|
عريضات الأجنحة العصر: الأوردوفيشي–البرمي | |
|---|---|
 |
|
| المرتبة التصنيفية | رتبة، وطويئفة، وطائفة |
| التصنيف العلمي | |
| فوق النطاق | حيويات |
| مملكة عليا | حقيقيات النوى |
| مملكة | حيوان |
| عويلم | ثنائيات التناظر |
| مملكة فرعية | أوليات الفم |
| شعبة عليا | انسلاخيات |
| شعبة | مفصليات الأرجل |
| شعيبة | كلابيات القرون |
| طائفة | وركيات الفم |
| رتبة | سيفيات الذيل |
| رتبة | Xiphosurida |
| الاسم العلمي | |
| Eurypterida هيرمان بورمايستر ، 1843 |
|
عريضات الأجنحة (الاسم العلمي: Eurypterids) من اليونانية (eury) = "عريض" + (pteron) = "جناح". (وتسمى عقارب البحر وهي مجموعة عملاقة منقرضة من شعبة مفصليات الأرجل والمتصلة بالعنكبوتيات والتي تتضمن أكبر مجموعة مفصليات الأرجل المعروفة التي عاشت على الإطلاق. وهم أعضاء في الرُتبة العملاقة اليوريبتيريدات المنقرضة (Eurypterida) (الكلابيات (Chelicerata)) وهي أكثر رتبة كلابيات متنوعة من العصر الباليوزي فيما يتعلّق بالأنواع.[1] ويُشتق الاسم Eurypterida من الكلمة اليونانية eury- والتي تعني "واسع" أو "عريض" والكلمة اليونانية pteron والتي تعني "جناح"،[2] إشارة إلى زوج اللواحق العريض للسباحة على أول حفرية لليوريبتيريدات تُكتشف. ترجع اليوريبتيريدات إلى الأسماك الأولى. وقد وصل أكبر جيكيلوبتروس (Jaekelopterus)، إلى 2.5 متر (8 قدم 2 بوصة) أو أكثر من حيث الطول، لكن معظم الأنواع كانوا أقل من 20 سنتيمتر (8 بوصة). وتُعد هذه الأنواع مفترسة (predator) بصورة مرعبة حيث تعيش في المياه الضحلة الدافئة في كلٍ من البحار والبحيرات،[3] في العصر الأوردفيشي إلى العصر البرمي منذ 460 إلى 248 مليون سنة مضت2. وعلى الرغم من أنه يُطلق عليها "عقارب البحر" بشكل غير رسمي، فالأوائل من هذه العقارب كانت بحرية (أما اللاحقة فتعيش في المياه المالحة أو المياه العذبة) ولم تكن هذه الأنواع عقارب (scorpion) حقيقية. وطبقًا للنظرية، فإن الانتقال من البحر إلى المياه العذبة يحدث غالبًا من قِبل الحقبة الفرعية البنسلفانية (Pennsylvanian). ويُعتقد أن اليوربتيريدات تمر بمرحلة الانسلاخ (ecdysis)، جاعلة بذلك صعوبة تقييم أهميتها في الأنظمة الاقتصادية، حيث يصعب معرفة الحفرية المنسلخة من هيكل الحفرية الحقيقي.[4] ولقد أصبحت اليوريبتيريدات عملاقة خلال حالة الانقراض التي حدثت في العصر البرمي الترياسي، ونجد أن حفرياتهم موزعة في جميع أنحاء العالم تقريبًا.
وهناك حوالي اثنتي عشرة عائلة من اليوريبتيريدات المعروفة. ومن المحتمل أن الجنس الأكثر انتشارًا من اليوريبتيريدات هو اليوريبتروس (Eurypterus)، والذي منه هناك حوالي 16 حفرية معروفة. وقد وُصف الجنس يوريبتروس (Eurypterus) عام 1825 بواسطة عالم الحيوان جيمس دي كاي ألسويرث (ecdysis)،. فقد تعرّف على طبيعة مفصليات الأرجل في أول عينة يوريبتريد توصف على الإطلاق، والتي اكتشفها الدكتور إس إل ميتشل. وفي عام 1984، سُميت، هذه الأنواع،يوريبتيروس ريبمبز (Eurypterus remipes) حفريات الدولة في نيويورك.
تركيب الجسم
نموذج

وقد وُصفت اليوريبتيريدات بشكلٍ رسمي كالآتي:[5]
"هي عبارة عن ميروستومس (merostomes) ذات شكل سناني طويل، وجسد ثلاثي الفصوص على نحوٍ نادر؛ حيث تتكون من بروسوما [الرأس] ذات حجم متوسط وأوبيثوسما [جسم] مع 12 قطعة متحركة ودُبير أبري الشكل إلى نهاية مستديرة ويد ضيقة (يشبه الملعقة) [ذيل]، مع تقاسيم مشتركة إلى ما قبل الجسم 7 قطع وما بعد الجسم 5 قطع؛ و6 لواحق prosomal [رأسية]، مكونة بذلك 3 تأشيرات مشتركة، وأرجل للسير وعادةً يتحوّل آخر زوج إلى أرجل سباحة. ولها فم في المنتصف، يتوّسع بعد ذلك عن طريق فتحة داخلية وفتحة تالية. وبها أيضًا غطاء واقٍ ذو لاحقة رقيقة متوسطة ولواحق جوفية مفلطحة الشكل ذات خياشيم غير مُصفحة. من العصر البرمي الأوردوفيكي."
قد كان اليوريبتيريد المثالي ذو درع قرني (carapace)، نصف مستدير كبير ومسطح، يليه جزء مُلحق، وأخيرًا ذيل مرن مستدق الطرف، وينتهي في الغالب بعمود فقري طويل في نهايته (الحشرات المجنحة (Pterygotus) على الرغم من أن لها ذيل طويل إلا أنها ذات عمود فقري أصغر). ونجد خلف رأس اليوريبتريدات أن هناك 12 شريحة في جسمها. وتُشكل هذه الشرائح بواسطة اللوحة الظهرية المسماة tergite، واللوحة الباطنية التي تُسمى sternite. ويكون الذيل، والمعروف باسم الدبير في معظم اليوريبتيريدات مثلما في العقارب الحديثة وفي بعض الأنواع ربما يُستخدم لحقن السم، لكن حتى الآن لم يثبُت أن أيًا من هذه اليرويبتيريدات تُعد سامة. وتمتلك معظم اليوريبتيريدات مغاديف تتجه نحو نهاية الدرع القرني وخلفه، والتي تُستخدم لدفع نفسها خلال تواجدها في المياه. وتمتلك رتيبة الستايلونورينا أرجل تسير بها بدلاً من المغاديف. ويتناقش البعض حول أن هذه المغاديف تُستخدم أيضًا في الحفر. ومن المحتمل أنها تُستخدم لكل من الغرضين. وفي الأسفل، بالإضافة إلى وجود زوج اللواحق التي تُستخدم في السباحة، يمتلك هذا المخلوق 4 أزواج من الأرجل المشتركة للسير، ومخلبين في الأمام، تأشيرات (chelicerae)، وهي تتوسع في البتيريجوتيدات (الجناحيات pterygotids). يكون على أرجل السير شعر غريب، وهو يشابه السلطعون الحديث. وهناك صفات أخرى مشتركة بين مفصليات الأرجل القديمة والحديثة من هذا النوع، متضمنة زوجًا واحدًا من العيون المجمعة (compound eye) وزوج من العيون الأصغر والتي تُسمى عيينة (ocelli)، وهي تكون بين الزوج الأكبر الآخر من العيون.

(Carboniferous) المُتأخر.
ونجد أن العديد من اليوريبتيردات كانت لديها أرجل كبيرة وطويلة كافية للقيام بما هو أكثر من الزحف في قاع البحر؛ حيث هناك عدد من الأنواع (وخاصةً هيبرتوبتروس [hibbertopterids]) التي كان لديها أرجل كبيرة قوية البنية ومن المحتمل أنها كانت قادرة على التنقل في البرية (مثل السلطعون الأرضي الموجود حاليًا). وهناك دراسات حول ما يُعتقد أنها آثار تخص تلك الأنواع والتي تُشير إلى أن اليوريبتيريدات في مرحلة ما كانت كائن سداسي الأرجل (ستة أرجل) وكائن أخطبوطي (ذو ثمان أرجل). وقد تكون بعض الأنواع برمائية، والتي تظهر على الأرض على الأقل كجزء من دورة حياتها؛ حيث من الممكن أنها تكون قادرة على التنفس في كلٍ من الماء والهواء.[6] وإن مفصليات الأرجل الضارية والتي تُعرف آثارها باسم بروتكنايتس(Protichnites)،[7] قد وجدت في طبقات العصر الكمبري والتي تعود إلى 510، فمن المحتمل أن تكون مجموعة جذعية (stem group) لليوريبتيريد، وهي من ضمن الدليل الأول لوجود الحيوانات على الأرض في هذا العصر.[8]
ومن ضمن اليوريبتيريدات الهيبرتوبترودس (hibbertopteroids)، والتي سُميت باسم أخصائي علم الأحافير البريطاني دكتور صامويل هيبرت والذي وصف الهيبرتوبترودس (Scouleri) على محجر كريكتون الشرقي للحجر الجيري في اسكتلندا عام 1836. وقد تم التعرّف على آثار حفريات (شكل من أشكال الأثر الحفري مؤخرًا في لوثيان الشرقية، اسكتلندا حيث قام بها الهيبرتوبترودس (hibbertopteroids) طوله 1.6  متر.[9] وكان الجنس الأوسع من حيث الوصف لعقارب البحر هو بتيرجوتوس (Pterygotus)، وهو من شعبة مفصليات الأرجل بحجم التمساح. وتكون حفريات بتيرجوتوس (Pterygotus) شائعة نسبيًا على الرغم من ندرة الحفريات المكتملة. وعند الطول 2.1 متر (6 قدم 11 بوصة)، كانت أكبر مفصليات الأرجل المعروفة أنها قد عاشت على الإطلاق حتى وقتٍ حديث. وقد وجدت حفرياتها في جميع أنحاء العالم، فيما عدا في القارة القطبية الجنوبية (Antarctica). وفي عام 2007، وُجد مخلب طوله 46 سـم (18 بوصة) يخص جاكيلوبتروس رحنانيه (Jaekelopterus rhenaniae) (وهي أنواع وُصفت أساسًا عام 1914) بواسطة مجموعة من أخصائيي علم الأحافير يقودهم سايمون برادي (Simon Braddy) في جامعة بريستول، مشيرًا إلى أن جي رحنانيه (J. rhenaniae) كان يبلغ طوله 2.5 م (8 قدم 2 بوصة)، جاعلاً إياه أكبر كائن من مفصليات الأرجل وُجد على الإطلاق.[10][11]
علاقاتهم مع المجموعات الأخرى
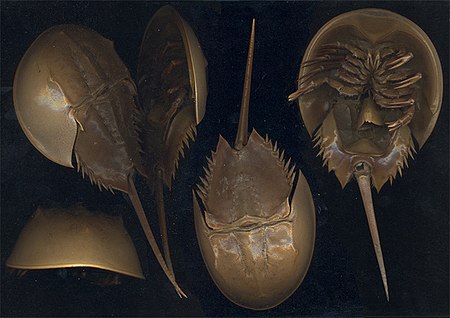
حيث كان يُنظر إلى اليوريبتيريدات عادةً على أنها أقارب السلطعون البحري، الذي على شكل حدوة حصان مُشكلين معًا بذلك مجموعة مسماة ذوات الفتحات المتقطعة (Merostomata). وقد وضعت دراسات لاحقة اليوريبتيريدات أقرب للعنكبوتيات (arachnid) في مجموعة تُسمى ذوات الفتحات التالية (Metastomata).[12] ولقد كان هناك أيضًا فكرة شائعة حول أن اليوريبتيريدات تتصل عن قرب بالعقارب (scorpion)، التي تشبهها.[13] وينعكس هذا الافتراض في الاسم الشائع "عقارب البحر". وقد تم الإدراك مؤخرًا أن هناك مجموعة ضخمة، ولكنها معروفة قليلاً، تُسمى كاسماتاسبيدا (Chasmataspida من كلابيات القرون) تتشارك أيضًا في صفات مع اليوريبتيريدات،[14] وكان يحدث أحيانًا التباسًا في المجموعتين. وقد عرّف أحدث موجز عن العلاقات بين مفصليات الأرجل وأقاربهم أن اليوريبتيريدات وسيفيات الذيل والعنكبيات كثلاث مجموعات رئيسية، لكن لم يستطع الموجز أن يحلل أية تفاصيل مشتركة بينهم.[15]
قائمة الفصائل والأجناس
- طالع أيضًا: List of eurypterids

(Silurian)



هناك 246 نوعًا صالحًا من اليوريبتيريدات بحسب إحصائيات 2011. جميعها منقرضة. ونجدهم مجمعين في الآتي:
- رُتيبة ستايلونيورينا (Stylonurina) دينير، 1924
-
- فيلق رينوبترويديا (Rhenopteroidea) سترومر، 1951
- رينوبترويديا (Rhenopteroidea) سترومر، 1951
- آلكينوبتروس (Alkenopterus) سترومر، 1974
- برايتشيوبتريلا (Brachyopterella) كجيلسفيج-ويرينج، 1966
- برايتشيوبتروس (Brachyopterus) سترومر، 1951
- كياروبتروس (Kiaeropterus) وترستون، 1979
- ليوبتريلا (Leiopterella) لامسدل وبرادي ولوفلر وداينالي، 2010
- رينوبتروس (Rhenopterus) سترومر، 1936
- فيلق ستايلونورويديه (Stylonuroidea) كجيلسفيج-ويرينج، 1959
- باراستايلونوريديه (Parastylonuridae) وترستون، 1979
- باراستايلونوروس (Parastylonurus) كجيلسفيج-ويرينج، 1966
- ستايلونوريلا (Stylonurella) كجيلسفيج-ويرينج، 1966
- ستايلونيوريديه (Stylonuridae)داينر (Diener)، 1924
- سينوبرتروس (Ctenopterus) كلارك ورودنمان، 1912
- لوري إيبتروس (Laurieipterus) كجيلسفيج-ويرينج، 1966
- بيجا (Pagea) وترستون، 1962
- ستايلونوروس (Stylonurus) بايج، 1856
- فيلق كوكومبتيرويديه (Kokomopteroidea) كجيلسفيج-ويرينج، 1966
- كوكومبتيرويديه (Kokomopteroidea) كجيلسفيج-ويرينج، 1966
- كوكومبتيروس (Kokomopterus) كجيلسفيج-ويرينج، 1966
- لامونتوبرتوس (Lamontopterus) وترستون، 1979
- هارديوبتريديه (Hardieopteridae) تولرتون (Tollerton)، 1989
- هوليبيتورس (Hallipterus) كجيلسفيج-ويرينج، 1963
- تارسوبتيريلا (Tarsopterella) سترومر، 1951
- فيلق هيبرتوبتيرويديه (Hibbertopteroidea) كجيلسفيج-ويرينج، 1959
- دريبانوبتيريديه (Drepanopteridae) كجيلسفيج-ويرينج، 1966
- دريبانوبتيروس (Drepanopterus) لوري، 1892
- هيبرتوبتيريديه (Hibbertopteridae) كجيلسفيج-ويرينج، 1959
- كامبيلوسفلوس (Campylocephalus) آيكوولد، 1860
- سيرتوكتينوس (Cyrtoctenus) سترومر، 1968
- دانسبوتروس (Dunsopterus) وترستون، 1968
- هاستيمايما (Hastimima) وايت، 1908
- هيبرتوبتيروس (Hibbertopterus) كجيلسفيج-ويرينج، 1959
- فيرنونوبيتروس (Vernonopterus) وترستون، 1957
- ميكتيروبتيديه (Mycteroptidae) كووب، 1886
- ماجاراكن (Megarachne) هانكيين، 1980
- ميكتيروبس (Mycterops) كووب، 1886
- وودواردوبتيروس (Woodwardopterus) كجيلسفيج-ويرينج، 1959
- ستايلونوريدس (Stylonuroides) كجيلسفيج-ويرينج، 1966
؛رُتيبةاليوريبترينا (Eurypterina) بورميستر، 1843
- Plesion وحدة تصنيفية
- أونيكوبتيريلا (Onychopterella) سترومر، 1951
- فيلق موزلوبتيريديه (Moselopteroidea) لامسدل وبرادي وتيتلي، 2010
- موزلوبتيريديه (Moselopteroidea) لامسدل وبرادي وتيتلي، 2010
- موزلوبتيورس (Moselopterus) سترومر، 1974
- فينتوبتيروس (Vinetopterus) بوشمان وتيتلي، 2004
- فيلق ميجالوجرابوتويديه (Megalograptoidea) كجيلسفيج-ويرينج، 1955
- دميجالوجرابوتويديه (Megalograptoidea) كجيلسفيج-ويرينج، 1955
- آكينوجناثيوس (Echinognathus) والكوت، 1882
- ميجالوجرابتوس (Megalograptus) ميلر، 1874
- فيلق يوريبترويديا (Eurypteroidea) بورميستر، 1843
- دولكوبيتريديه (Dolichopteridae) كجيلسفيج-ويرينج & ستورمر، 1952
- دولكوبيتروس (Dolichopterus) هول، 1859
- رودينمانبتيروس (Ruedemannipterus) كجيلسفيج-ويرينج، 1966
- بافلوبتيروس (Buffalopterus) كجيلسفيج-ويرينج & هيوبتش، 1962
- ستروبيلوبتيروس (Strobilopterus) رودمان، 1935
- سينتوموبتيروس (Syntomopterus) كجيلسفيج-ويرينج، 1961
- الإيوريبترينا (Eurypterina) بورميستر، 1843
- إيوريبتيروس (Eurypterus) دي كاي، 1825
- إيريوبتريديه (Erieopteridae) تولرتون (Tollerton)، 1989
- إيريوبتريوس (Erieopterus) كجيلسفيج-ويرينج، 1958
- فيلق ميكسوبتيرويديه (Mixopteroidea) كجيلسفيج-ويرينج، 1955
- السلطعونيات (Carcinosomatidae) سترومر، 1934
- كارسينوسوما (Carcinosoma) كلاي بول, 1890
- إوركاسينوسوما (Eocarcinosoma) كجيلسفيج-ويرينج، 1964
- باراكارسينوسوما (Paracarcinosoma) كجيلسفيج-ويرينج & كاستر، 1964
- رينوكارسينوسوما (Rhinocarcinosoma) نوفوجيلوف، 1962
- ميكروبتيريديه (Micopteridae) كجيلسفيج-ويرينج & كاستر، 1955
- لاناركبتيروس (Lanarkopterus) ريتشي (Ritchie)، 1968
- ميكسوبتيروس (Mixopterus) رودمان، 1921
- فيلق ويرينجوبتيرديه (Waeringopteroidea)
- ويرينجوبتيرديه (Waeringopteroidea) (لم يتم نشرها بشكل رسمي)
- جروسبيتروس (Grossopterus) سترومر، 1934
- أوركانبتيروس (Orcanopterus) ستوت وتيتلي وبرادي وناولان وجلاسر وديفييرو، 2005
- ويرينجوبتيروس (Waeringopterus) ليوتز، 1961
- فيلق أديلوفثالمويديه (Adelophthalmoidea) بورماستر، 1989
- آديلوفثالمويديه (Adelophthalmoidea) توليرتون، 1989
- آديلوفثالمويديه (Adelophthalmoidea) جوردانفي جوردان وفون ماير، 1854
- باسيبتيروس (Bassipterus) كجيلسفيج-ويرينج ولوتز، 1966
- أيسيلوبتيروس (Eysyslopterus) بوتشمان وتيتلي، 2008
- نانوهجميلاريا (Nanahughmilleria) كجيلسفيج-ويرينج، 1961
- باراهوجهميليريا (Parahughmilleria) كجيلسفيج-ويرينج، 1961
- بيتسفورديبتيروس (Pittsfordipterus) كجيلسفيج-ويرينج ولوتز، 1966
- فيلق بتيرجوتيوديه (Pterygotioidea) كلارك ورودنمان، 1912
- هجميليريديه (Hughmilleriidae) كجيلسفيج-ويرينج، 1951
- هيرفوردبتيروس (Herefordopterus) تيتلي، 2006
- هوجهميليريا (Hughmilleria) سارل، 1902
- سيليمونيديه (Slimonidae) نوفوجيلوف، 1968
- سليمونيا (Slimonia) بايج، 1856
- سالتيروبتيروس (Salteropterus) كجيلسفيج-ويرينج، 1951
- بتيريجوتيديه (Pterygotidae) كلارك ورودنمان، 1912
- بتيريجوتوس (Pterygotus) أغاسيز، 1839
- أكيوتيرموس (Acutiramus) رودمان، 1935
- سيوركوبتيروس (Ciurcopterus) بريجز وتيتلي، 2009
- إيريتوبتيروس (Erettopterus) سالترفي هكسلي وسالتر، 1859
- جايكيلوبتيروس (Jaekelopterus) وترستون، 1964
- كلاركيبتيروس (Clarkiepterus) كجيلسفيج-ويرينج، 1966
- دورفوبتيروس (Dorfopterus) كجيلسفيج-ويرينج، 1955
- هولميبتيروس (Holmipterus) كجيلسفيج-ويرينج، 1979
- مارسوبيبتيروس (Marsupipterus) كجيلسفيج-ويرينج & كاستر(Kjellesvig-Waering&Caster)، 1955
- برايتشيوبتروس(Brachyopterus)سترومر، 1951
- يونيونوبتيروس (Unionopterus) تشيرنيشيف، 1948
نجد أن آثار أثر حفريات بواسطة اليوربتيريدات المنقرضة موضوعة في بصمات الحفريات البالميتشنيوم(Palmichnium).[16]
تاريخ تطور السلالة

إن المخطط التوضيحي المقدم هنا هو مبسط عن دراسة قام بها تيتلي.[17] ويقوم أهم توقف في الخاص بتطور السلالات على أساس إبداعين رئيسيين اللذان ميزا تطور اليوريبتيريدات. كان أهمهم تحوُّل اللاحقة الجسدية الأولية الأكثر تأخرًا إلى مغاديف للسباحة (كما وُجد في اليوريبتيرينا كليد [clade Eurypterina]). والإبداع الثاني هو توّسُع كلابيات القرون، (كما وُجد في شعبة بتيريجوتيديه)، والسماح باستخدام تلك المغاديف في الإمساك الحيوي بالفريسة.
ونجد أنه حوالي 75% من الأنواع هي يوربترينات؛ وهو ما يُمثل 99% من النماذج.[1] ويُعتبر الفيلق بتيريجويتدواديه (Pterygotioidea) هو أكثر فرع حيوي في الأنواع ثراءً، يليه أديلوفثالوميديه(Adelophthalmoidea) ذو 43 نوعًا، مثل الأصناف الشقيقة، فهما يكونان اليوريبتيريدات الأكثر اشتقاقًا. وتتضمن بتيريجويتدواديه(Pterygotioidea) البتيريجوتيدات، والتي تعتبر اليوربتيريدات الوحيدة التي تمتلك توزيع كوزموبوليتاني (عالمي).[17] ويُعد هذا الفرع الحيوي في التصنيف هو أحد أفضل الدعائم خلال اليوربتيريدات.
وقد اقتُرح أن تطور الدرع الجلدي (dermal armour) في بعض مجموعات الفقاريات عديمة الفك (jawless vertebrate) (مثل، هيتيروستراسي (Heterostraci) وأوستيوستراسي (Osteostraci)) هو استجابة لضغط الضراوة (predation) من قبل كائنات اليوربتيريدات المتطورة الضارية بنحو متزايد[18] (خاصةً البتيريجوتيدات pterygotid) على الرغم من أنه لم يتم التحقق من ذلك بعد بالتحليل المفصّل. ويرتبط التزايد في تنوع الأسماك بالانخفاض في تنوع اليوربتيريدات في العصر الديفوني الأدنى (Lower Devonian)،[19] على الرغم من أنه لا يُعتقد بأن ذلك يمثل إحلالاً تنافسيًا، ففي الواقع، يُعد ذلك ندرة في السجلات الحفرية (fossil record).[20]
| Eurypterida | |
- The position of the megalograptids is uncertain.
مقالات ذات صلة
المراجع
- Dunlop, J. A. (2008). "How many species of fossil arachnids are there?". Journal of Arachnology. 36 (2): 267–272. doi:10.1636/CH07-89.1. ISSN 0161-8202.
- Webster's New Universal Unabridged Dictionary. 2nd ed. 1979.
- Joseph T. Hannibal, Spencer G. Lucas, Allan J. Lerner & Dan S. Chaney (2005). S. G. Lucas, K. E. Zeigler & J. A. Spielmann (المحرر). "The Permian of Central New Mexico" ( كتاب إلكتروني PDF ). New Mexico Museum of Natural History and Science Bulletin. 31: 34–38. مؤرشف من الأصل في 04 مارس 2016. نسخة محفوظة 4 مارس 2016 على موقع واي باك مشين.
- Brandt, Danita (2009). "Scorpion taphonomy: criteria for distinguishing fossil scorpion molts and carcasses". Journal of Arachnology. 37.3: 312+.
- L. Størmer (1955). "Merostomata". Treatise on Invertebrate Paleontology, Part P Arthropoda 2, Chelicerata. صفحة 23.
- "Gigantic scorpion' fossil found in Fife". بي بي سي نيوز. April 24, 2010. مؤرشف من الأصل في 10 ديسمبر 2019.
- Richard Owen (1852). "Description of the impressions and footprints of the Protichnites from the Potsdam Sandstone of Canada". Quarterly Journal of the Geological Society of London. 8: 214–225. doi:10.1144/GSL.JGS.1852.008.01-02.26.
- Lancaster D. Burling (1917). "Protichnites and Climactichnites. A critical study of some Cambrian trails". American Journal of Science. 44 (263): 387–398. doi:10.2475/ajs.s4-44.263.387.
- Whyte, Martin A. (2005). "Palaeoecology: A gigantic fossil arthropod trackway". [[نيتشر (مجلة)|]]. 438 (7068): 576–576. doi:10.1038/438576a. PMID 16319874.
- Simon J. Braddy, Markus Poschmann & O. Erik Tetlie (2008). "Giant claw reveals the largest ever arthropod". رسائل الأحياء. 4 (1): 106–109. doi:10.1098/rsbl.2007.0491. PMC . PMID 18029297.
- "Giant claw points to monster sea scorpion". نيو ساينتست. November 21, 2007. مؤرشف من الأصل في 08 مايو 2015.
- P. Weygoldt & H. F. Paulus (1979). "Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata". Zeitschrift für zoologische Systematik und Evolutionsforschung. 17 (2): 85–116, 177–200. doi:10.1111/j.1439-0469.1979.tb00694.x.
- J. Versluys & R. Demoll (1920). "Die Verwandtschaft der Merostomata mit den Arachnida und den anderen Abteilungen der Arthropoda". Koninklijke Akademie van Wetenschappen Amsterdam. 23: 739–765.
- O. Erik Tetlie & Simon J. Braddy (2004). "The first Silurian chasmataspid, Loganamaraspis dunlopi gen. et sp. nov. (Chelicerata: Chasmataspidida) from Lesmahagow, Scotland, and its implications for eurypterid phylogeny". الجمعية الملكية في إدنبرة: Earth Sciences. 94 (3): 227–234. doi:10.1017/S0263593300000638.
- Jeffrey W. Shultz (2007). "A phylogenetic analysis of the arachnid orders based on morphological characters". Zoological Journal of the Linnean Society. 150 (2): 221–265. doi:10.1111/j.1096-3642.2007.00284.x. CiteSeerX: 10
.1 .1 .119 .850. - Adolf Seilacher (2007). "Arthropod trackways". Trace Fossil Analysis. Springer. صفحات 18–31. . مؤرشف من الأصل في 10 ديسمبر 2019.
- O. Erik Tetlie (2007). "Distribution and dispersal history of Eurypterida (Chelicerata)" ( كتاب إلكتروني PDF ). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. مؤرشف من الأصل في 14 أغسطس 2012. نسخة محفوظة 14 أغسطس 2012 على موقع واي باك مشين.
- Romer, A. S. (1933). "Eurypterid influence on vertebrate history". ساينس. 78 (2015): 114–117. doi:10.1126/science.78.2015.114. PMID 17749819.
- D. E. G. Briggs (1988). "Extinction and the fossil record of arthropods". Clarendon press.: 171–209.
- Ridley, Mark (2004). Evolution. Wiley-Blackwell. .
كتابات أخرى
- Simon J. Braddy (2001). "Eurypterid Palaeoecology: palaeobiological, ichnological and comparative evidence for a 'mass-moult-mate' hypothesis". Palaeogeography, Palaeoclimatology, Palaeoecology. 172 (1–2): 115–132. doi:10.1016/S0031-0182(01)00274-7.
- Samuel J. Ciurca (1998). "The Silurian Eurypterid Fauna". مؤرشف من الأصل في 21 مايو 201925 يوليو 2004.
- John M. Clarke & R. Rudolf (1912). The Eurypterida of New York. Albany, New York: New York State Education Department.
- Neal S. Gupta, O. Erik Tetlie, Derek E. G. Briggs & Richard D. Pancost (2007). "The fossilization of eurypterids: a result of molecular transformation" ( كتاب إلكتروني PDF ). Palaios. 22 (4): 439–447. doi:10.2110/palo.2006.p06-057r. مؤرشف من الأصل في 28 مارس 2020.
- Phillip L. Manning & Jason A. Dunlop (1995). "The respiratory organs of eurypterids" ( كتاب إلكتروني PDF ). Palaeontology. 38 (2): 287–297. مؤرشف من الأصل في 14 أغسطس 2012.
- O. Erik Tetlie (2007). "Distribution and dispersal history of Eurypterida (Chelicerata)" ( كتاب إلكتروني PDF ). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. مؤرشف من الأصل في 14 أغسطس 2012.
- O. Erik Tetlie & Michael B. Cuggy (2007). "Phylogeny of the basal swimming eurypterids (Chelicerata; Eurypterida; Eurypterina)". Journal of Systematic Palaeontology. 5 (3): 345–356. doi:10.1017/S1477201907002131.
وصلات خارجية
- Eurypterids.co.uk – eurypterid information website
- Life-like reconstruction of a eurypterid
- Eurypterida from the Palaeos website
- Jaekelopterus rhenaniae at the Palaeoblog