La dominance est, en génétique, une relation entre deux allèles d'un même gène, dans laquelle le phénotype des individus hétérozygotes est déterminé intégralement par l'un des allèles, sans contribution du second. Les deux allèles sont qualifiés de dominant et récessif l'un par rapport à l'autre. On dit aussi qu'un allèle dominant n'a besoin d'être présent qu'en un seul exemplaire pour être exprimé. Ce type de dominance s’appelle la dominance complète, un allèle dominant masque l’expression de l’allèle récessif chez les organismes hétérozygotes.
Les caractères phénotypiques associés à l'allèle dominant sont alors exprimés à la fois chez les individus homozygotes pour cet allèle et chez les hétérozygotes, tandis que les caractères associés à l'allèle récessif ne s'expriment que chez les individus homozygotes pour cet allèle.
De plus, il existe d'autres formes de dominance telles que la dominance incomplète et la codominance.
Ce système dominant/récessif s'applique uniquement aux caractères héréditaires mendéliens. Par exemple les caractères «couleur des cheveux» ou «couleur des yeux» sont des caractères polygéniques, c'est-à-dire que le phénotype de ces caractères est déterminé par l'action simultanée de plusieurs gènes. La détermination du phénotype pour ces caractères suit donc des procédures beaucoup plus complexes.
Exemple
Soit le gène A déterminant le caractère «présence ou non de taches de rousseur», avec l'allèle dominant A "tâches de rousseur" et l'allèle récessif a "absence de tâches de rousseur" . Un individu de génotype (A/a) aura le phénotype de l'allèle dominant, c'est-à-dire des tâches de rousseur.
Historique
Gregor Mendel
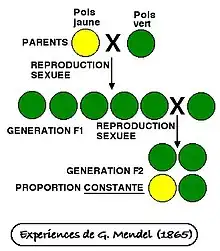
Le concept de dominance a été introduit par Gregor Johann Mendel, le « père de la génétique ». En effet, en 1866, Gregor Mendel publie les résultats de ses expériences de croisement sur les pois, plante choisie car elle atteint sa maturité sexuelle rapidement (en quelques semaines) et présente des traits facilement identifiables. Ainsi, lors de ses expériences, Mendel examine 7 traits : la couleur des graines et la couleur des gousses (verte ou jaune), la texture des graines (ridée ou lisse), la forme des gousses (enflée ou comprimée), la couleur des fleurs (pourpres ou blanches), la position des fleurs (axiale ou terminale) et la taille des plants (petit ou grand).
1re étape de son expérience
Dans un premier temps, Mendel s’intéresse à la couleur des graines des pois pour vérifier si les plantes à graines jaunes et vertes sont des souches pures. Si c’est le cas, leur descendance aura toujours la même couleur de graines que celle de leurs parents (qu’on appelle lignée pure). Mendel réalise alors une autopollinisation des plantes avec des graines vertes et des plantes avec des graines jaunes. Pour cela, il dépose le pollen de chaque plante à son propre stigmate, permettant la fécondation de l’ovule de chaque plante par son propre pollen. Il observe que les graines obtenues portent uniquement des informations génétiques identiques à leurs parents. En effet, toutes les plantes à graines jaunes autofécondées n’ont donné que des descendants à graines jaunes, et toutes les plantes à graines vertes autofécondées n’avaient que des descendants à graines vertes. Ce croisement, appelé croisement P, a permis à Mendel de s’assurer que ses plants de pois étaient de véritables souches pures.
2e étape de son expérience
Dans un deuxième temps, Mendel effectue une pollinisation croisée et artificielle de ses pois, en retirant les étamines afin d’empêcher une autofécondation. En effet, il croise les plantes pures à graines jaunes avec les plantes pures à graines vertes et observe une première descendance, notée F1, présentant uniquement des graines vertes.
3e étape de son expérience
Dans un troisième temps, Mendel croise une plante à graines vertes F1 avec une autre plante à graines vertes F1 et observe la réapparition du caractère jaune dans les descendants issus de la deuxième génération (notée F2) : ¾ de pois à graines vertes et ¼ de pois à graines jaunes sont obtenues. La descendance avait donc un rapport graines vertes/graines jaunes de 3∶1. Mendel en déduit que le facteur responsable de la coloration jaune n’avait pas disparu, mais qu’il avait été masqué par le facteur responsable de la coloration verte. De cette façon, Mendel a conclu que certains caractères sont dominants et d’autres récessifs. Les caractères dominants apparaissent toujours si l’organisme hérite de l’information génétique qui code pour le caractère. Les caractères récessifs, eux, peuvent disparaître lors de la pollinisation croisée.
Conclusion de ses analyses
À partir de cette expérience, Mendel a établi les lois de l’hérédité, dont la loi de la dominance. Cette loi affirme qu'une version du caractère cachera une autre version du caractère si l'organisme en possède deux versions différentes. Dans ce cas, les deux versions du gène sont dites hétérozygotes et l'organisme est hybride. Si un organisme reçoit deux versions du même caractère, cette paire de gènes est dite homozygote. C’est le seul cas où le caractère récessif peut être révélé. Plus simplement, si les descendants d'un croisement impliquant deux lignées pures et différant par un seul caractère présentent tous le même phénotype, ce phénotype (caractère) est qualifié de dominant[1].
Les expériences de Thomas Hunt Morgan
En 1908, Thomas Hunt Morgan, embryologiste et généticien américain, commence à étudier Drosophila melanogaster, la mouche du vinaigre, pour mettre en évidence les changements brusques de caractères héréditaires[2]. Ces expériences surviennent à la suite de la redécouverte des lois de Mendel au début du siècle, une théorie à laquelle il ne croit pas[3].
En 1910, ses élèves, Alfred H. Sturtevant, Hermann J. Muller et Calvin B. Bridges, remarquent une mouche mâle avec des yeux blancs tandis que toutes les autres ont des yeux rouges[2]. L’insecte est isolé puis croisé avec une femelle aux yeux rouges. Les descendants du premier croisement ont tous les yeux rouges. Le phénotype semble avoir disparu. Il réapparait avec le deuxième croisement : on a alors 782 mâles aux yeux blancs sur 3470 mouches, soit environ un quart de la population[3].
Morgan confirme alors les théories de Mendel sur la dominance et la récessivité. Il détermine également la théorie de l’hérédité liée au sexe. Il confirme que les chromosomes sont le support des gènes. Par la suite, Morgan et ses élèves établissent les premières cartes génétiques donnant la position des gènes sur les chromosomes[2].
La controverse de 1930
L’histoire de la dominance génétique commence par une controverse, une des plus violentes qu'ait connu la biologie de l'évolution et qui oppose dans les années 1930 les deux acteurs principaux de la discipline, Ronald Aylmer Fisher et Sewall Green Wright[4]. Ils vont initier une nouvelle avancée en faisant la synthèse des théories et des principes précédents. Celle-ci est une application des lois mendéliennes à l’échelle des populations en utilisant des outils statistiques, ce qui permettra de faire le lien avec la théorie de la sélection naturelle de Darwin: c’est la fusion de la génétique et de l’évolution[5].
Fisher, qui est un statisticien et généticien britannique, fut le premier à proposer l'idée d'une évolution de la dominance: il pense que la dominance peut être une conséquence directe de la sélection naturelle, les meilleurs allèles étant sélectionnés et devenant «dominants».
Wright, un généticien Américain, pense pour sa part que la sélection naturelle n’exerce pas une force suffisante pour expliquer le phénomène. Il introduit un principe connu sous le nom de «dérive génétique» c’est-à-dire le fait que des événements dus au hasard, se produisant au sein de petites populations, peuvent entrainer, indépendamment de la sélection naturelle, des conséquences importantes en ce qui concerne la modification des fréquences des gènes.
Fisher et Wright ont permis de donner une base mathématique à la théorie évolutionniste, utilisant à cet effet les méthodes statistiques[6].
Finalement, après de nombreuses révocations, l'explication de Wright est devenue un fait établi en génétique et le débat a été largement clos. Cependant, certains traits peuvent voir leur dominance influencée par des mécanismes évolutifs[4].
Depuis 2010
Ces dernières années, de nouvelles recherches ont été effectuées notamment sur le système d’auto-incompatibilité chez les plantes à fleurs.
En 2010, une équipe japonaise confirme la théorie de l’existence d’un modifieur de dominance proposé par Wright dans les années 1930. Ce modifieur est un petit ARN codé par une portion d’un allèle qui inhibe l’expression d’un autre allèle en reconnaissant une partie de sa séquence[7].
Grâce à la combinaison de bio-informatique et de validation fonctionnelle par transformation, cette théorie est confirmée par une expérience effectuée chez le chou pour une paire d’allèles d’auto-incompatibilité codant pour le déterminant du pollen[7].
Une nouvelle étude est publiée dans le magazine Sciences en 2014. Six allèles ont été sélectionnés chez Arabidopsis halleri par les chercheurs. À la suite d’une centaine de croisements contrôlés, ils obtiennent un grand nombre de combinaisons de couples d’allèles. Ils déterminent ensuite leurs phénotypes pour identifier l’allèle dominant de chaque couple[8].
Ces croisements démontrent qu’il existe une hiérarchie linéaire dans les relations de dominance. Ainsi, le premier allèle est dominant sur tous les autres, le deuxième est dominant sur tous les autres sauf le premier, jusqu’au dernier allèle qui va alors être récessif vis-à-vis de tous les autres.
Couplé à des méthodes de bio-informatique, des petits ARN et leurs cibles potentielles ont été recherchés sur les six allèles sélectionnés. Dans plus de 90% des relations de dominance étudiées, on observe un petit ARN sécrété par l’allèle dominant qui va alors cibler précisément l’allèle récessif. On parle de système petit ARN-cible.
Des transformations génétiques sont ensuite effectuées chez Arabidopsis thaliana (dépourvu de système d’auto-incompatibilité). Les chercheurs importent des éléments du système d’auto-incompatibilité d’Arabidopsis halleri avec un système petit ARN-cible spécialement sélectionné. Ils remarquent alors que la présence du complexe petit ARN-cible est essentiel pour qu’il y ait une relation de dominance[8].
En étendant les analyses à un plus grand nombre d’allèles, les chercheurs observent que la mise en place de ce type de réseau de dominance peut être due à des processus de duplication, délétion et diversification des petits ARNs et de leurs cibles.
Par l’acquisition de petits ARN, certains allèles sont devenus dominants tandis que l’acquisition de cibles a rendu d’autres allèles récessifs[8].
Les types de dominance
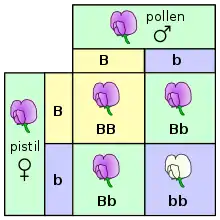
La dominance complète
La dominance complète est une forme de dominance à l’état hétérozygote dans laquelle l’allèle considéré comme dominant masque complètement l’effet de l’allèle récessif[9]. En conséquence, les génotypes homozygotes dominant et hétérozygote engendrent l’un et l’autre un phénotype dominant. Le phénotype récessif apparaît seulement chez les homozygotes récessifs[10].
Par exemple, soit B, l’allèle dominant qui donne une couleur « violette » à la fleur et soit b, l’allèle récessif qui donne une couleur « blanche » à la fleur. À la suite du croisement des deux fleurs hétérozygotes (Bb), trois génotypes sont distinguables : (BB) et (bb) qui sont homozygotes et (Bb) qui est hétérozygote. Les individus portant les deux allèles dominants (BB) présentent le phénotype dominant, la fleur est de couleur « violette ». Les individus portant les deux allèles récessifs (bb) présentent le phénotype récessif, la fleur est de couleur « blanche ». Les individus hétérozygotes porteurs à la fois d’un allèle dominant B et d’un allèle récessif b engendrent un phénotype dominant identique au phénotype de l’homozygote (BB). On parle de dominance complète : l’allèle dominant B masque l’expression de l’allèle récessif b chez les organismes hétérozygotes.
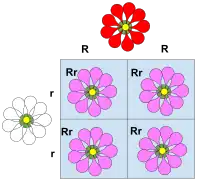
La dominance incomplète
La dominance incomplète (également appelée dominance partielle) est une exception aux lois de Mendel. Il s'agit d'une forme de dominance à l'état hétérozygote dans laquelle les allèles d’un organisme hétérozygote se combinent pour constituer un nouveau phénotype distinct. Le phénotype de l'hétérozygote est souvent un intermédiaire par rapport aux phénotypes des parents homozygotes croisés.
Par exemple, le caractère couleur de la fleur du muflier est homozygote pour le rouge ou le blanc. Lorsque l'on effectue un croisement entre la fleur homozygote rouge et la fleur homozygote blanche, le résultat est une fleur de muflier rose. Le muflier rose est le résultat d'une dominance incomplète.
Un type similaire de dominance incomplète se retrouve dans la plante Belle-de-nuit, où la couleur rose est produite lorsque des parents de souche pure de fleurs blanches et rouges sont croisés.
En génétique quantitative, où les phénotypes sont mesurés et traités numériquement, si le phénotype d'un hétérozygote se situe exactement entre (numériquement) celui des deux homozygotes, on dit que le phénotype ne présente aucune dominance, c'est-à-dire que la dominance existe uniquement lorsque la mesure du phénotype de l'hétérozygote est plus proche d'un homozygote que de l'autre.
Lorsque les plantes de la génération F1 sont autopollinisées, le rapport phénotypique et génotypique de la génération F2 sera de 1:2:1 (Rouge:Rose:Blanc) [11].
La codominance
On parle de codominance lorsque les différents allèles d'un gène ont le même effet à l'état hétérozygote et apparaissent dans le phénotype. C'est-à-dire que ces deux derniers s'expriment et qu'il y a absence d'une relation dominance-récessivité.
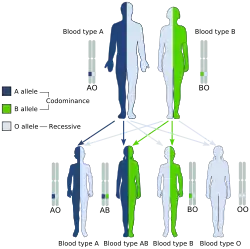
Une condition préalable à la codominance est qu'un organisme soit au moins diploïde, c'est-à-dire qu'il possède au moins un double jeu de chromosomes, c'est le cas de la plupart des animaux et des plantes. De plus, le support doit avoir des variantes différentes d'un certain gène, au moins deux allèles différents, c'est-à-dire être hétérozygote. La codominance ne peut donc pas se produire si les allèles présents sont les mêmes, en homozygotie.
Un exemple de codominance est l'hérédité des caractéristiques des groupes sanguins A et B dans le système ABO. Si une personne a hérité de la constitution génétique du groupe sanguin A d'un parent et de celle du groupe sanguin B de l'autre parent, ses globules rouges ont à la fois les antigènes A et B. Son groupe sanguin est donc AB. En revanche, si les allèles sont hétérozygotes, l'hérédité est dominante / récessive pour A et O ainsi que pour B et O. Les caractéristiques A et B prévalent sur O, l'absence des deux antigènes. Parce que la présence de l'allèle dominant respectif sur un seul chromosome de la paire de chromosomes est suffisante pour former les antigènes correspondants. Un autre exemple de codominance est le système de groupe sanguin MNS moins connu[12].
Nomenclature
En génétique, les symboles commencent par des substituts algébriques. Lorsqu'un allèle est dominant par rapport à un autre, la convention la plus ancienne consiste à symboliser l'allèle dominant par une lettre majuscule. L'allèle récessif se voit attribuer la même lettre en minuscule. Dans l'exemple du pois, une fois que la relation de dominance entre les deux allèles est connue, il est possible de désigner l'allèle dominant qui produit une forme ronde par un symbole R en majuscule, et l'allèle récessif qui produit une forme ridée par un symbole r en minuscule.
Les génotypes homozygote dominant, hétérozygote et homozygote récessif s'écrivent alors RR, Rr et rr, respectivement. Il serait également possible de désigner les deux allèles par W et w, et les trois génotypes WW, Ww et ww, les deux premiers produisant des pois ronds et le troisième des pois ridés. Le choix de "R" ou "W" comme symbole de l'allèle dominant ne préjuge pas du fait que l'allèle causant le phénotype "rond" ou "ridé" lorsqu'il est homozygote est le dominant.Un gène peut avoir plusieurs allèles. Chaque allèle est symbolisé par le symbole du locus suivi d'un exposant unique. Chez de nombreuses espèces, l'allèle le plus courant dans la population sauvage est désigné comme l'allèle de type sauvage. Il est symbolisé par un caractère + en exposant. Les autres allèles sont dominants ou récessifs par rapport à l'allèle de type sauvage. Pour les allèles récessifs, le symbole du locus est en lettres minuscules. Pour les allèles ayant un degré quelconque de dominance par rapport à l'allèle de type sauvage, la première lettre du symbole du locus est en majuscule.
Par exemple, voici quelques-uns des allèles du locus a de la souris de laboratoire, Mus musculus : Ay, jaune dominant ; a+, type sauvage ; et abt, noir et feu. L'allèle abt est récessif par rapport à l'allèle de type sauvage, et l'allèle Ay est codominant par rapport à l'allèle de type sauvage. L'allèle Ay est également codominant par rapport à l'allèle abt, mais la démonstration de cette relation dépasse les limites des règles de la nomenclature génétique de la souris.
Les règles de la nomenclature génétique ont évolué au fur et à mesure que la génétique devenait plus complexe. Des comités ont normalisé les règles pour certaines espèces, mais pas pour toutes. Les règles peuvent légèrement changer d’une espèce à une autre[13],[14].
Relations avec d'autres concepts génétiques
Allèles multiples ou polyallèles
Bien que tout individu d'un organisme diploïde possède au maximum deux allèles différents à un locus donné (sauf aneuploïdies), en effet un provenant de la mère et l'autre du père. La plupart des gènes existent dans un grand nombre de versions alléliques dans l'ensemble de la population. L'allélisme multiple est un héritage non-Mendélien, impliquant plus de deux allèles. On pourrait retrouver jusqu'à 5 allèles différents pour un seul et même gène, de ce fait cela signifie qu'il y a plus de deux phénotypes possibles, dépendant de la dominance ou la récessivité des allèles [15].
Prenons l'exemple de la couleur du pelage des chats domestiques, affectée par une série d'allèles du gène TYR (qui code pour l'enzyme tyrosinase).
Il y a 4 allèles pour ce gène:
- C conférant une couleur complète du pelage
- cb conférant un pelage de type birman (Sacré de Birmanie)
- cs conférant un pelage de type siamois (chat siamois)
- ca conférant un pelage de type albinos (chat albinos)
L'allèle "C" est complètement dominant sur les trois autres allèles, c'est pour cela que l'on va l'écrire, selon les règles de nomenclature avec un C. Au contraire, l'allèle ca conférant le caractère albinisme au pelage du chat est totalement récessif aux trois autres allèles. De ce fait nous avons alors C > cb > cs > ca.
Dominance autosomique vs dominance gonosomique
Chez l’espèce humaine et certaines autres espèces animales, le sexe des individus est déterminé par 2 gonosomes (chromosomes sexuels) appelés chromosome X et chromosome Y[16]. Les femelles portent deux chromosomes X (XX) tandis que les mâles portent un chromosome X et un chromosome Y (XY). Le reste du caryotype est commun aux deux sexes et est composé de chromosomes non sexuels appelés autosomes[17]. Les caractères génétiques associés aux gènes portés par ces chromosomes sont dits autosomiques et peuvent être dominants ou récessifs. Les caractères génétiques associés aux chromosomes X et Y sont, quant-à-eux, dits « liés au sexe » ou gonosomiques. On leur doit cette appellation en raison de leur lien physique avec les gonosomes et non car ils sont caractéristiques d’un sexe en particulier. Dans les faits, le terme « lié au sexe » fait presque toujours référence aux caractères lié à l’X et un grand nombre d’entre eux (tels que la déficience de la vision des couleurs rouge et verte) ne sont pas affectés par le sexe. Les femelles possèdent donc deux copies de chacun de leurs gènes localisés sur le chromosome X, comme pour les autosomes de leur caryotype, avec les mêmes relations de dominance et de récessivité entre les allèles. Cependant, les mâles ne possèdent qu’une seule copie des gènes portés par le chromosome X, ils sont ainsi considérés comme hétérozygotes pour ces gènes. Le chromosome Y, quant-à lui, est beaucoup plus petit que le chromosome X et porte donc moins de gènes. Parmi ces gènes se trouvent ceux responsables des caractères dits « masculins » comme, le gène SRY (ou TDF pour ‘facteur déterminant des testicules’ en français) qui inhibe le développement de la gonade en ovaire et favorise donc le développement des testicules. Les règles de la dominance des gènes liés au sexe sont donc déterminées par le comportement des allèles au sein du génotype des femelles car le génotype mâle ayant un seul allèle associé à l’X (sauf dans le cas d’une aneuploïdie), cet unique allèle est alors systématiquement exprimé, qu’il soit dominant ou non[18].
Il existe également d’autres systèmes comme celui les oiseaux dont les gonosomes sont opposés aux nôtres. En effet, chez eux, les mâles sont ZZ et les femelles sont ZW. Cependant, l'hérédité des caractères rappelle le système XY que nous connaissons bien.
Par exemple, les pinsons zébrés mâles peuvent porter le gène de la coloration blanche dans l'un de leurs deux chromosomes Z, mais les femelles développent toujours une coloration blanche.
Il existe également les sauterelles qui ont un système XO. Les femelles ont un chromosome XX, mais les mâles n'ont qu'un chromosome X. Il n'y a pas de chromosome Y du tout.
Epistasie
L'épistasie est un phénomène de génétique dans lequel l'effet de la mutation d'un gène dépend de la présence ou de l'absence de mutations dans un ou plusieurs autres gènes, respectivement appelés gènes modificateurs. En d'autres termes, l'effet de la mutation dépend du contexte génétique dans lequel elle apparaît. Les mutations épistatiques ont donc des effets différents lorsqu'elles sont isolées et lorsqu'elles se produisent ensemble. À l'origine, le terme d'épistasie signifiait spécifiquement que l'effet d'une variante d'un gène est masqué par celui d'un autre gène[19]: ["epi + stasis = s'asseoir sur le dessus"]. L'épistasie modifie le rapport caractéristique 9:3:3:1 attendu pour deux gènes non épistatiques.
Exemple d'une épistasie récessive : la couleur des poils chez les mammifères (souris)
Il y a 5 gènes régissant la couleur des poils: A, B, C, D et S.
Nous allons nous intéresser aux gènes A et C[20]:
Le gène A:
- l'allèle sauvage A confère le phénotype agouti
- L'allèle a confère un phénotype non-agouti (foncé)
Le gène C:
- L'allèle sauvage C confère l'expression de la couleur
- L'allèle c confère un phénotype albinos
L'allèle c est épistatique sur les autres gènes de la coloration (épistasie récessive).
Voici un tableau de croisement entre deux souris ayant, respectivement, le génotype (AaCc).
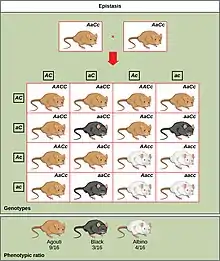
Les résultats de ce croisement sont les suivants :
- 9 souris au génotype A_C_ ayant le phénotype [agouti].
- 3 souris au génotype A_cc ayant le phénotype [albinos].
- 3 souris au génotype aaC_ ayant le phénotype [noir].
- 1 souris au génotype aacc ayant le phénotype [albinos].
Nous pouvons voir une distorsion de type 9/3/4 aux lois de Mendel typique de l'épistasie récessive.
La couleur du pelage chez la souris est donc la résultante d'interactions complexes entre plusieurs gènes.
Exemple d'une épistasie dominante : la couleur des courges en été[21]
Un croisement entre un homozygote dominant (AABB) avec un homozygote récessif (aabb) est réalisé :
F1 = 100% d'individus au génotype (AaBb) au phénotype [blanc].
- L'allèle A est le responsable de la couleur blanche.
- L'allèle B est le responsable de la couleur jaune.
- L'allèle b est la responsable de la couleur verte.
On réalise un croisement entre F1x F1, tous deux de génotypes (AaBb), la descendance est de ces proportions :
- (A_B_) = 9/16 ayant un phénotype [blanc]
- (A_bb) = 3/16 ayant un phénotype [blanc]
- (aaB_) = 3/16 ayant un phénotype [jaune]
- (aabb) = 1/16 ayant un phénotype [vert]
L'épistasie dominante a un donc une proportion différente de celle récessive, qui est de 12/3/1.
Explication et exemple de l'épistasie supplémentaire / complémentaire dans les plantes
L'épistasie supplémentaire se produit lorsque deux loci affectent le même phénotype. Par exemple, si la couleur du pigment est produite par CC ou Cc mais pas cc, et par DD ou Dd mais pas dd, alors le pigment n'est produit dans aucune combinaison génotypique avec cc ou dd. Autrement dit, les deux loci doivent avoir au moins un allèle dominant pour produire le phénotype. Cela produit un rapport caractéristique de 9:7 entre les plantes pigmentées et non pigmentées.
L'épistasie complémentaire, en revanche, produit une plante non pigmentée si et seulement si le génotype est cc et dd, et le rapport caractéristique est de 15:1 entre les plantes pigmentées et non pigmentées[22].
La génétique classique considérait les interactions épistatiques entre deux gènes à la fois. La génétique moléculaire a montré, aujourd'hui, que tous les loci génétiques sont impliqués dans des interactions complexes avec de nombreux autres gènes (par exemple, les voies métaboliques peuvent impliquer des dizaines de gènes), ce qui crée des interactions épistatiques beaucoup plus complexes que les modèles classiques à deux locus.
Principe de Hardy-Weinberg (estimation de la fréquence porteuse)
Article principal: principe de Hardy-Weinberg
La fréquence de l'état hétérozygote (qui est l'état porteur d'un trait récessif) peut être estimée à l'aide de la formule de Hardy-Weinberg :

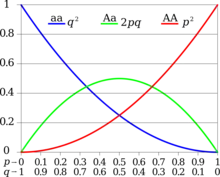
[23]Cette formule s'applique à un gène avec exactement deux allèles et relie les fréquences de ces allèles dans une grande population aux fréquences de leurs trois génotypes dans cette population. Par exemple, si p est la fréquence de l'allèle A et q est la fréquence de l'allèle a, alors les termes p², 2pq et q² sont les fréquences des génotypes AA, Aa et aa respectivement. Puisque le gène n'a que deux allèles, tous les allèles doivent être A ou a et p + q = 1. Maintenant, si A est complètement dominant sur a alors la fréquence du génotype porteur Aa ne peut pas être directement observée (puisqu'il a les mêmes traits que le génotype homozygote AA), mais elle peut être estimée à partir de la fréquence du trait récessif dans la population , puisqu'il s'agit du même que celui du génotype homozygote aa, c'est-à-dire les fréquences alléliques individuelles peuvent être estimées : q = √f(aa), p = 1 − q, et à partir de celles-ci la fréquence du génotype porteur peut être dérivée : f(Aa) = 2pq.
Cette formule repose sur un certain nombre d'hypothèses et une estimation précise de la fréquence du trait récessif. En général, toute situation réelle s'écartera de ces hypothèses dans une certaine mesure, introduisant des inexactitudes correspondantes dans l'estimation. Si le trait récessif est rare, il sera difficile d'estimer sa fréquence avec précision, car un très grand échantillon sera nécessaire.
La différence entre dominant et avantageux
Le caractère dominant d’un allèle est parfois confondu avec la notion d’avantage et le caractère récessif avec la notion de délétère, bien que cela soit des phénomènes complètement distincts. La relation de dominance des allèles décrit le concept de phénotype hétérozygote par rapport aux phénotypes homozygote sans notion d’avantage ou de désavantage de ces phénotypes. Comme beaucoup d’allèles provoquant des maladies génétiques sont récessifs et comme le mot « dominance » à une connotation positive, l'hypothèse selon laquelle le phénotype dominant est supérieur est souvent avancée. Ce n’est cependant pas vrai, bien que, comme mentionné précédemment, la plupart des maladies génétiques soit récessives et délétère, il existe des maladies génétiques dominantes ayant un effet délétère.
Néanmoins, cette confusion, omniprésente tout au long de l'histoire de la génétique, persiste encore aujourd'hui. Le principe de Hardy-Weinberg a en partie été créé et réfléchi dans le but d’éclairer cette confusion[24].
Dominance et maladies génétiques chez l'Homme
Un des grands enjeux de la génétique humaine est de prédire la transmission de maladies génétiques. Cette tâche peut s’avérer compliquée étant donné qu’il n’est pas possible de réaliser des «croisements» comme on pourrait le faire avec d’autres espèces. Il faut donc se contenter d’un nombre généralement faible de descendants, comparé à ce qu’il faudrait pour obtenir des conclusions statistiquement fiables.
L’étude de la transmission se base généralement sur l’analyse d’arbres généalogiques[25].
Plusieurs critères observables sur un arbre permettent d’identifier un mode de transmission dominant. On distinguera l’hérédité dominante autosomique de l’hérédité dominante liée au sexe[26] :
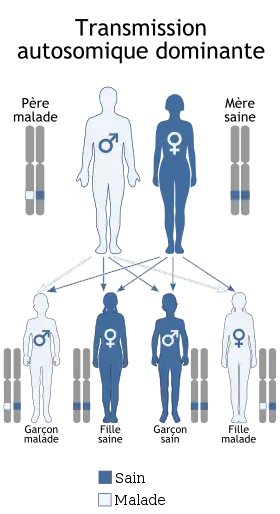
Hérédité autosomique dominante
Il existe plusieurs caractéristiques permettant de déterminer qu'une maladie est induite par une transmission autosomique dominante:
- La maladie ne saute pas de génération: un individu atteint aura forcément un parent atteint (sauf mutation)
- Un individu atteint à 50% de risque de transmettre la maladie à ses enfants (c'est-à-dire 50% à chaque grossesse)
- Deux parents sains donneront naissance à un enfant sain (sauf mutation)
- Les deux sexes sont atteints en proportions égales
Exemples de maladies à transmission autosomique dominante
- Maladie de Charcot-Marie-Tooth
- Syndrome de Marfan
- Maladie de Huntington
- Maladie d'Alzheimer[27]
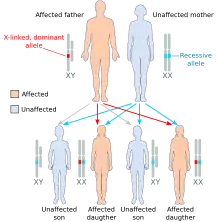
Hérédité liée au sexe dominante
Parfois confondue avec la transmission autosomique dominante, l'hérédité liée au sexe (ou gonosomique) dominante en diffère par plusieurs caractéristiques:
- On retrouve plus de femmes atteintes que d’hommes
- Un homme malade transmet la maladie à toutes ses filles mais à aucun de ses fils
- Une femme atteinte transmet la maladie à la moitié de ses descendants, quel que soit le sexe de l'enfant (père non atteint)
- Une femme homozygote dominant transmet la maladie à tous ses descendants
Exemples de maladies à transmission gonosomique dominante
- Syndrome de l'X fragile
Les mutations de novo
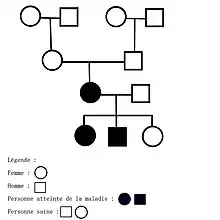
Une mutation de novo[28](terme latin signifiant "de nouveau"[29]) est une mutation survenant spontanément lors de la gamétogénèse, dans le gamète masculin ou féminin durant les premières divisions du zygote, ou bien après la fécondation de l'œuf. Elle est ainsi présente chez l'enfant, mais absente chez les parents. La maladie dominante une fois développée, a 50% de chance d'être transmise aux descendants.
Cas où la mutation apparait dans un gamète
Le risque de récidive est supposée être très faible. Cependant, pour les maladies dominantes létales, le pourcentage de mutation de novo peut monter jusqu'à 100%. On peut prendre l'exemple du nanisme thanatophore, une maladie caractérisant une dysplasie osseuse primaire[30].
Mosaïque germinale
Dans le cas d'une mosaïque germinale, la mutation est survenue dans une cellule germinale aboutissant à plusieurs gamètes. Cette situation est rare, mais reste imprévisible.
Cas particulier
- Le phénotype de la maladie peut être accentué sur un homozygote par rapport à un hétérozygote, il y a donc possibilité que l’homozygote dominant ne vive pas[31]
- Si l’allèle dominant est à pénétrance incomplète, un individu portant le gène dominant pourrait ne pas être malade
- Un organisme peut comporter un gène létal. Cela détermine un gène pouvant provoquer la mort prématurée d'un organisme portant les deux allèles de la maladie dominante (état homozygote). Lorsque l'organisme ne porte qu'un seul de ces allèles (état hétérozygote), il ne porte que la maladie sans risque de mort prématurée[32]
Mécanisme moléculaire
Zygosité
Historiquement, les lois de la recombinaison indépendantes de Mendel supposaient que les allèles ségrégeaient de manière aléatoire et indépendantes avec un allèle dit « dominant ». La zygosité, qui est le degré de similarité entre allèles d’un même organisme, peut affecter la dominance. Dans un organisme diploïde, ces allèles seraient définis par les interactions de type haplotype entre les allèles.
L’haploïdie d’un gène pourrait résulter dans un seul allèle fonctionnel qui permet à l’organisme de synthétiser suffisamment de protéine pour générer un phénotype identique à celui d’un homozygote. Trois grands types d’interactions de type haplotype sont possibles :
Haplo-suffisance
Dans un organisme diploïde, un allèle fonctionnel d’un gène haplo-suffisant serait considéré comme dominant alors qu’un allèle non fonctionnel serait considéré comme récessif. Par exemple, en supposant que la quantité standard d’enzyme produite par un homozygote soit de 100%, avec les 2 allèles contribuant chacun à 50% de la production. L’unique allèle fonctionnel de l’hétérozygote produit 50% de la quantité standard d’enzyme mais cette quantité est suffisante pour générer le phénotype standard. Si l’hétérozygote avec son unique allèle fonctionnel et l’homozygote ont le même phénotype, l’allèle fonctionnel est dominant sur l’allèle non fonctionnel. Cette situation se produit pour le locus génétique codant pour le caractère albinos : l’hétérozygote produit suffisamment d’enzyme pour convertir le pigment précurseur en mélanine, ainsi l’individu hétérozygote a une pigmentation standard. Par exemple, chez les humains et les autres organismes, la peau non pigmentée du phénotype des albinos survient lorsqu’un individu est homozygote pour un gène codant pour une version non-fonctionnelle de l’enzyme nécessaire pour produire le pigment de la peau: la mélanine.
Haplo-suffisance incomplète
Plus rarement, la présence d’un unique allèle fonctionnel engendre un phénotype anormal, bien que moins sévère qu’un individu homozygote non fonctionnel. Cette situation se produit lorsque l’allèle fonctionnel n’est pas haplo-suffisant, par conséquent les termes d’«haplo-insuffisance» ou de «dominance incomplète» sont applicables dans ces cas de figure. Les interactions intermédiaires se produisent lorsque le génotype hétérozygote produit un phénotype intermédiaire entre les 2 types d’homozygotes (dominant et récessif). Dépendamment du niveau de ressemblance entre l’homozygote et l’hétérozygote, il est dit qu’un allèle montre une dominance incomplète comparé aux autres. Par exemple, chez les humains le locus du gène Hb est responsable de la création de la protéine à chaine beta (HBB) qui est une des deux protéines globulaires qui entrent dans la composition du pigment du sang, l’hémoglobine. La plupart des gens est homozygote pour un allèle nommé HbA; plusieurs personnes possèdent une alternative de cet allèle appelé HbS ; soit en tant qu’homozygotes ou en tant qu’hétérozygotes. Les molécules d’hémoglobine de l’homozygote HbS/HbS subissent un changement de conformation qui déforme la morphologie des globules rouges. Ce qui cause une forme sévère d’anémie potentiellement mortelle nommée anémie falciforme. Les personnes hétérozygotes HbA/HbS présentent une forme bien moins prononcée de cette anomalie sous forme de « traces » d’anémie falciforme. Du fait que le phénotype induit par l’hétérozygotie HbA/HbS soit similaire mais non identique au phénotype homozygote HbA/HbA, l’allèle HbA est considéré comme partiellement dominant sur l’allèle HbS.
Haplo-insuffisance complète
Il est possible que l’unique allèle fonctionnel d’un hétérozygote ait une production de protéines insuffisante à cause d’une dominance de l’allèle non fonctionnel. Le phénotype de l’individu va donc ressembler à celui d’un individu homozygote pour l’allèle non fonctionnel au lieu de ressembler au phénotype sauvage. Ainsi, l’allèle non fonctionnel est considéré comme dominant sur l’allèle sauvage fonctionnel. Cette situation peut se produire lorsque l’allèle non fonctionnel produit une forme défectueuse qui interfère avec la fonction de la protéine fonctionnelle produite par l’allèle «standard». La présence de cette protéine défectueuse «domine» la protéine standard et le phénotype altéré de l’hétérozygote ressemble plus à celui de l’homozygote pour deux allèles défectueux. Le terme de «dominance» est souvent mal utilisé pour désigner les allèles dont le phénotype homozygote n’a pas été examiné alors qu’ils engendrent un phénotype différent de l’hétérozygote avec l’allèle normal. Ce phénomène survient dans le cadre de maladies causées par un répétition de tri-nucléotides, un exemple étant la maladie de Huntington. Dans un cas maladie de Huntington, l’haplo-insuffisance complète engendre les effets d’une protéine mutante. Une personne «normale» possède aux alentours de 20 répétitions des nucléotides C-A-G au sein de leur gène HTT mais les individus souffrant de la maladie de Huntington possèdent plus de 40 répétitions C-A-G. Un autre exemple est le syndrome de Marfan, une déficience héritée concernant le tissu conjonctif causée par le gène de la fibrilline-1 (FBN1). Une copie normale du gène FBN1 est héritée par un autre parent.
Mutation dominante négative
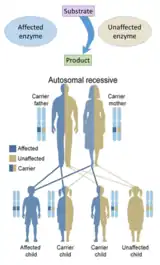
Beaucoup de protéines sont normalement actives sous forme multimérique[33], c'est un agrégat de plusieurs copies de la même protéine, mieux connue sous le nom de protéine homomultimérique. En réalité, une majorité des 83 000 différents enzymes provenant de 9 800 différents organismes (BRENDA Enzyme Database[34]) représentent des protéines homomultimériques[35]. Quand la version sauvage de la protéine est présente avec une version mutante, un mélange des deux forme un multimère. Cette mutation qui amène à une protéine mutante, qui interromps l'activité de la protéine sauvage dans le multimère, est une mutation dominante négative[36].
Une mutation dominante négative peut apparaitre dans une cellule somatique humaine et donner un avantage de prolifération à la cellule mutante, ce qui amène à une expansion de la cellule. Une mutation dominante négative dans un gène est nécessaire pour le processus de mort cellulaire (apoptose) qui intervient en réponse à des lésions de l'ADN qui peut le faire devenir résistant à l'apoptose. Ceci va permettre la prolifération de copies qui contiennent même un ADN excessivement endommagé. Ces mutations dominantes négatives font en sorte de supprimer le gène p53 [37]dans la tumeur. La protéine sauvage p53 est normalement présente sous la forme d'un multimère composé de 4 protéines (oligotetramère). La mutation dominante négative p53 apparait dans nombre de différents types de cancers et de lésions pré-cancéreuses (ex: tumeur du cerveau, cancer du sein, lésions pré-cancéreuses ORL, cancers ORL)[37].
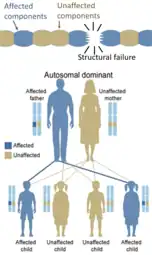
Les mutations dominantes négatives peut aussi avoir un rôle à jouer dans les gènes suppresseurs d'autres tumeurs. Depuis peu, deux lignes de germes dominant négatif mutantes ont été identifiés dans le gène Ataxia telangiectasia mutée (ATM) ce qui augmente les chances de cancer du sein[38].
Les mutations dominantes négatives qui possèdent le facteur de transcription C/EBPα peut causer une leucémie myéloïde aiguë[39]. Les mutations dominantes négatives héréditaires peuvent également augmenter le risque de maladies autres que le cancer. Les mutations dominantes négatives avec un récepteur gamma Peroxysome proliférateur activé (PPARγ)[40] sont associés avec une résistance sévère à l'insuline, diabète[41] et hypertension[39],[42].
Les mutations dominantes négatives ont également étés observées dans des organismes non-humains. En réalité, la première étude fait mention d'une protéine mutante qui inhibe le fonctionnement normal de protéines sauvages dans un mélange de multimère avec un bactériophage T4[43] qui comprend une queue en fibre de protéine GP37. Les mutations qui produisent une protéine tronquée plutôt qu'une protéine mutante complète apparaissent avoir le plus fort effet dominant négatif dans l'étude de P53, ATM, C/EBPα ainsi que le bactériophage T4[44] GP37[45].
Emploi des "dominantes" et "récessives" pour les maladies génétiques chez l'Homme
Chez l’Homme, les caractères ou maladies génétiques sont qualifiées simplement de «dominante» ou «récessive». L’appellation simplifiée «récessive», fait référence à une maladie induite par des gènes récessifs , mais ne permet pas de décrire les principes moléculaire sous-jacents. Cela peut mener à une incompréhension de la nature de la dominance.
Par exemple, la maladie récessive de la phénylcétonurie[46] résulte d’un grand nombre d’allèles situés sur locus de l’enzyme de la phénylalanine hydroxylase (PAH)[47]. La plupart de ces allèles mènent à une faible ou a une non-production de PAH. Ainsi , la phénylalanine ( substrat de la PAH) et ses produits métaboliques s’accumulent dans le système nerveux central. Cela peut entrainer de graves séquelles si ce n’est pas traité. La plupart des séquelles sont des déficiences intellectuelles.
Pour illustrer ces nuances, les génotypes et conséquences phénotypiques pour les interactions entre 3 allèles hypothétiques pour la PAH sont inscrits dans le tableau suivant[48].
| Génotype | Activité de la PAH | Concentration de phénylalanine |
Maladie de phénylcétonurie |
| AA | 100% | 60 μM | Non |
| AB | 30% | 120 μM | Non |
| CC | 5% | 200 ~ 300 μM | Hyperphénylalaninémie |
| BB | 0.3% | 600 ~ 2400 μM | Oui |
Chez les personnes saines homozygotes, portant un allèle standard fonctionnel (AA), l’activité de la PAH est standard (100%). La concentration dans le sang en phénylalanine est normale : 60 µmol / L.
Chez les personnes non traitées homozygotes pour un des allèles induisant la phénylcétonurie (BB), l’activité de la PAH est proche de zéro. La concentration en phénylalanine et de 10 à 40 fois supérieure à la concentration normale. L’individu est atteint de la phénylcétonurie.
Chez une personne hétérozygote (AB), l’activité de la PAH est de seulement 30% ( et non 50%) par rapport à la normale. La concentration en phénylalanine dans le sang est deux fois plus élevée que la normale. L’individu ne manifeste pas la phénylcétonurie. Ainsi, l’allèle A est dominant sur l’allèle B concernant la phénylcétonurie. Mais l’allèle B n’est pas complètement dominant sur l’allèle A concernant ces effets moléculaires, ce qui détermine l’activité de la PAH (0,3%, < 30% << 100%).
Enfin, l’allèle A n’est pas complètement dominant sur l’allèle B concernant la concentration dans le sang de phénylalanine, d’où des concentrations différentes (60 µM , 120 µM , 600 µM).
À noter une fois de plus qu'il n'est pas pertinent d'utiliser le terme récessif ou dominant dans ce cas où un allèle récessif entraine un phénotype plus extrême pour la concentration en phénylalanine.
Pour un troisième allèle C, un homozygote CC produit une très faible quantité de PAH, ce qui amène à une concentration de phénylalanine particulièrement haute dans le sang, ce qui amène à une hyperphénylalaninémie. Cette dernière englobe toutes les maladies liées à des déficit en PHA ou en tétrahydrobioptérine (BH4). Elle entraine aussi des déficiences mentales.
La phénylcétonurie est un type d’hyperphénylalaninémie[49].
Ainsi, la relation de dominance de deux allèles peut varier, selon le caractère du phénotype que nous observons. Il est plus pertinent de parler des conséquences phénotypiques des interactions alléliques impliquées dans un génotype, plutôt que de les placer dans des catégories «récessive» et «dominante».
Articles connexes
- Allèle
- Codominance
- Maladie génétique
- Superdominance
- Transmission autosomique récessive
- Transmission récessive liée à l'X
- Transmission autosomique dominante
Références
- ↑ « Lois de Mendel (1866) [Naissance et développement de la génétique] », sur uel.unisciel.fr (consulté le )
- 1 2 3 Éditions Larousse, « Thomas Hunt Morgan - LAROUSSE », sur www.larousse.fr (consulté le )
- 1 2 « Thomas Hunt Morgan, seigneur des mouches », Le Temps, (ISSN 1423-3967, lire en ligne, consulté le )
- 1 2 « Dominance-récessivité des gènes, épisode 1 : les hypothèses des évolutionnistes », sur INRAE Institutionnel (consulté le )
- ↑ Clément Barthélémy, Adaptation génétique et détection de la sélection dans le cadre d'évolutions expérimentales, Université de La Rochelle, (lire en ligne)
- ↑ Encyclopædia Universalis, « SEWALL WRIGHT », sur Encyclopædia Universalis (consulté le )
- 1 2 « Dominance-récessivité des gènes, épisode 2 : le mécanisme moléculaire », sur INRAE Institutionnel (consulté le )
- 1 2 3 « Dominance-récessivité des gènes, épisode 3 : retour à l’évolution », sur INRAE Institutionnel (consulté le )
- ↑ (en-US) Vix, « Complete dominance - Definition and Examples - Biology Online Dictionary », sur Biology Articles, Tutorials & Dictionary Online, (consulté le )
- ↑ « Action génétique en dominance : définition et explications », sur AquaPortail (consulté le )
- ↑ Sandra Pennington, Introduction to genetics, Blackwell Science, (ISBN 0-632-04438-1 et 978-0-632-04438-2, OCLC 40980009, lire en ligne)
- ↑ « Codominance : définition et explications », sur AquaPortail (consulté le )
- ↑ « MGI-Guidelines for Nomenclature of Genes, Genetic Markers, Alleles, & Mutations in Mouse & Rat », sur www.informatics.jax.org (consulté le )
- ↑ « Maize Genetics Nomenclature », sur www.maizegdb.org (consulté le )
- ↑ (en) « Law of Multiple Alleles », sur ThoughtCo (consulté le )
- ↑ « Définition de gonosomique - Encyclopædia Universalis », sur www.universalis.fr (consulté le )
- ↑ « Dictionnaire », sur www.universalis.fr (consulté le )
- ↑ « Fiche explicative de la leçon : Sexe et mode autosomique | Nagwa », sur www.nagwa.com (consulté le )
- ↑ (en) Rieger R, Michaelis A, Green MM, A Glossary of Genetics and Cytogenetics: Classical and Molecular, New York, Springer-Verlag, (ISBN 9780387076683)
- ↑ « Epistasi recessiva e dominante » (consulté le )
- ↑ « 6 types d'épistasis les plus importants », sur 6 types d'épistasis les plus importants (consulté le )
- ↑ « Extensions to Mendelian Analysis », sur www.mun.ca (consulté le )
- ↑ Hardy–Weinberg principle
- ↑ orphaschool, « transmission des maladies génétiques » [PDF], sur orphaschool
- ↑ Diallo N'Dalaba, « Biologie - Transmission de l’information génétique - Génétique humaine », sur www.icours.com (consulté le )
- ↑ « Hérédité et transmission », sur www.asl-hsp-france.org (consulté le )
- ↑ « Qu’est-ce que la MAAD (maladie d’Alzheimer à transmission autosomique dominante) ? | Réseau de la maladie d’Alzheimer à transmission autosomique dominante », sur dian.wustl.edu (consulté le )
- ↑ « Dictionnaire médical de l'Académie de Médecine », sur www.academie-medecine.fr (consulté le )
- ↑ « Grand dictionnaire terminologique - de novo », sur gdt.oqlf.gouv.qc.ca (consulté le )
- ↑ INSERM US14-- TOUS DROITS RESERVES, « Orphanet: Dysplasie thanatophore », sur www.orpha.net (consulté le )
- ↑ (en) « Les gènes létaux », sur Le Lab-o-Rats (consulté le )
- ↑ « Génétique », sur leschinchillasduboisjoli.com (consulté le )
- ↑ « Multimeric protein - Big Chemical Encyclopedia », sur chempedia.info (consulté le )
- ↑ Ida Schomburg, Antje Chang, Christian Ebeling et Marion Gremse, « BRENDA, the enzyme database: updates and major new developments », Nucleic Acids Research, vol. 32, no Database issue, , D431–D433 (ISSN 0305-1048, PMID 14681450, DOI 10.1093/nar/gkh081, lire en ligne, consulté le )
- ↑ Kosuke Hashimoto, Hafumi Nishi, Stephen Bryant et Anna R. Panchenko, « Caught in self-interaction: evolutionary and functional mechanisms of protein homooligomerization », Physical biology, vol. 8, no 3, , p. 035007 (ISSN 1478-3967, PMID 21572178, PMCID 3148176, DOI 10.1088/1478-3975/8/3/035007, lire en ligne, consulté le )
- ↑ (en) « Dominant-Negative Mutation (Concept Id: C1512032) - MedGen - NCBI », sur www.ncbi.nlm.nih.gov (consulté le )
- 1 2 (en) Amanda M Goh, Cynthia R Coffill et David P Lane, « The role of mutant p53 in human cancer », The Journal of Pathology, vol. 223, no 2, , p. 116–126 (DOI 10.1002/path.2784, lire en ligne, consulté le )
- ↑ (en) Georgia Chenevix-Trench, Amanda B. Spurdle, Magtouf Gatei, Helena Kelly, Anna Marsh, Xiaoqing Chen, Karen Donn, Margaret Cummings, Dale Nyholt, Mark A. Jenkins, Clare Scott, Gulietta M. Pupo, Thilo Dörk, Regina Bendix, Judy Kirk, Katherine Tucker, Margaret R. E. McCredie, John L. Hopper, Joseph Sambrook, Graham J. Mann, Kum Kum Khanna, « Dominant Negative ATM Mutations in Breast Cancer Families » (Article scientifique), sur academic.oup.com,
- 1 2 (en) Thomas Pabst, Beatrice U. Mueller, Pu Zhang et Hanna S. Radomska, « Dominant-negative mutations of CEBPA, encoding CCAAT/enhancer binding protein-α (C/EBPα), in acute myeloid leukemia », Nature Genetics, vol. 27, no 3, , p. 263–270 (ISSN 1546-1718, DOI 10.1038/85820, lire en ligne, consulté le )
- ↑ I. Barroso, M. Gurnell, V. Crowley et M. Agostini, « Dominant negative mutations in human PPARγ associated with severe insulin resistance, diabetes mellitus and hypertension », Nature, (DOI 10.1038/47254, lire en ligne, consulté le )
- ↑ I. Barroso, M. Gurnell, V. E. Crowley et M. Agostini, « Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension », Nature, vol. 402, no 6764, 1999 dec 23-30, p. 880–883 (ISSN 0028-0836, PMID 10622252, DOI 10.1038/47254, lire en ligne, consulté le )
- ↑ (en) I. Barroso, M. Gurnell, V. E. F. Crowley et M. Agostini, « Dominant negative mutations in human PPARγ associated with severe insulin resistance, diabetes mellitus and hypertension », Nature, vol. 402, no 6764, , p. 880–883 (ISSN 1476-4687, DOI 10.1038/47254, lire en ligne, consulté le )
- ↑ (en) Harris Bernstein, Kathleen M Fisher, « DOMINANCE IN BACTERIOPHAGE T4D », sur academic.oup.com,
- ↑ H. Bernstein et K. M. Fisher, « Dominance in bacteriophage T4D », Genetics, vol. 58, no 3, , p. 307–318 (ISSN 0016-6731, PMID 5662621, PMCID 1211863, DOI 10.1093/genetics/58.3.307, lire en ligne, consulté le )
- ↑ Harris Bernstein et Kathleen M. Fisher, « Dominance in Bacteriophage T4d », Genetics, vol. 58, no 3, , p. 307–318 (ISSN 0016-6731, PMID 5662621, PMCID 1211863, lire en ligne, consulté le )
- ↑ (en) OMIM, « Hyperphenylalaninemia, non-PKU mild - 261600 »
- ↑ « Entry - *612349 - PHENYLALANINE HYDROXYLASE; PAH - OMIM », sur omim.org (consulté le )
- ↑ « One Gene, One Enzyme », sur www.mun.ca (consulté le )
- ↑ « Hyperphénylalaninémie : Causes, Symptômes, Traitement Et Prévention », sur healthquery.net (consulté le )