Chez les espèces utilisant la reproduction sexuée, l'attirance sexuelle (ou attraction sexuelle) désigne l'attirance vers d'autres membres de la même espèce pour réaliser la copulation ou pour des activités érotiques.
Plusieurs hypothèses relatives à la théorie de l'évolution suggèrent que certaines caractéristiques particulières des organismes (les caractères sexuels secondaires : la roue des paons, la dimension des bois des cerfs, la peau sexuelle des primates ; le volume des seins, la jeunesse et le faible rapport taille-hanche des femmes[1],[2], la forte masse musculaire et la faible masse grasse des hommes[3],[4],[5],[6], etc.), qui provoqueraient de l'attirance sexuelle, ont été sélectionnés par l'évolution pour augmenter les chances de reproduction avec le partenaire le plus approprié.
La nature de l'attirance sexuelle ne semble pas être uniquement biologique. Ainsi plusieurs facteurs attractifs populaires, telles que le volume des seins ou le rapport taille-hanche faible de la femme, pourraient correspondre à des préférences sexuelles apprises en raison de caractéristiques culturelles particulières.
NB : cet article présente les théories et les données scientifiques concernant les caractéristiques morphologiques, sonores, olfactives, etc., qui provoquent une attirance sexuelle entre les animaux. Les processus neurobiologiques spécifiques qui induisent une attirance durable selon le sexe du partenaire sexuel sont présentés dans l'article Orientation sexuelle. Les processus neurobiologiques qui sont à l'origine de préférences sexuelles pour certaines caractéristiques des partenaires ou pour quelques partenaires particuliers sont présentés dans l'article Préférence sexuelle.
Cadre général
La synthèse des données actuellement disponibles suggère que la plupart des animaux sont biologiquement organisés pour la copulation hétérosexuelle[7]. En fonction des caractéristiques des circuits neurobiologiques connus, c'est la communication chimique qui apparaît comme la plus adaptée pour transmettre les signaux déclencheurs de l'attraction sexuelle[8].
Circuits neurobiologiques de la copulation
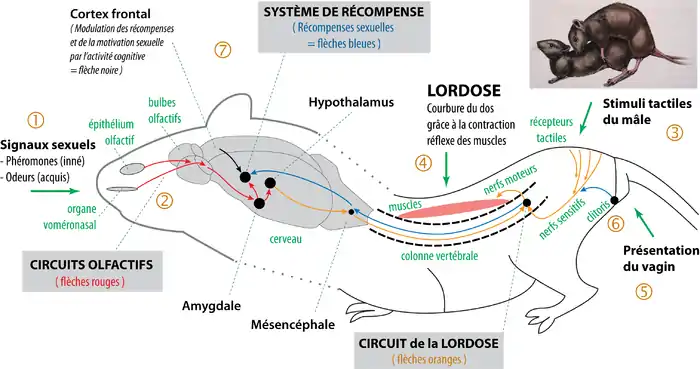
Les études scientifiques récentes montrent que l'organisation neuroanatomique générale des mammifères non-primates est spécifiquement conçue pour la copulation hétérosexuelle[7]. En simplifiant, il existe trois grands circuits neurobiologiques précablés, contrôlés par les hormones :
- Les circuits olfactifs (flèches rouges, schéma ci-contre), à l'origine de l'excitation sexuelle et de l'orientation sexuelle[9] ;
- Les circuits des réflexes sexuels (lordose[10], érection, éjaculation : flèches orange), qui permettent la copulation ;
- Les circuits des récompenses sexuelles (système de récompense associé au pénis/clitoris : flèches bleues), qui sont impliqués dans les apprentissages sexuels[11] (en particulier de la motivation sexuelle)[8].
En fonction de cette organisation, chez les mammifères, ce sont les circuits olfactifs qui apparaissent comme les plus adaptés pour traiter de manière innée les signaux déclencheurs de l'attraction sexuelle.
Organisation des systèmes sensoriels
L'étude des différents systèmes sensoriels suggère que seule la communication chimique est apparemment la plus adaptée pour véhiculer des signaux relatifs à l'attraction sexuelle[12].
En effet, une molécule « chimique » peut avoir des caractéristiques physiques et chimiques spécifiques, et qui sont bien distinctes des milliers d'autres molécules chimiques existant dans l'environnement. La communication chimique existe à tous les niveaux d'un organisme : dans la cellule, entre les cellules, entre les tissus et les organes. De plus, les molécules chimiques peuvent être facilement synthétisées, émises et détectées par un organisme. Chez les mammifères, plusieurs organes sensoriels (langue, ganglion de Grueneberg, organe septal, etc.) peuvent détecter des molécules chimiques. Mais c'est l'épithélium olfactif et surtout l'organe voméronasal qui semblent spécifiquement organisés pour traiter les signaux chimiques déclencheurs de l'attraction sexuelle[13].
Par rapport aux stimulations corporelles, le système somatosensoriel est constitué de plusieurs types de récepteurs (mécanorécepteurs, nocicepteurs, thermorécepteurs, etc.) localisés dans la peau, les viscères, les muscles et les tendons de l'organisme. À la différence des systèmes sensoriels où les récepteurs sont localisés dans des organes sensoriels, les différents récepteurs somatosensoriels sont disséminés dans tout le corps. Ces récepteurs permettent la détection de stimuli nociceptifs, de la température, de l'étirement, de la pression, des vibrations ou du contact[14]. À ce niveau, on remarque qu'aucun de ces récepteurs ne peut être spécifique d'un signal sexuel. Tout contact avec le corps active des récepteurs somatosensoriels, que ce contact soit sexuel ou non. Il n'existe donc pas de signaux somatosensoriels élémentaires spécifiques du comportement de reproduction. Néanmoins, il existe des connexions spécifiques entre le pénis / clitoris et le système de récompense[15],[16], qui permettent de déclencher les réflexes sexuels et d'induire de nombreux apprentissages sexuels[11]. Mais le système somatosensoriel n'apparaît pas organisé pour traiter des signaux déclencheurs d'une attraction sexuelle.
La vision est un canal sensoriel majeur chez les hominidés. Plusieurs théories ont donc supposé que des signaux visuels pourraient servir de signaux spécifiques au déclenchement de l'attraction sexuelle. Mais les caractéristiques fonctionnelles du système visuel ne semblent guère adaptées pour traiter des signaux innés de l'attraction sexuelle. En effet, les photons qui constituent le rayonnement électromagnétique et la lumière sont des particules d'énergie. Ils peuvent être distingués les uns des autres par leur longueur d'onde comprise entre 103 et 10−14 mètre. Les structures sensorielles de la vision sont les récepteurs (trois types de cônes et un type de bâtonnets) des cellules neurosensorielles de la rétine qui détectent les photons dont la longueur d'onde est comprise entre 400 et 700 nm[14]. En fonction de ces caractéristiques, au mieux, à la lumière du jour, il n'est possible que de détecter trois signaux visuels élémentaires. De plus, au niveau anatomique, il ne semble pas exister chez les rongeurs de connexions entre les structures visuelles et les neurones de l'hypothalamus qui contrôlent le système hormonal de la reproduction (neurones à GnRH/LHRH)[17]. Pour ces raisons, il est peu probable qu'il existe un signal visuel élémentaire inné du comportement de reproduction. Néanmoins, il pourrait être possible qu'une information élaborée, extraite du traitement des stimuli visuels élémentaires, puisse servir de signal inné. Ce traitement, bien que peu probable en raison de son inhérente complexité, pourrait avoir lieu dans les parties corticales associatives du système visuel[12].
Par rapport au système auditif, les structures sensorielles de l'audition sont les cellules épithéliales ciliées sensorielles de l'organe spiral de la cochlée, qui détectent les mouvements provoqués par les vibrations moléculaires de l'air ou de l'eau[14]. Les vibrations moléculaires n'ayant pas de caractéristiques sexuelles spécifiques, il ne peut donc apparemment pas exister de signal auditif élémentaire inné déclencheur de l'attraction sexuelle. De plus, au niveau anatomique, il ne semble pas exister chez les rongeurs de connexions entre les structures auditives et les neurones de l'hypothalamus qui contrôlent le système hormonal de la reproduction (neurones à GnRH/LHRH)[17]. Éventuellement, il pourrait être possible qu'une information élaborée, extraite du traitement des stimuli auditifs élémentaires dans les régions corticales associatives, puisse servir de signal inné. Mais des expériences chez les rongeurs indiquent qu'il n'existe vraisemblablement pas de signaux sexuels auditifs innés, ni chez la femelle[18], ni chez le mâle[19].
En conclusion, en l'état actuel des connaissances, seul le système olfactif chez les mammifères apparaît spécifiquement organisé pour traiter différents types de signaux innés susceptibles de déclencher une attraction sexuelle[12].
Signaux déclencheurs de l'attractivité sexuelle
En cohérence avec l'organisation biologique des systèmes sensoriels, des expériences ont mis en évidence l'importance des différentes informations (sexe, âge, statut endocrinien, génotype, etc.) véhiculées par les molécules chimiques et leur rôle dans l'attractivité sexuelle[13],[9].
D'autres signaux, non chimiques et dont il est actuellement difficile d'établir s'ils sont innés ou acquis, jouent également un rôle dans l'attractivité sexuelle. Ces signaux, qui relèvent du dimorphisme sexuel et des comportements précopulatoires (parades nuptiales), sont :
- caractéristique morphologiques (couleurs, bois, crinière, musculature) ;
- chant (oiseaux et baleines à bosse) ;
- cris (oiseaux) ;
- danse (paons) ;
- crissement des ailes (cigales et drosophiles).
- Signaux non chimiques participants à l'attractivité sexuelle
 La roue du paon.
La roue du paon. Les couleurs du perroquet.
Les couleurs du perroquet. Le bois des cerfs.
Le bois des cerfs. La crinière du lion.
La crinière du lion. La peau sexuelle de certains primates.
La peau sexuelle de certains primates.
Chez l'être humain, on a également mis en évidence différents types de signaux qui interviennent dans l'attractivité sexuelle :
Signaux féminins attractifs pour l'homme hétérosexuel :
- Signaux opérants à distance
- Signaux opérants à proximité
- Timbre de voix féminin
- Odeurs
Signaux masculins attractifs pour la femme hétérosexuelle :
- Signaux opérants à distance
- Signaux opérants à proximité
Mais, en raison de problèmes méthodologiques, il est actuellement difficile de préciser si l'attractivité de ces signaux est innée ou acquise.
Problèmes méthodologiques
Le principal problème méthodologique dans l'étude de l'attractivité sexuelle est de déterminer si la caractéristique qui déclenche une attractivité est apprise ou si elle dépend d'un processus de type inné. Car le système nerveux des mammifères est spécifiquement organisé pour réaliser de nombreux apprentissages, dès la période fœtale, ce qui implique que de nombreuses caractéristiques physiologiques, émotionnelles, comportementales et cognitives sont apprises. Pour comprendre les comportements, il est très important d'identifier les caractéristiques innées, qui sont les facteurs primordiaux dans le développement et le fonctionnement de l'organisme[22].
Il est assez fréquent que des études uniquement descriptives mettent en évidence une caractéristique qui apparaît d'abord comme innée et universelle, avant que sa nature apprise et donc relative soit révélée par des études pluridisciplinaires plus approfondies. Par exemple, certains auteurs ont supposé que des caractéristiques morphologiques, telles la peau sexuelle des primates ou le rapport taille-hanche et les seins dans l'espèce humaine, constitueraient des signaux visuels innés déclencheurs de l'attractivité sexuelle. Mais des études pluridisciplinaires ultérieures ont montré que ces caractéristiques seraient apprises et dépendraient chez l'humain plutôt du contexte culturel.
Le signal sexuel visuel de la peau sexuelle est supposé avoir remplacé la perte des signaux olfactifs[23],[24]. Ce signal visuel, par sa taille et sa couleur rouge, est suffisant pour provoquer par exemple la masturbation chez les babouins[25]. Mais ce signal de la peau sexuelle serait appris et proviendrait en fait des renforcements érotiques, puisque le primatologue Alan Dixson a obtenu les mêmes réactions comportementales par conditionnement sexuel avec un gant[26].

Le chercheur Devendra Singh a proposé que chez la femme le rapport entre la circonférence de la taille et celle des hanches, quand il s'approche de 0,7, serait un signal visuel inné déclencheur de l'attractivité sexuelle[27]. Ce signal serait inné car il active des structures cérébrales impliquées dans la motivation sexuelle[28]. Mais cette étude a été réalisée en Occident sur un petit échantillon non représentatif (14 hommes), et les résultats ne sont donc pas extrapolables à la population occidentale ni à d'autres cultures. De plus, il est très difficile d'expliquer comment le système visuel calculerait ce rapport taille-hanche, et plusieurs expérimentations montrent que le rapport taille-hanche préféré change en fonction du statut socio-économique[29] ou de l'évaluation cognitive[30]. Mais surtout, on observe que ce rapport taille-hanche change d'une société à l'autre, ce qui suggère fortement qu'il s'agit d'une caractéristique essentiellement apprise[31].
Le zoologiste Desmond Morris a proposé que les seins correspondraient chez l'être humain à des signaux sexuels innés[32]. Mais les mamelles ne jouent aucun rôle dans la copulation dans toutes les autres espèces de mammifères, et les seins ne sont pas considérés comme « sexuels », et ne sont pas perçus comme attractifs dans toutes les sociétés humaines[33]. Par exemple, les Mangaians « sont complètement sidérés par l'intérêt que portent les Américains et les Européens aux seins des femmes. Les Polynésiens considèrent que cet organe ne peut intéresser qu'un bébé affamé »[34].
D'autres caractéristiques qui sont perçues comme sexuellement attractives dans certaines sociétés traditionnelles (voir photographies ci-après) sont également très probablement le résultat d'apprentissages culturels.
- Caractéristiques culturelles jugées attractives dans certaines sociétés traditionnelles.
.jpg.webp) Plateau labial et scarification (femme de la vallée de l'Omo, Éthiopie).
Plateau labial et scarification (femme de la vallée de l'Omo, Éthiopie). Peintures faciales (danse yaake des hommes wodaabes, Niger).
Peintures faciales (danse yaake des hommes wodaabes, Niger). Collier-spirale (femme padaung, Birmanie).
Collier-spirale (femme padaung, Birmanie).
En conclusion, tant que des études pluridisciplinaires n'auront pas décrit en détail les différents processus par lesquels des signaux non chimiques peuvent déclencher une attraction sexuelle, il est plutôt probable que l'attractivité de ces signaux provienne d'apprentissages[8].
Notes et références
Notes
Références
- ↑ (en) Maryanne L. Fisher, The Oxford Handbook of Women and Competition, Oxford University Press, (ISBN 978-0-19-937639-1, lire en ligne), p. 603
- ↑ Nettle D, « Women's height, reproductive success and the evolution of sexual dimorphism in modern humans », Proceedings. Biological Sciences, vol. 269, no 1503, , p. 1919–23 (PMID 12350254, PMCID 1691114, DOI 10.1098/rspb.2002.2111, lire en ligne)
- ↑ Science Post - Étude – Voici le type de corps masculin que les femmes préfèrent - Yohan Demeure - 14 mai 2021 : «Publiée en 2017 dans la revue Proceedings of the Royal Society B, une étude de la Griffith University (Australie) avait récolté des informations sur le type de corps masculins que les femmes préfèrent. [...] Selon les résultats, les femmes préfèrent les torses arborant de fiers abdominaux. L’étude est d’ailleurs unanime puisqu’aucune femme interrogée n’a déclaré préférer les corps masculins moins dessinés.»
- ↑ Glassenberg AN, Feinberg DR, Jones BC, Little AC, Debruine LM, « Sex-dimorphic face shape preference in heterosexual and homosexual men and women », Archives of Sexual Behavior, vol. 39, no 6, , p. 1289–96 (PMID 19830539, DOI 10.1007/s10508-009-9559-6, S2CID 25066289, lire en ligne)
- ↑ Sell A, Lukazsweski AW, Townsley M, « Cues of upper body strength account for most of the variance in men's bodily attractiveness », Proceedings. Biological Sciences, vol. 284, no 1869, , p. 20171819 (PMID 29237852, PMCID 5745404, DOI 10.1098/rspb.2017.1819)
- ↑ Mautz BS, Wong BB, Peters RA, Jennions MD, « Penis size interacts with body shape and height to influence male attractiveness », Proceedings of the National Academy of Sciences of the United States of America, vol. 110, no 17, , p. 6925–30 (PMID 23569234, PMCID 3637716, DOI 10.1073/pnas.1219361110
 , Bibcode 2013PNAS..110.6925M)
, Bibcode 2013PNAS..110.6925M) - 1 2 (en) Agmo A. « Functional and dysfunctional sexual behavior »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- 1 2 3 Wunsch S., Comprendre les origines de la sexualité humaine. Neurosciences, éthologie, anthropologie [PDF], L'Esprit du Temps, 2014.
- 1 2 (en) Keller M., Bakker J. Special issue (12 articles) : Pheromonal communication in higher vertebrates and its implication for reproductive function. Behavioural Brain Research, 200(2):237-358, 2009.
- ↑ Kow L.M., Florea C., Schwanzel-Fukuda M., Devidze N., Kami K.H., Lee A., Zhou J., Maclaughlin D., Donahoe P., Pfaff D. Development of a sexually differentiated behavior [lordosis] and its underlying CNS arousal functions. Curr. Top. Dev. Biol., 79:37-59, 2007.
- 1 2 Pfaus J.G., Kippin T.E., Coria-Avila G.A., Gelez H., Afonso V.M., Ismail N., Parada M. Who, what, where, when (and maybe even why)? How the experience of sexual reward connects sexual desire, preference, and performance. Archives of Sexual Behavior, 41(1):31-62, 2012.
- 1 2 3 Wunsch S., Le comportement sexuel (thèse de doctorat), EPHE-Sorbonne, Paris, 2007. Serveur des thèses du CNRS [PDF].
- 1 2 (en) Ferrero D.M. et Liberles S.D., The secret codes of mammalian scents, WIREs Systems Biology and Medicine, 2:23-33, 2010.
- 1 2 3 (en) Breedlove S.M., Watson N.V. Biological Psychology. An introduction to behavioral, cognitive, and clinical neuroscience. Sinauer Associates, Seven edition, 2013.
- ↑ (en) Matsumoto J., Urakawa S., Hori E., de Araujo M.F., Sakuma Y., Ono T., Nishijo H. Neuronal responses in the nucleus accumbens shell during sexual behavior in male rats. The Journal of Neuroscience, 32(5):1672-1686, 2012.
- ↑ (en) Cibrian-Llanderal T., Tecamachaltzi-Silvaran M., Triana-Del R.R., Pfaus J.G., Manzo J., Coria-Avila G.A. Clitoral stimulation modulates appetitive sexual behavior and facilitates reproduction in rats. Physiology & Behavior, 100(2):148-153, 2010.
- 1 2 (en) Yoon H., Enquist L.W., Dulac C. Olfactory inputs to hypothalamic neurons controlling reproduction and fertility. Cell, 123(4):669-682, 2005.
- ↑ (en) Snoeren E.M., Agmo A. The incentive value of males' 50-kHz ultrasonic vocalizations for female rats (Rattus norvegicus). J. Comp Psychol., 128(1):40-55, 2014.
- ↑ (en) Snoeren E.M., Agmo A. Female ultrasonic vocalizations have no incentive value for male rats. Behavioral Neuroscience, 127(3):439-450, 2013.
- ↑ Au féminin - Sexualité - Pourquoi les hommes aiment-ils les seins ?, sur aufeminin.com.
- ↑ (en) Feinberg D.R., Jones B.C., Law Smith M.J. et al., Menstrual cycle, trait estrogen level, and masculinity preferences in the human voice, Horm Behav., février 2006, 49(2):215-22.
- ↑ (en) Doty R.L., The Great Pheromone Myth (en), Johns Hopkins Press, 2010.
- ↑ (en) Nei M., Niimura Y. et Nozawa M., The evolution of animal chemosensory receptor gene repertoires: roles of chance and necessity, Nat. Rev. Genet., 9(12), 951-963, 2008.
- ↑ (en) Zhang J., Webb D.M. Evolutionary deterioration of the vomeronasal pheromone transduction pathway in catarrhine primates. Proceedings of the National Academy of Sciences of the United States of America, 100(14):8337-8341, 2003.
- ↑ (en) Girolami L., Bielert C. Female perineal swelling and its effects on male sexual arousal: an apparent sexual releaser in the chacma baboon (Papio ursinus). Int. J. Primatol., 8:651-666, 1987.
- ↑ (en) Dixson A.F. Primate sexuality: Comparative studies of the Prosimians, Monkeys, Apes, and Human Beings. Oxford University Press, 2009.
- ↑ (en) Singh D. Adaptive significance of female physical attractiveness: role of waist-to-hip ratio. J. Pers. Soc. Psychol., 65(2):293-307, 1993.
- ↑ (en) Steven M. Platek et Devendra Singh, « Optimal Waist-to-Hip Ratios in Women Activate Neural Reward Centers in Men », PLOS One, vol. 5, no 2, , e9042 (ISSN 1932-6203, DOI 10.1371/journal.pone.0009042, lire en ligne, consulté le ).
- ↑ (en) Swami V., Knight D., Tovee M.J., Davies P., Furnham A. Preferences for female body size in Britain and the South Pacific. Body Image, 4(2):219-223, 2007.
- ↑ (en) Swami V., Furnham A., Chamorro-Premuzic T., Akbar K., Gordon N., Harris T., Finch J., Tovee M.J. More than just skin deep? Personality information influences men's ratings of the attractiveness of women's body sizes. J. Soc. Psychol., 150(6):628-647, 2010.
- ↑ (en) Yu D.W. et Shepard G.H., Jr., Is beauty in the eye of the beholder?, Nature, 396(6709), 321-322, 1998.
- ↑ (en) Desmond Morris, Le Singe nu, Grasset 1968.
- ↑ (en) Mascia-Lees F., Why women have breasts, Anthropology Now, 1(1), 4-11, 2009.
- ↑ Marshall D.S., Sexual behavior on Mangaia, dans Marshall D. S., Suggs Robert C. (Éds), Human sexual behavior: Variations in the ethnographic spectrum, Basic Books, (5), 103-162, 1971.
Annexes
Articles connexes
- Préférence sexuelle
- Orientation sexuelle
- Sex-symbol
- Bombe sexuelle
- The Mating Mind
Liens externes
- (en) FaceResearch, Recherche scientifique et études en ligne sur le rôle du visage dans l'attirance sexuelle, sur faceresearch.org