الانحراف الوراثي أو الانحراف الأليلي (Genetic drift) هي قوة تطورية تعمل مع الاصطفاء الطبيعي عن طريق تغيير خصائص الأنواع على مر الزمن. هو غالباً ما يعرف بالتغير العشوائي في تواترات الأشكال المغايرة للجينات (ألائل) الذي يحدث نتيجة لخطأ الاستعيان (الخطأ في تمثيل العينة للكل).[1] فجزء معين فقط من مجموع الألائل الموجودة في جيل الآباء تنتقل إلى جيل الأبناء. الألائل الموجودة لدى الأبناء هي عينة عن الألائل الموجودة لدى الآباء، ولها دور في تحديد ما إذا كان الفرد سينجو ويتكاثر. تواتر أليل معين في التجمع هو النسبة التي تشكلها نسخ هذا الأليل.[2] وعندما يصل تواتر الأليل إلى صفر، فهذا يعني أنًّ هذا الأليل اختفى ولم يعد متوفرا. قد يؤدي الانحراف الوراثي إلى اختفاء الألائل كلياً، وبذلك يقلل من التنوع الجيني.
تأثير الانحراف الجيني يكبر مع تضاؤل عدد نسخ الأليل، ويصغر عندما تتواجد نسخاً كثيرة من الأليل. وقد دارت جدالات شديدة حول مدى أهمية الاصطفاء الطبيعي مقابل السيرورات التطورية الحيادية الأخرى، ومنها الانحراف الوراثي. ورأى عالم الأحياء التطوري رونالد فيشر أنَّ الانحراف الوراثي، على أكثر تقدير، يلعب دوراً صغيراً في تحريك التطور. وهذا الاعتقاد بقي هو السائد لعدة عقود. وفي عام 1968 أشعل Mooto Kimura الجدال من جديد بنظرية التطور الجزيئي الحيادية، التي تدعي أنَّ معظم الحالات التي ينتشر فيها التغير الجيني عبر التجمع (ليس بالضرورة تغيرا بالنمط الظاهري) يكون سببها الانحراف الوراثي.[3]
وقد أشار المسعودي في كتابه مروج الذهب إلى الانحراف الوراثي الذي حدث في الحمضيات أثناء نقلها من السند
إلى مصر، وسجل هذا الانحراف على أصناف من الليمون.
محاكاة الكريات في الجرة
يمكن إيضاح عملية الانحراف الوراثي بواسطة عشرين كرية في جرة لتمثل عشرين كائنا حيا في تجمع.[4] وذلك باعتبار الجرة الحاوية للكريات الجيل الأول للتجمع. الكريات في الجرة نصفها أحمر اللون ونصفها الآخر أزرق. واللونان يمثلان أليلين مختلفين لجين معين في التجمع. وفي كل جيل يتكاثر أفراد التجمع بشكل عشوائي. تتم محاكاة هذا التكاثر عن طريق اختيار كرية عشوائية من الجرة، ووضع كرية إضافية بنفس اللون في جرة أخرى (الكرية التي تم اختيارها تعاد إلى الجرة الأولى). وتكرر هذه العملية حتى يصل عدد الكريات في الجرة الثانية إلى عشرين. الجرة الثانية الآن تحتوي على كريات ذات ألوان مختلفة ممثلةً للجيل الثاني، جيل الأبناء. وفي حال لم تكن في الجرة عشر كريات حمراء وعشر زرقاء بالضبط، فإنَّه حدث تغيير عشوائي في تواترات الكريات الحمراء والزرقاء، وبكلمات أخرى: حدث انحراف "وراثي".
مع تكرار العملية عدة مرات، يتم إنتاج "أجيال" لاحقة من الكريات بشكل عشوائي. عدد الكريات الحمراء والزرقاء في كل جيل "ينحرف": أحياناً يكون عدد الحمراء أكبر من الزرقاء، وأحيانا العكس. وهذا الانحراف يناظر الانحراف الوراثي - تغير في تواترات ألائل التجمع ينتج عن التغير العشوائي في توزيع الجينات من جيل لآخر يليه.
تقنية الانحراف الوراثي
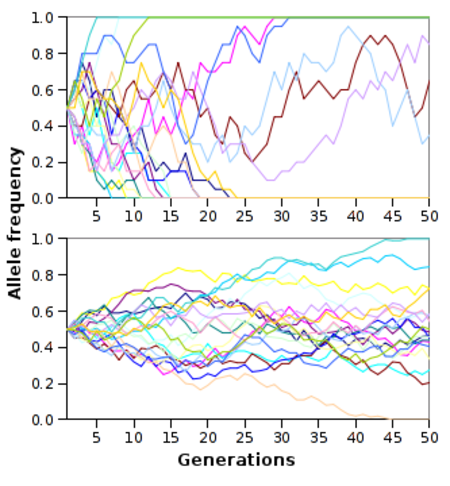
الانحراف الوراثي هو تغيير عشوائي وبارز في تواتر الأليلات من جيل إلى جيل. الانحراف الوراثي يحدث تقريباً فقط في المجموعات السكانية الصغيرة. يحدث هذا بسبب، أن القانون الضعيف للأعداد الكبيرة ينص، أنه في النماذج الصغيرة هنالك احتمال جيد بأن عمليات ذات احتمال متساوي (مثل توريث كل واحد من الأليلات متساوية التوزيع), لن تحدث بعدد متساوي من الإمكانيات. على سبيل المثال: في مجموعة سكانية ذات 10 عائلات وتوزيع متساوي للأليلات التي تعطي عيناً زرقاء والأليلات التي تعطي عيناً بنية، هنالك احتمال جيد لجيل الآباء أن يورث بالصدفة أليلات من النوع التي تعطي عيوناً زرقاء أكثر من النوع الآخر لجيل الأبناء. وهذا بالتناقض مع أن مجموعة سكانية بشرية والتي تضم 10,000 عائلة هنالك احتمال توريت غير متساوي هو قريب للصفر.
أثر المؤسس
- مقالة مفصلة: تأثير المؤسس
أثر المؤسس - أو تأثير المؤسس - هو حالة من الانحراف الوراثي التي تحصل بسبب هجرة وتأسيس مجموعة سكانية جديدة. في هذه الحالة تغير أليلات معينة توزيعها نسبة للتوزيع الذي كان في المجموعة السكانية الاصلية بسبب أنه في مجموعة المهاجرين الصغيرة والي أسست المجموعة السكانية الجديدة كانوا بالصدفة ذوي توزيع غير طبيعي. على سبيل المثال، قبل اكتشاف أمريكا
على يد كولومبوس، كان سكان أمريكا ذوي دم من نوع O, وهذا بالتناقص مع باقي سكان العالم الذين كانوا ذوي دم من نوع آخر. من الممكن أن هذا حدث بسبب أن المهاجرين الأوائل كانوا بالصدفة ذوي دم من نوع O.
نتائج الانحراف الوراثي
هنالك نتيجتان لعمليلات الانحراف الوراثي وهي:
- ترسيخ عشوائي للأليلات - وهذا يحدث حين تتكون مجموعة سكانية صغير لأي سبب كان، انتشار الأليلات المختلفة فيها لكل جين، من الممكن أن تكون مختلفةً تماماً عن المجموعة السكانية الاصلية. زيادة على ذلك، كلما كانت المجموعة السكانية صغيرة أكثر، تترسخ فيها أليلات بشكل عشوائي بدون علاقة لمدى الفاعلية أو الضرر التي تسببها هذه الأليلات (فيما عدا الأليلات القاتلة).
- فقدان اللاتجانس - كلما كانت أليلات معينة تميل لأن تتعرض لترسيخ أو انقراض في المجموعة السكانية، عدد الأفراد اللامتجانسين لجين معين يصغر. نتيجة لهذا من الممكن أن يتعرض هذا الجين لطفرات متنحية مضرة والتي قبل هذا لم تحدث في الأفراد الذين كانوا لامتجانسين بالنسبة لهذه الصفة - الوضع الذين من الممكن أن يؤدي لأضرار كبيرة في هذه المجموعة السكانية لهذا النوع.
أمثلة لبعض التطبيقات في الهندسة الوراثية
الخنزير المحور جينيا: هو خنزير غرس فيه هرمون النمو البشري. ينمو بسرعة فائقة ولكنه "أحول" يسير بطريقة فوضوية وغير منتظمة، يعاني من تقرح في المعدة والتهاب في المفاصل وتعفن في الجهاز التنفسي وأمراض الكلى وتشوهات في المفاصل. غرس وإدماج مورثة هرمون النمو لم يحدث فقط التغيرات المرجوة بل وأيضا تغيرات غير متوقعة وغير مرغوب فيها. يفسر علماء الوراثة ذلك بمفعول موقع المورثة: الذي يؤثر على كامل الرصيد الوراثي. هندسة الجينات "تقنية" حديثة ورغم ذلك فالبعض يتمادى في المغامرة ويتلاعب بالكائنات الحية. في حالة الخنزير المحور جينيا كان الهدف منه الحصول على خنزير ينمو بسرعة ويتجاوز وزنه المعدل العام وذلك للحصول على كمية أوفر من اللحم. أي أن الغاية كانت تجارية بحتة. النتيجة كانت عكسية مما دفع الخبراء إلى التخلي عن هذه التجربة.
(التلقيح) مانع الحمل لدى الفئران: أراد بعض الخبراء الأستراليين صنع تلقيح مانع للحمل لدى الفئران. استعملوا فيروس الجدري الخاص بالفئران ليقوم هذا الأخير بنقل مورثة اختيرت لهذا الغرض وذلك إلى نواة خلايا الفئران. فجأة تحول الفيروس الوديع إلى فيروس قاتل. ماتت الفئران في ظرف 9 أيام. هرع الخبراء وخصوصا وأنه بإمكان هذا الفيروس أن ينفلت من داخل المخبر ويسبب كارثة بيئية. تدخلت السلطات الأسترالية الرسمية مباشرة وأتلفت كل أجهزة المخبر. إلى يومنا هذا لم يعرف العلماء سبب تحول الفيروس "الوديع" إلى فيروس قاتل، "وربما تكون طفرة قد سببت ذلك".
الانحراف الوراثي مقابل الاصطفاء الطبيعي
في الجمهرات الطبيعية، لا يؤثر الانحراف الوراثي والاصطفاء الطبيعي بمعزل عن بعضهما، إذ يكون حضورهما معاً دائماً، إلى جانب الطفرات والهجرة، فيكون التطور المُحايد نتاجاً للطفرات والانحراف معاً، وليس الأخير بمفرده، وبشكل مشابه، حتى إذا هيمن الاصطفاء على الانحراف الوراثي، فإنه لن يتمكن من التأثير إلا على المُغايرات التي توفرها الطفرات.
وبينما يوجّه الاصطفاء الطبيعي التطور نحو التكيفات الوَروثة مع البيئة الحالية، لا يمتلك الانحراف الجيني أية وجهة، وإنما تقوده الاحتمالات الرياضية فقط،[5]فيؤثر كنتيجة لذلك على تواترات النمط الجيني ضمن جمهرة ما بغض النظر عن التأثيرات على النمط الظاهري.
أما الاصطفاء فيفضل في المقابل انتشار الألائل التي تزيد تأثيراتها على النمط الظاهري من بُقيا و/أو تكاثر حامليها، ويقلل من تواتر الألائل المسؤولة عن صفات غير مرغوبة، ويتجاهل تلك الحيادية منها.[6]
ويتوقع قانون الأعداد الكبيرة أنه عندما يكون العدد المُطلق لنسَخ الأليل صغيراً (كما في الجمهرات الصغيرة)، يكون مقدار الانحراف على تواترات هذا الأليل في الجيل الواحد أكبر، ويكون مقدار الانحراف كبيراً بما يكفي ليتفوق على الاصطفاء عند تواتر أي من الألائل عندما يكون معامل الاصطفاء أقل من 1 بالتقسيم على الحجم الفعال للعينة.
وبالتالي يُعتبر التطور غير القابل للتكيف والناتج عن فعل الطفرات والانحراف الوراثي آليةً ناتجة بشكل طبيعي عن التغير الوراثي بشكل أساسي ضمن الجمهرات الصغيرة والمنعزلة.[7]
وتعتمد رياضيات الانحراف الوراثي على الحجم الفعال للجمهرة، لكن آلية تعلّق ذلك بالعدد الفعلي للأفراد في الجمهرة غير واضحة،[8] ويمكن أن يخفّض الارتباط الجيني بالجينات الأخرى الخاضعة للاصطفاء من الحجم الفعال للجمهرة التي تمتلك أليلاً حيادياً، ومع تزايد معدل التأشيب، يتناقص هذا الارتباط ومعه التأثير الموضعي على الحجم الفعال للجمهرة.[9][10]
ويُشاهد هذا التأثير في البيانات الجزيئية كترابط بين التأشيب الموضعي والتنوع الوراثي،[11] وترابط سلبي بين الكثافة والتنوع الجينيين في أجزاء الـ(DNA) غير المرمزة،[12] ويختلف التسلسل العشوائي المترافق مع الارتباط بالجينات الأخرى الخاضعة للاصطفاء عن خطأ الاعتيان، ويُعرف أحياناً بالترافق الجيني (genetic draft) لتمييزه عن الانحراف الوراثي (genetic drift).[8]
عندما يكون تواتر الأليلات صغيراً للغاية، يمكن للانحراف أن يتغلب على الاصطفاء حتى في الجمهرات الكبيرة أيضاً، فعلى سبيل المثال، في الوقت الذي تُلغى فيه الطفرات غير الملائمة بسرعة عادةً في الجمهرات الكبيرة، تكون الطفرات المفيدة الجديدة عرضة للفقدان من خلال الانحراف الوراثي بقدر تلك المحايدة تقريباً، ولن ينعدم تأثير الانحراف الوراثي حتى يصل تواتر أليل الطفرة المفيدة لعتبة معينة.[6]
عنق الزجاجة السكانية
يتمثّل نموذج عنق الزجاجة السكانية في تقلص جمهرة ما لحجم أصغر بكثير على مدى فترة قصيرة من الزمن نتيجة حدث بيئي عشوائي، ويكون احتمال بُقيا أي فرد من الجمهرة في نموذج حقيقي لعنق الزجاجة السكانية عشوائياً صرفاً، ولا يتحسّن بأي ميزة جينية موروثة معينة، وقد يؤدي هذا النموذج لتغيرات جذرية في تواترات الألائل بشكل مستقل تماماً عن الاصطفاء.[13]
ويمكن أن يكون تأثير عنق الزجاجة السكانية مستداماً، حتى وإن نتج عن حدث طارئ لمرة واحدة، ككارثة طبيعية على سبيل المثال، ومن الأمثلة المثيرة للاهتمام عن تسبب عنق الزجاجة بتوزع جيني غير اعتيادي، العدد الكبير نسبياً للأفراد المصابين بعمى ألوان تام ضمن آتول بينجلاب المرجاني التابع لولايات ميكرونيسيا المتحدة.
وبعد هيمنة نموذج عنق الزجاجة يتزايد زواج الأقارب، وهو ما يفاقم الضرر الذي تسببه الطفرات المتنحية المؤذية بعملية تُعرف بانحدار زواج الأقارب، ويتم تجنب اصطفاء أسوأ هذه الطفرات، ما يؤدي لفقدان الألائل الأخرى المتعلقة جينياً بها، في عملية تُدعى باختيار الخلفية،[1] وبالنسبة للطفرات المتنحية المؤذية فقد يتعزز هذا الاختيار كنتيجة لعنق الزجاجة، بسبب التظهير الجيني (تناقص تواتر ألّيل ضار نتيجة ازدياد فعالية الاصطفاء الطبيعي المحفّز بزواج الأقارب)، وهو ما يؤدي بدوره للمزيد من خسارة التنوع الوراثي.
يمكن لنموذج عنق الزجاجة أن يُنقص من التنوع الوراثي لجمهرة ما بشكل كبير، حتى أنه قد يلغي التكيفات المفيدة بشكل دائم،[14] علماً أن فقدان التنوع يترك الجمهرة الناجية عرضة لأية ضغوطات اصطفائية جديدة، كالمرض، والتغير المناخي وتغير موارد الطعام المتاحة، لأن التكيّف استجابةً للتغيرات البيئية يتطلب تنوعاً وراثياً كافياً في الجمهرة حتى يتسنى للاصطفاء الطبيعي الحدوث.[15][16]
وُجدت العديد من الحالات المعروفة لعنق الزجاجة السكانية في الماضي القريب، فقبل وصول الأوروبيين لأمريكا الشمالية، كانت البراري فيها موطناً لملايين حيوانات الطهيوج ريشي الشكل، ففي إلينوي لوحدها تراجعت أعداد هذه الكائنات من حوالي 100 مليون طير عام 1900 لحوالي 50 طير فقط في تسعينيات القرن ذاته، وذلك نتيجة صيدها وتدمير مواطنها، ما أدّى لفقدان هذا النوع معظم تنوعه الوراثي، ويوثق تحليل الـ(DNA) المقارن لطيور أواسط القرن العشرين مع طيور تسعينيات القرن ذاته انحداراً سحيقاً في التنوع الوراثي خلال هذه العقود المعدودة لوحدها، ويعاني الطيهوج ريشي الشكل حالياً من انخفاض نجاحه التناسلي.[17] في المقابل قد تزيد الخسارة الجينية التي يسببها عنق الزجاجة والانحراف الوراثي من الصلاحية، كما في حالة الجراثيم الإيرليخية.[18]
سبب الصيد الجائر في القرن التاسع عشر نموذج عنق زجاجة سكانية شديداً لدى فيل البحر الشمالي أيضاً، ويمكن استنتاج انحدار التنوع الجيني الناتج لدى هذه الكائنات بمقارنتها مع ذاك الخاص بفيل البحر الجنوبي الذي لا يجري اصطياده بشدة كبيرة.[19]
مراجع
- Masel J (2011). "Genetic drift". Current Biology. 21 (20): R837–R838. doi:10.1016/j.cub.2011.08.007. PMID 22032182. مؤرشف من الأصل في 11 ديسمبر 2018.
- Futuyma, Douglas (1998). Evolutionary Biology. Sinauer Associates. صفحة Glossary. .
- Futuyma, Douglas (1998). Evolutionary Biology. Sinauer Associates. صفحة 320. .
- "Evolution 101:Sampling Error and Evolution". University of California Berkeley. مؤرشف من الأصل في 10 يوليو 201501 نوفمبر 2009.
- "Natural Selection: How Evolution Works". Actionbioscience. Washington, D.C.: American Institute of Biological Sciences. مؤرشف من الأصل في 6 يناير 201024 نوفمبر 2009. An interview with Douglas J. Futuyma. See answer to question: Is natural selection the only mechanism of evolution?
- Cavalli-Sforza, Menozzi & Piazza 1996
- Zimmer 2001
- Gillespie JH (November 2001). "Is the population size of a species relevant to its evolution?". Evolution; International Journal of Organic Evolution. جون وايلي وأولاده for the Society for the Study of Evolution. 55 (11): 2161–9. doi:10.1111/j.0014-3820.2001.tb00732.x. PMID 11794777.
- Golding 1994، صفحة 46
- Charlesworth B, Morgan MT, Charlesworth D (August 1993). "The effect of deleterious mutations on neutral molecular variation" ( كتاب إلكتروني PDF ). Genetics. Genetics Society of America. 134 (4): 1289–303. PMC . PMID 8375663. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 12 مارس 2020.
- Presgraves DC (September 2005). "Recombination enhances protein adaptation in Drosophila melanogaster". Current Biology. Cell Press. 15 (18): 1651–6. doi:10.1016/j.cub.2005.07.065. PMID 16169487.
- Nordborg M, Hu TT, Ishino Y, Jhaveri J, Toomajian C, Zheng H, Bakker E, Calabrese P, Gladstone J, Goyal R, Jakobsson M, Kim S, Morozov Y, Padhukasahasram B, Plagnol V, Rosenberg NA, Shah C, Wall JD, Wang J, Zhao K, Kalbfleisch T, Schulz V, Kreitman M, Bergelson J (July 2005). "The pattern of polymorphism in Arabidopsis thaliana". PLoS Biology. المكتبة العامة للعلوم. 3 (7): e196. doi:10.1371/journal.pbio.0030196. PMC . PMID 15907155.

- Robinson R, المحرر (2003). "Population Bottleneck". Genetics. 3. New York: Macmillan Reference USA. . LCCN 2002003560. OCLC 614996575. مؤرشف من الأصل في 3 مارس 201614 ديسمبر 2015. نسخة محفوظة 9 يونيو 2019 على موقع واي باك مشين.
- Futuyma 1998، صفحات 303–304
- O'Corry-Crowe G (March 2008). "Climate change and the molecular ecology of Arctic marine mammals". Ecological Applications. Ecological Society of America. 18 (2 Suppl): S56-76. doi:10.1890/06-0795.1. PMID 18494363.
- Cornuet JM, Luikart G (December 1996). "Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data". Genetics. Genetics Society of America. 144 (4): 2001–14. PMC . PMID 8978083.
- Sadava et al. 2008، chpts. 1, 21–33, 52–57
- Dale C, Moran NA (August 2006). "Molecular interactions between bacterial symbionts and their hosts". Cell. 126 (3): 453–65. doi:10.1016/j.cell.2006.07.014. PMID 16901780.
- "Bottlenecks and founder effects". Understanding Evolution. University of California, Berkeley. مؤرشف من الأصل في 4 ديسمبر 201514 ديسمبر 2015.