Il découvre dans leur sang le même parasite que celui trouvé à Constantine.
- En 1898, Ronald Ross démontre l'existence de Plasmodium dans l'estomac de l'anophèle[3],[4]. Cette découverte lui vaut le prix Nobel en 1902 tandis qu'Alphonse Laveran le reçoit en 1907 « en reconnaissance de son travail sur le rôle joué par le protozoaire dans la cause des maladies ».
- En 1899, l'Italien Giovanni Battista Grassi démontre que le paludisme ne peut être transmis que par les anophèles via leur système digestif[5].
Espèces impliquées en pathologie humaine
- Plasmodium falciparum (qui cause une forme maligne du paludisme, ou « fièvre tierce maligne » car ayant une périodicité des recrudescences fébriles tous les deux jours).
- Plasmodium vivax (qui cause une forme bénigne du paludisme ou « fièvre tierce » car ayant une périodicité des recrudescences fébriles tous les deux jours).
- Le plus rare, Plasmodium ovale (qui cause une forme bénigne du paludisme ou « fièvre tierce » car ayant une périodicité des recrudescences fébriles tous les deux jours), n'est rencontré que dans certaines zones de l'Afrique intertropicale.
- Plasmodium malariae (qui est la cause du paludisme bénin ou « fièvre quarte » car ayant une périodicité des recrudescences fébriles tous les trois jours).
- Plasmodium knowlesi, proche génétiquement de Plasmodium vivax, et, microscopiquement, de Plasmodium malariae. Il a été découvert récemment chez l'Homme en Malaisie (mais était connu antérieurement chez le singe).
Morphologie microscopique
Il se présente sous la forme d'un protozoaire très petit (1 à 2 µm selon les formes) ; la coloration au May-Grünwald-Giemsa montre qu'il est constitué d'un cytoplasme bleu pâle entourant une vésicule nutritive de teinte claire, et contenant un noyau rouge et du pigment brun-doré ou noir (hémozoïne).
Biologie du plasmodium
Le cycle évolutif assez complexe nécessite deux hôtes : l'homme et la femelle hématophage d'un moustique du genre anophèle[6]. D'un point de vue strictement biologique, le véritable hôte définitif de cet hématozoaire est le moustique (la reproduction sexuée parasite l'anophèle ). L'homme et les autres vertébrés ne sont que des hôtes intermédiaires dans son cycle réplicatif. Néanmoins, pour des raisons anthropocentriques, on considère que le vecteur n'est pas l'homme mais le moustique.
Mince fuseau de 12 µm / 1 µm, le sporozoïte infectieux est injecté à l'homme lors de la piqûre d'une femelle d'anophèle infectieuse. Il atteint le foie en 1⁄2 heure et pénètre dans un hépatocyte : c'est le début d'une crise pré-érythrocytaire hépatique qui va durer 6 jours pour P. falciparum, 8 jours pour P. vivax, 9 jours pour P. ovale et probablement 12 jours pour P. malariae. Le plasmodium cryptozoïte va subir dans l'hépatocyte une intense multiplication asexuée aboutissant au corps bleu, énorme schizonte mûr de 40 à 80 µm. Ce corps bleu bourgeonne alors, développant des vésicules qui contiennent les jeunes mérozoïtes. Ces vésicules faits de membrane morte ne sont pas phagocytées par les macrophages en raison d'une action biochimique des parasites sur la membrane qui supprime les signaux de destruction phagocytaire des cellules mortes. Ces vésicules sont libérées dans les sinusoïdes hépatiques pour rejoindre ensuite la circulation sanguine.
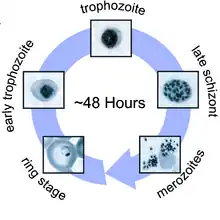
C'est une véritable technique de cheval de Troie qui est ici utilisée pour passer des cellules hépatiques au sang. Les vésicules y libèrent alors leur flot de jeunes mérozoïtes pré-érythrocytaires qui vont s'y répandre. C'est le début de la longue phase sanguine : les mérozoïtes s'accolent aux érythrocytes (margination), les envahissent, s'y développent (trophozoïtes) puis s'y divisent (schizontes). L'éclatement des schizontes mûrs ou « rosaces » termine le premier cycle schizogonique érythrocytaire en libérant dans le sang, avec les déchets du métabolisme plasmodial (pigments et débris cellulaires), une nouvelle génération de plasmodiums, les mérozoïtes érythrocytaires.
Une succession régulière de cycles semblables va suivre, qui sera progressivement remplacée, les défenses immunitaires s'organisant, par des cycles érythrocytaires gamogoniques préparant les formes sexuées : les trophozoïtes, au lieu de se diviser, modifient leur rapport nucléo-plasmatique et donnent des formes à noyau volumineux et cytoplasme densifié, les gamétocytes mâles et femelles, qui vont demeurer en attente dans le sang circulant. Déglutis par une femelle anophèle lors d'un repas sanguin , ces gamétocytes terminent le cycle sexué : devenus gamètes mâles et femelles ils se conjuguent donnant l'ookynète, qui après avoir traversé la paroi intestinale, s'arrête sur la face externe de l'epithelium, devient oocyste puis sporocyste ; son éclatement libère les sporozoïtes infectieux qui s'accumulent, en attente, dans les glandes salivaires, rendant ainsi la femelle d'anophèles infectieuse. Lorsque cette femelle piquera un homme, elle injectera les sporozoïtes avec sa salive et le cycle évolutif sera bouclé.
Pour P. falciparum, c'est là toute l'évolution biologique. Chez P. vivax, P. malariae et P. ovale, certains mérozoïtes pré-érythrocytaires ne gagnent pas le sang, mais s'attaquent à de nouveaux hépatocytes : c'est le début des cycles exo-érythrocytaires secondaires qui vont entretenir dans le foie la parasitose pendant 3 à 5 ans ou plus pour P. vivax, 2 ou 3 ans pour P. ovale et pendant la vie entière pour P. malariae.
Le cycle de Plasmodium est très complexe. Il démarre chez l'homme par la piqûre d'un moustique femelle du genre anophèle ou, plus rarement, par transfusion de sang déjà contaminé. Ces sporozoïtes migrent vers le foie via la circulation sanguine ou lymphatique où ils vont envahir les hépatocytes. Ils se différencient alors en schizonte hépatique qui relarguent des mérozoïtes dans le sang. Chez certaines espèces (P. ovale et P .vivax), un stade cryptozoïte reste caché dans le foie, le parasite se réveille alors plusieurs mois ou années plus tard pour reprendre son cycle. Les mérozoïtes infectent ensuite les globules rouges où ils se différencient sous la forme des stades anneau et amiboïde (trophozoïtes). Les parasites actifs (mérozoïtes) qui sont alors produits quittent l'érythrocyte et voyagent avec le sang pour envahir de nouveaux érythrocytes. Après que les mérozoïtes ont envahi de nouvelles hématies, les stades sexuels (gamétocytes) sont produits et peuvent être aspirés par un moustique anophèle femelle. Ils forment des zygotes qui se divisent pour produire de grands nombres de minuscules sporozoïtes. Les sporozoïtes (qui est le stade infectieux) migrent vers les glandes salivaires du moustique où ils sont injectés dans le sang de l'hôte suivant. Les sporozoïtes se meuvent alors dans le foie où ils répètent le cycle. Les fièvres de la malaria sont dues aux activités des parasites dans le sang qui cassent les cellules et produisent des substances toxiques. À cause de la complexité du cycle de vie de ces parasites, il est difficile de développer un vaccin.
Une étude[7] de l'Unité de Biologie et Génétique du Paludisme de l'Institut Pasteur, publiée en , montre que le parasite peut aussi se reproduire dans la peau et dans le follicule pileux.
Liste des espèces
- P. achiotense
- P. aegyptensis
- P. aeuminatum
- P. agamae
- P. anasum
- P. atheruri
- P. azurophilum
- P. balli
- P. bambusicolai
- P. basilisci
- P. berghei
- P. bigueti
- P. brasilianum
- P. brygooi
- P. booliati
- P. bubalis
- P. bucki
- P. coatneyi
- P. cathemerium
- P. cephalophi
- P. chabaudi
- P. chiricahuae
- P. circularis
- P. cnemidophori
- P. coatneyi
- P. coggeshalli
- P. colombiense
- P. corradettii
- P. coturnix
- P. coulangesi
- P. cuculus
- P. cyclopsi
- P. cynomolgi
- P. diminutivum
- P. diploglossi
- P. dissanaikei
- P. dominicana
- P. durae
- P. egerniae
- P. elongatum
- P. eylesi
- P. fabesia
- P. fairchildi
- P. falciparum
- P. fallax
- P. fieldi
- P. foleyi
- P. forresteri
- P. floridense
- P. fragile
- P. gaboni
- P. garnhami
- P. gallinaceum
- P. giganteum
- P. giovannolai
- P. girardi
- P. gonatodi
- P. gonderi
- P. georgesi
- P. gracilis
- P. griffithsi
- P. guanggong
- P. gundersi
- P. guyannense
- P. heischi
- P. hegneri
- P. hermani
- P. heteronucleare
- P. hexamerium
- P. holaspi
- P. huffi
- P. hylobati
- P. icipeensis
- P. inopinatum
- P. inui
- P. jefferi
- P. josephinae
- P. juxtanucleare
- P. kempi
- P. knowlesi
- P. kentropyxi
- P. leanucteus
- P. lemuris
- P. lophurae
- P. lepidoptiformis
- P. lygosomae
- P. mabuiae
- P. mackerrasae
- P. maculilabre
- P. maior
- P. malariae
- P. marginatum
- P. matutinum
- P. mexicanum
- P. minasense
- P. morulum
- P. nucleophilium
- P. octamerium
- P. odocoilei
- P. ovale
- P. Papernai
- P. paranucleophilum
- P. parvulum
- P. pedioecetii
- P. pelaezi
- P. percygarnhami
- P. petersi
- P. pifanoi
- P. pinotti
- P. pinorrii
- P. pitheci
- P. pitmani
- P. polare
- P. praecox
- P. reichenowi
- P. relictum
- P. rhadinurum
- P. rhodaini
- P. robinsoni
- P. rouxi
- P. sandoshami
- P. sasai
- P. schweitzi
- P. silvaticum
- P. simium
- P. semiovale
- P. shortii
- P. Smirnovi
- P. subpraecox
- P. tenue
- P. tejerai
- P. tomodoni
- P. torrealbai
- P. traguli
- P. tribolonoti
- P. tropiduri
- P. uilenbergi
- P. watteni
- P. wenyoni
- P. vacuolatum
- P. vastator
- P. vaughani
- P. vinckei
- P. vivax
- P. volans
- P. yoelii
- P. youngi
Notes et références
- ↑ Laveran A., « Un nouveau parasite trouvé dans le sang de malades atteints de fièvre palustre. Origine parasitaire des accidents de l’impaludisme. », Bull Mém Soc Méd Hôpitaux Paris, 2e série, vol. 17, , p. 158-164 (ISSN 0366-1334, BNF 34378153, lire en ligne, consulté le )
- ↑ Alphonse Laveran, Nature parasitaire des accidents de l'impaludisme : Description d'un nouveau parasite trouvé dans le sang des malades atteints de fièvre palustre., Paris, J.-B. Baillière, , 101 p. (BNF 30755395, lire en ligne)
- ↑ Ronald Ross, « The role of the mosquito in the evolution of the malarial parasite: the recent researches of Surgeon-Major Ronald Ross, I.M.S. 1898 », The Yale Journal of Biology and Medicine, vol. 75, no 2, , p. 103–105 (ISSN 0044-0086, PMID 12230308, PMCID PMCPMC2588727, lire en ligne, consulté le )
- ↑ P. Manson, « SURGEON-MAJOR RONALD ROSS'S RECENT INVESTIGATIONS on the MOSQUITO-MALARIA THEORY », British Medical Journal, vol. 1, no 1955, , p. 1575–1577 (ISSN 0007-1447, PMID 20757898, PMCID PMCPMC2411754, lire en ligne, consulté le )
- ↑ (it) Grassi B. et Bignami A., « Ulteriori ricerche sul ciclo dei parassiti malarici umani nel corpo del zanzarone », Atti Della Reale Accademia Nazionale Dei Lincei. Classe Di Scienze Fisiche, Matematiche e Naturali. Rendiconti., Tip. della R. Accademia dei Lincei, 5e série, vol. 8, , p. 21-28 (ISSN 1124-5042, lire en ligne, consulté le )
- ↑ Elamaran Meibalan et Matthias Marti, « Biology of Malaria Transmission », Cold Spring Harbor Perspectives in Medicine, vol. 7, no 3, (ISSN 2157-1422, PMID 27836912, PMCID PMCPMC5334247, DOI 10.1101/cshperspect.a025452, lire en ligne, consulté le )
- ↑ Pascale Gueirard, Joana Tavares, Sabine Thiberge, Florence Bernex, Tomoko Ishino, Genevieve Milon, Blandine Franke-Fayard, Chris J. Janse, Robert Ménard & Rogerio Amino ; Development of the malaria parasite in the skin of the mammalian host, PNAS, 4 octobre 2010 [(en) lire en ligne].
Voir aussi
Bibliographie
- (en) Virginia M. Howick, Andrew J. C. Russell, Tallulah Andrews, Haynes Heaton, Adam J. Reid et al., « The Malaria Cell Atlas: Single parasite transcriptomes across the complete Plasmodium life cycle », Science, vol. 365, no 6455, , article no eaaw2619 (DOI 10.1126/science.aaw2619)
Liens externes
- Malaria Atlas Project
- Cours de microbiologie eucaryote : les Apicomplexans
- Plasmodium sur MicrobeWiki
- Ressources relatives au vivant :
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :