
تعتبر الهياكل البلورية لجزيئات البروتين والحمض النووي والمجمعات الخاصة بها أساسية لممارسة معظم أجزاء علم الأحياء-فيزيائي، وشكل الكثير مما نفهمه علميا على مستوى التفصيل الذري لعلم الأحياء.وتؤكد الأمم المتحدة أهمية هذه الأنشطة بإعلان عام 2014 سنة دولية للتصوير البلوري، في الذكرى المئوية لجائزة نوبل التي حصلت عليها ماكس فون لاوي عام 1914 لاكتشاف حيود الأشعة السينية بواسطة البلورات. تعتمد هذه القائمة التسلسلية للتراكيب البروتينية والحمض النووي البارزة على المراجعة في المجلة الحيوية الطبيعية . [1] وتشمل القائمة جميع الهياكل الاثنى عشر المتميزة، تلك التي فتحت آفاقا جديدة في الموضوع أو الطريقة، وتلك التي أصبحت نظما نموذجية للعمل في مجالات البحوث البيولوجية - الفيزيائية في المستقبل.
الميوجلوبين


1960 - كان الميوغلوبين أول هيكل بلوري عالي الدقة لجزيء البروتين. [2] تخمد العضلة العضلية مجموعة الهيمي التي تحتوي على الحديد والتي تربط بشكل عجيبي الأكسجين لاستخدامه في تزويد ألياف العضلات بالطاقة، وكانت تلك البلورات الأولى من الميوغلوبين من حوت الحيوانات المنوية ، الذي تحتاج عضلاته إلى تخزين كبير من الأوكسجين للغوصات العميقة. يتكون هيكل الميوجلوبين ثلاثي الأبعاد من 8 حلزونات ألفا ، وأظهرت البنية البلورية أن التشكل كان باليد اليمنى وتطابق بشكل وثيق الهندسة التي اقترحها لينوس بولينج، مع 3.6 بقايا لكل لفة وروابط هيدروجين رئيسية من بيبتيد NH لبقايا واحدة إلى بيبتيد CO من المخلفات i+4. الميوجلوبين هو نظام نموذجي للعديد من أنواع الدراسات الفيزيائية الحيوية، [3] ويشمل ذلك بصفة خاصة عملية للأجناس الصغيرة مثل الأكسجين وأول أكسيد الكربون .
الهيموغلوبين
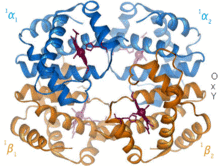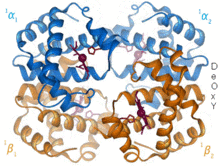
1960 - أظهر هيكل بلورات الهيموغلوبين [4] أظهر رباعي الأوجه (Tetramer) من نوعين من السلاسل المرتبطة وتم حله بدقة أقل بكثير من مونوميريك ميغلوبين، لكن بوضوح كان له نفس بنية اللولب الثمانية الأساسية (والتي تسمى الآن "جلوبين فلايت"). المزيد من الهيموجلوبين هياكل البلور في دقة أعلى [PDB 1MHB, 1Dhb) قريبا أظهرت التغير المتقارن بين كل من كونيتيكشن المحلي والكواترنائي بين حالات أوكسي وديوكسي من الهيموجلوبين، [5] مما يفسر تعاون رابط الأكسجين في الدم وتأثير اللوستريك عوامل مثل الرقم الهيدروجيني و DPG . لعقود من الزمن كان الهيموغلوبين مثال التدريس الأساسي لمفهوم الخلاصة، بالإضافة إلى كونه تركيزًا مكثفًا للبحث والمناقشة حول التباين. في عام 1909 ، استخدمت بلورات الهيموغلوبين من أكثر من 100 نوع لربط التصنيف بالخصائص الجزيئية. [6] وقد استشهد بيروتز بهذا الكتاب في تقرير عام 1938 [7] عن بلورات الهيموغلوبين في الخيول التي بدأت ملحمته الطويلة لحل التركيب البلوري. بلورات الهيموغلوبين هي متعدد التلاون - الأحمر الداكن في اتجاهين والأحمر الباهت في الثلث [6] - بسبب توجه hemes ، وفرقة مشرق من الهيمي مجموعات البورفيرين تستخدم في التحليل الطيفي للهيموجلوبين ربط..

الليزوزيم الابيض- البيض-الدجاج
1965 - الليزوزيم الابيض- البيض-الدجاج (ملف PDB 1lyz). [8] الدهن البيض البيض الليزوزيم (ملف PDB 1lyz). كان التركيب البلوري الأول لإنزيم (يشبك الكربوهيدرات الصغيرة في سكريات بسيطة) ، ويستخدم للدراسات المبكرة لآلية الإنزيم[9] كانت تحتوي على ورقة بيتا (مضادة للأضداد) بالإضافة إلى حلزونات، وكانت أيضًا أول بنية جزيئية كبيرة تمت صقل إحداثياتها الذرية (في الفضاء الحقيقي). [10] [11]يمكن شراء مادة البداية الخاصة بالتحضير من متجر البقالة، ويتبلور الليسوزيمي بالبيض بشكل سريع جدا في العديد من مجموعات الفضاء المختلفة; وهي حالة الاختبار المفضلة للتجارب والأدوات البلورية الجديدة. ومن الأمثلة الحديثة على ذلك النانو-البلورات في اللايزومي لجمع بيانات الليزر الحرة الإلكترون
وبلورات دقيقة لحيود الإلكترون الصغير.. [12]

Ribonuclease
1967 - ريبونوكلياز A (ملف PDB 2RSA) [13] هو إنزيم ينقذ الحمض النووي الريبوزي (RNA) يتم تثبيته بواسطة 4 روابط غير حقيقية. وقد تم استخدامه في الأبحاث الأساسية التي أجراها أنفينسن حول طي البروتين والتي أدت إلى مفهوم أن البنية ثلاثية الأبعاد للبروتين قد تم تحديدها بتسلسل الحمض الأميني. Ribonuclease S ، كان الشكل المشقوق المكون من عنصرين، الذي درسه فريد ريتشاردز، نشطًا أيضًا من الناحية الأنزيمية، وكان له بنية بلورية متطابقة تقريبًا (ملف PDR 1RNS) [14] وقد ظهر أنه نشط حافزًا حتى في البلورة، [15] المساعدة في تبديد الشكوك حول أهمية التركيبات البلورية للبروتين للوظيفة البيولوجية.

سيرينproteases
1967 - serine proteases هي مجموعة هامة جدا تاريخيا من هياكل الإنزيمات، لأنها معا أضاءت الآلية الحفازة (في حالتها، من قبل سيره-ASP "Triad الحفازة" ، أساس الاختلاف في الخصائص الأساسية للركيزة وآلية التنشيط التي من خلالها يقوم الانفصال الأنزيمي الخاضع للتحكم بحفر نهاية السلسلة الجديدة لإعادة ترتيب الموقع النشط بشكل صحيح [16] وتضمنت المباني وضوح الشمس في وقت مبكر كيموتربسين (ملف PDB 2CHA)، [17] مولد الكيموتربسين (ملف PDB 1CHG)، [18] التربسين (1PTN ملف PDB)، [19] و الإيلاستاز (ملف PDB 1EST). [20] كانت أيضًا أول هياكل بروتينية أظهرت مجالين شبه متطابقين، يُعتقد أنهما مرتبطان بتكرار الجينات.. أحد أسباب استخدامها على نطاق واسع كأمثلة للكتب المدرسية والفصول الدراسية هو نظام ترقيم كود الإدخال (الذي يكره جميع مبرمجي الكمبيوتر) ، مما جعل Ser195 و His57 ثابتًا ولا يُنسى على الرغم من الاختلافات في تسلسل البروتين
غراء
1968 - غراء


إنزيم البروتياز
1969 - كاربوكسي بيبتيديز A هو ميتالوبروتياز الزنك. أظهر هيكلها البلوري (ملف PDB 1CPA) [21] أظهر أول هيكل بيتا متواز: ورقة مركزية كبيرة ملتوية من 8 خيوط مع Zn موقع نشط في الطرف الطرفي على شكل C من الخيوط الوسطى والصفيحة المحاطة على كلا الجانبين بجدائل ألفا.. وهو exopeptidase الذي يشبك الببتيدات أو البروتينات من الطرف النهائي للكربوكسيل بدلاً من التسلسل الداخلي. في وقت لاحق، تم حل مثبط بروتين صغير من الكربوكسيببتيداز (ملف PDB 4CPA) [22] ويقوم ذلك بإيقاف الحفاز ميكانيكيا عن طريق تقديم الطرف الطرفي C الخاص به والذي يلتصق فقط من بين حلقة من الروابط غير الحقيقية مع بنية ضيقة خلفها، مما يمنع الإنزيم من الامتصاص في السلسلة بعد البقايا الأولى.
بروتياز
Lactate dehydrogenase
1970 - اللاكتات ديهيدروجينيزLactate dehydrogenase
مثبط التربسين
1970 - مثبط التربسين البنكرياسي الأساسي ، أو BPTI ،Trypsin inhibitor
Rubredoxin
1970 - روبريدوكسين


الأنسولين
1971 - الأنسولين (ملف PDIN 1INS) [23] هو هرمون مركزي لعملية التمثيل الغذائي لتخزين السكر والدهون، وهو مهم في الأمراض التي تصيب الإنسان مثل السمنة والسكري .ومن الناحية البيولوجية، يلاحظ وجود ترابط Zn، وتوازنه بين حالات الأمونومر، والديمر، والكسر، وقدرته على تكوين البلورات في الجسم الحي، وتركيبه كشكل "مناصر" أطول، ثم يتم تقسيمه إلى قسمين نشطين مرتبطين بالسلسلة SS . لقد حقق الأنسولين نجاحًا في برنامج النمو البلوري التابع لوكالة ناسا على متن مكوك الفضاء ، حيث أنتج مستحضرًا ضخمًا من بلورات صغيرة جدًا موحدة للجرعة التي يتم التحكم فيها.
المكورات العنقودية النووية
1971 - المكورات العنقودية النووية
السيتوكروم
1971 - Cytochrome C
T4 فج الليزوزيم
1974 - T4 فج الليزوزيم
المناعية
1974 - الغلوبولين المناعي
ديسموتاز سوبيروكسيد
1975 - Cu، Zn Superoxide disutaseSuperoxide dismutase
نقل الحمض النووي الريبي
1976 - نقل الحمض النووي الريبي

ثلاثي الفوسفات إيزوميراز
1976 - ثلاثي الفوسفات إيزوميراز
بيبسين مثل البروتياز الأسبارتيك
1976 - ريزوبوسبسين
1976 - Endothiapepsin
1976 - البنسلوبيسين
فيروسات الايكوساهدرا
1978 - فيروس إيكوساهدرا
1981 - ديكرسون B-شكل DNA dodecamer
1981 - كرامبين
1985 - الكالمودولين
1985 - بوليميريز الحمض النووي
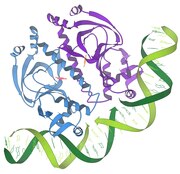
تفاعلات المثبط / الحمض النووي
1986 - تفاعلات المثبط / الحمض النووي
1987 مجمع التوافق النسيجي الرئيسي
1987 يوبيكيتين
1987 ROP البروتين
فيروس نقص المناعة البشرية - 1 البروتيني الأسبارتيك
1989 فيروس نقص المناعة البشرية -1 البروتيني
1990 باكتيورودوبسين
1991 لفائف GCN4 ملفوف
فيروس نقص المناعة البشرية -1 عكس نسخة
1991 فيروس نقص المناعة البشرية -1 نسخة عكسية
1993 Beta helix of Pectate lyase
1994 الكولاجين
1994 Barnase / مجمع barstar
1994 F1 ATPase
1995 البروتينات غير المتجانسة G
1996 CDK / cyclin complex

كينيسين
1996 - البروتين الحركي Kinesin
1997 GroEL / ES chaperone
1997 Nucleosome
1998 المجموعة الأولى إنترون الربط الذاتي

توبويسوميراز الحمض النووي
1998 - يؤدي الحمض النووي توبويزوميراز قم بتنفيذ المهمة الحيوية والضرورية لفك تشابك خيوط DNA أو حلقاتها التي تتشابك مع بعضها البعض أو التواء بإحكام أثناء العمليات الخلوية العادية مثل نسخ المعلومات الجينية.
1998 توبولين ألفا / بيتا ديمر
1998 قناة البوتاسيوم
1998 تقاطع هوليداي
2000 ريبوسوم
2000 AAA + ATPase
2002 يكرر أنكرين
2003 تصميم البروتين TOP7
2004 البروتينات السيانوباكتيرية على مدار الساعة
2004 ريبوسويتش
2006 exosome الإنسان
2007 مستقبلات G- البروتين

قبو الجسيمات
2009 يعد جزيء Vault هو اكتشاف جديد مثير للاهتمام لجزيئات جوفاء كبيرة شائعة في الخلايا، مع العديد من الاقتراحات المختلفة لوظيفتها البيولوجية المحتملة. الهياكل البلورية (PDB ملفات 2Zuo، 2ZV4، 2ZV5
[24] و 4hl8 [25] ) أن كل نصف من قبو يتكون من 39 نسخة من بروتين طويل النطاق يضم 12 مجالًا معًا لتشكيل العلبة. يشير الاضطراب في الأطراف العلوية والسفلية إلى وجود فتحات لإمكانية الوصول إلى الجزء الداخلي من الخزانة.
علم بلوري الليزر الحر
2010 - علم بلورات الليزر الإلكترون الحر
المراجع
- "Biophysical highlights from 54 years of macromolecular crystallography". Biophysical Journal. 106: 510–525. 2014. doi:10.1016/j.bpj.2014.01.001. PMID 24507592.
- "Structure of myoglobin: A three-dimensional Fourier synthesis at 2Å resolution". Nature. 185 (4711): 422–427. 1960. doi:10.1038/185422a0. PMID 18990802.
- "Myoglobin: the hydrogen atom of biology and a paradigm of complexity". Proceedings of the National Academy of Sciences USA. 100: 8615–8617. 2003. doi:10.1073/pnas.1633688100.
- "Structure of haemoglobin: A three-dimensional Fourier synthesis at 5.5-Å resolution, obtained by X-ray analysis". Nature. 185 (4711): 416–422. 1960. doi:10.1038/185416a0. PMID 18990801.
- Perutz MF (1970). "Stereochemistry of Cooperative Effects in Haemoglobin: Haem–Haem Interaction and the Problem of Allostery". Nature. 228 (5273): 726–734. doi:10.1038/228726a0.
- Edward Tyson Reichert & Amos Peaslee Brown (1909). The Differentiation and Specificity of Corresponding Proteins and other Vital Substances in Relation to Biological Classification and Organic Evolution: The Crystallography of Hemoglobins. Washington, DC: Carnegie Institution.
- "X-ray syudy of chymotrypsin and hemoglobin". Nature. 141: 523–524. 1938. doi:10.1038/141523a0.
- "Structure of hen egg-white lysozyme: A three-dimensional Fourier synthesis at 2Å resolution". Nature. 206 (4986): 757–761. 1965. doi:10.1038/206757a0. PMID 5891407.
- "Theoretical studies of enzymic reactions: Dielectric, electrostatic, and steric stabilization of the carbonium ion in the reaction of lysozyme". J Mol Biol. 103: 227–49. 1976. doi:10.1016/0022-2836(76)90311-9. PMID 985660.
- Diamond R (1974). "Real-space refinement of the structure of hen egg-white lysozyme". Journal of Molecular Biology. 82: 371–374. doi:10.1016/0022-2836(74)90598-1.
- "High-resolution protein structure determination by serial femtosecond crystallography" ( كتاب إلكتروني PDF ). Science. 337: 362–364. 2012. doi:10.1126/science.1217737. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 22 سبتمبر 2017.
- "Three-dimensional electron crystallography of protein microcrystals". eLife. 2: e01345–e01345. 2013. doi:10.7554/elife.01345.
- "Tertiary structure of ribonuclease". Nature. 213 (5079): 862–865. 1967. doi:10.1038/213862a0. PMID 6043657.
- "The structure of ribonuclease-S at 3.5 Å resolution". Journal of Biological Chemistry. 242: 3984–3988. 1967.
- "The activity of an enzyme in the crystalline state: Ribonuclease-S". Journal of Biological Chemistry. 238: 2399–2406. 1963.
- Structure and Action of Proteins. New York: Harper. 1969.
- "Three-dimensional structure of tosyl-α-chymotrypsin". Nature. 214 (5089): 652–656. 1967. doi:10.1038/214652a0.
- "Chymotrypsinogen: 2.5-Å crystal structure, comparison with α-chymotrypsin, and implications for zymogen activation". Biochemistry. 9 (9): 1997–2009. 1970. doi:10.1021/bi00811a022. PMID 5442169.
- "The refined crystal structure of bovine beta-trypsin at 1.8Å resolution, II. Crystallographic refinement, calcium binding site and active site at pH 7.0". Journal of Molecular Biology. 98: 693–697. 1975. doi:10.1016/s0022-2836(75)80005-2.
- "The atomic structure of crystalline porcine pancreatic elastase at 2.5Å resolution: comparison with the structure of alpha-chymotrypsin". Journal of Molecular Biology. 118: 137–208. 1978. doi:10.1016/0022-2836(78)90412-6.
- Lipscomb WN; Hartsuck JA; Reeke GN; Quiocho FA; Bethge PH; Ludwif=g ML; Steitz TA; Muirhead H; Coppola JC (1969). "The structure of carboxypeptidase A, VII. The 2.0-Å resolution studies of the enzyme and of its complex with glycyltyrosine, and mechanistic deductions". Brookhaven Symposia in Biology. 21: 24–90.
- "Refined crystal structure of the potato inhibitor complex of carboxypeptidase A at 2.5 A resolution". Journal of Molecular Biology. 160: 475–498. 1982. doi:10.1016/0022-2836(82)90309-6.
- "Atomic positions in rhombohedral 2-zinc insulin crystals". Nature. 231 (5304): 506–511. 1971. doi:10.1038/231506a0. PMID 4932997.
- "The Structure of Rat Liver Vault at 3.5 Angstrom Resolution". Science. 323 (5912): 384–388. 2009. doi:10.1126/science.1164975. PMID 19150846.
- "New features of vault architecture and dynamics revealed by novel refinement using the deformable elastic network approach". Acta Crystallographica. D69 (Pt 6): 1054–1061. 2013. doi:10.1107/S0907444913004472. PMID 23695250.