
En biologie, la sélection naturelle est l'un des mécanismes moteurs de l'évolution des espèces qui explique le succès reproductif différentiel entre des individus d'une même espèce et le succès différentiel des gènes présents dans une population. Elle est ainsi « un avantage ou un désavantage reproductif[1], procuré par la présence ou l'absence de variations génétiques propices ou défavorables, face à un environnement qui peut se modifier[2] », le système évolutif de la nature étant un immense jeu d'essais et d'erreurs. Selon l'environnement, les individus d'une même population sont soumis à des pressions de sélection qui exercent un tri orienté de la diversité génétique, faisant de la sélection naturelle un des aspects majeurs de la biodiversité, sur la planète, comme au sein des écosystèmes et des populations.
Cette sélection est définie de manière restrictive comme l'avantage reproductif procuré par les conditions de l'environnement aux individus ayant un caractère avantageux vis-à-vis de cet environnement et leur assurant une descendance plus importante que les individus n'ayant pas ce caractère. On peut aussi la définir comme un tri qui s’opère naturellement au sein d’une espèce. Elle se traduit par la reproduction des organismes qui ont les caractéristiques leur permettant de mieux survivre dans leur milieu, cela représente le processus de la sélection naturelle. Il en résulte qu'au fil des générations, ce mécanisme explique l'adaptation des espèces à leur environnement. La théorie de la sélection naturelle permet d'expliquer et de comprendre comment l'environnement influe sur l'évolution des espèces et des populations en sélectionnant les individus les plus adaptés, et elle constitue donc un aspect fondamental de la théorie de l'évolution. De façon sommaire, la sélection naturelle est le fait que les traits qui favorisent la survie et la reproduction dans un milieu donné voient leur fréquence s'accroître d'une génération à l'autre. Cela découle « logiquement » du fait que les porteurs de ces traits ont plus de descendants, et aussi que ces derniers portent ces traits (puisqu'ils sont héréditaires).
« J'ai donné à ce principe, en vertu duquel une variation si insignifiante qu'elle soit se conserve et se perpétue, si elle est utile, le nom de sélection naturelle[3] »[4]. C'est de cette manière que l'a définie Charles Darwin.
La valeur sélective (fitness en anglais) peut représenter à la fois une estimation et une prédiction du taux de sélection naturelle.
Histoire

Alors que plusieurs théories évolutives existaient déjà sous le nom de transformisme, Charles Darwin (1809-1882) propose ce mécanisme que l'on désigne sous le terme de darwinisme ou sélection darwinienne. Le terme « sélection naturelle » a été imaginé par Darwin par opposition à la sélection artificielle pratiquée par les humains depuis des millénaires : les agriculteurs ou éleveurs choisissent à chaque génération les individus présentant les « meilleures » caractéristiques pour les faire se reproduire. Le mécanisme de sélection darwinienne permet donc d'expliquer de façon naturaliste la complexité adaptative des êtres vivants, sans avoir recours au finalisme ni à une intervention surnaturelle, d'origine divine, par exemple.
Ce principe explicatif était déjà connu à l'époque d'Aristote, bien qu'il fût rejeté par ce dernier lui préférant une explication téléologique (finaliste). Aristote donne une des premières énonciations de ce principe à partir d’un exemple :
« Les dents, par exemple, naîtraient les unes, les incisives, tranchantes et propres à couper les aliments, les autres, les molaires, larges et aptes à les broyer ; car dit-on, elles ne seraient pas produites en vue de ces fonctions, mais par accident elles s’en trouveraient capables. De même pour toutes les autres parties qui sont, selon l’opinion générale, en vue de quelque chose. Les êtres chez lesquels il s’est trouvé que toutes les parties sont telles que si elles avaient été produites en vue de quelque chose, ceux-là ont survécu étant, par un effet du hasard, convenablement constitués ; ceux, au contraire, pour qui il n’en a pas été ainsi, ont péri et périssent […]. Mais il est impossible que dans la réalité il en soit ainsi[5]. »
Mais il est plus probable, selon l'historien de la biologie André Pichot, que Darwin se soit inspiré du pasteur Joseph Townsend et de son pamphlet A Dissertation on Poor Laws (1786) qui contient un passage présentant de grandes similitudes avec le mécanisme de la sélection naturelle.
Principes
La théorie de la sélection naturelle telle qu'elle a été initialement décrite par Charles Darwin, repose sur trois principes[6] : le principe de variation, le principe d'adaptation, et le principe d'hérédité.
Variations

En général, dans une population d'individus d'une même espèce, il existe des différences plus ou moins « importantes » entre ces individus. En biologie, on appelle caractère, tout ce qui est visible et pouvant varier d'un individu à l'autre. On dit qu'il existe plusieurs traits pour un même caractère. Par exemple, chez l'être humain, la couleur de la peau, la couleur des yeux sont des caractères pour lesquels il existe de multiples variations ou traits. La variation d'un caractère chez un individu donné constitue son phénotype. C'est là, la première condition pour qu'il y ait sélection naturelle : au sein d'une population, certains caractères doivent présenter des variations, c'est le principe de variation.
Adaptations, survie et reproduction

Certains individus portent des variations qui leur permettent de se reproduire davantage que les autres, dans un environnement précis. Ils disposeraient alors d'un avantage sélectif sur leurs congénères :
- La première possibilité est, par exemple, qu'en échappant mieux aux prédateurs, en étant moins malades, en accédant plus facilement à la nourriture, ces individus atteignent plus facilement l'âge adulte, pour être aptes à la reproduction. Ceux qui ont une meilleure capacité de survie pourront donc se reproduire davantage.
- Dans le cas particulier de la reproduction sexuée, les individus ayant survécu peuvent être porteurs d'un caractère particulièrement attirant pour les partenaires de sexe opposé. Ceux-là seront capables d'engendrer une plus grande descendance en copulant davantage.
Dans les deux cas, l'augmentation de la capacité à survivre et à se reproduire se traduit par une augmentation du taux de reproduction et donc par une descendance plus nombreuse, pour les individus porteurs de ces caractéristiques. On dit alors que ce trait de caractère donné offre un avantage sélectif, par rapport à d'autres. C'est dans ce principe d'adaptation uniquement, qu'intervient le milieu de vie.
L'évolution associée à un avantage reproductif peut être très rapide à l'échelle des temps géologiques, comme pour l'évolution insulaire. Ainsi, si les conditions écologiques font qu'une petite taille est légèrement avantagée par rapport aux grandes, si bien qu'à chaque génération la taille moyenne de la population diminue de un pour mille (ce qui représenterait deux millimètres à l'échelle humaine), il suffit de 700 générations pour que la taille moyenne soit divisée par deux. De ce fait, il est exceptionnel de pouvoir observer une spéciation en cours : le phénotype d'une espèce est en pratique celui qui assure en moyenne le meilleur avantage sélectif aux individus sur l'ensemble de son aire de répartition (aire sur laquelle est assuré le brassage génétique du pool génétique) ; et ce phénotype reste donc stable tant que les conditions écologiques le sont.
Caractéristiques avantageuses héréditaires

La troisième condition pour qu'il y ait sélection naturelle est que les caractéristiques des individus doivent être héréditaires, c'est-à-dire qu'elles puissent être transmises à leur descendance. En effet certains caractères, comme le bronzage ou la culture, ne dépendent pas du génotype, c'est-à-dire l'ensemble des gènes de l'individu. Lors de la reproduction, ce sont donc les gènes qui, transmis aux descendants, entraîneront le passage de certains caractères d'une génération à l'autre. C'est le principe d'hérédité.
Ces trois premiers principes entraînent donc que les variations héréditaires qui confèrent un avantage sélectif seront davantage transmises à la génération suivante que les variations moins avantageuses. En effet, les individus qui portent les variations avantageuses se reproduisent plus. Au fil des générations, on verra donc la fréquence des gènes désavantageux diminuer jusqu'à éventuellement disparaître, tandis que les variations avantageuses se répandront dans la population, jusqu'à éventuellement être partagées par tous les membres de la population ou de l'espèce. Par exemple, dans la population humaine, la bipédie est un caractère commun à tous les êtres humains modernes.
Une histoire racontée par Richard Dawkins rappelle un point important du mécanisme darwinien : deux brontosaures voient un tyrannosaure avancer dans leur direction et courent pour lui échapper. L'un des deux dit alors à l'autre : « Pourquoi courons-nous ? Nous n'avons pas la moindre chance de courir plus vite que le tyrannosaure ! ». Et l'autre lui répond cyniquement : « Je ne cherche pas à courir plus vite que le tyrannosaure. Je cherche juste à courir plus vite que toi ! ». L'idée est de rappeler que le processus concerne moins une compétition entre espèces, qu'une compétition à l'intérieur de chaque espèce. Ce conflit ajouté au conflit sexuel sont évoqués par Thierry Lodé[7] qui explique pourquoi le conflit au sens large (conflit sexuel, conflit de reproduction, coévolution) est un puissant vecteur d'évolution, né de multiples interactions antagonistes. Darwin, sensibilisé à la sélection sexuelle par les espèces qu'il avait observées aux îles Galápagos, en avait signalé le rôle en son temps. Ces interactions et les mécanismes coévolutifs entre espèces sont étudiés également dans l'hypothèse de la reine rouge.
Adaptation des espèces à leur milieu
Lorsque des espèces sont observées dans leur milieu de vie, elles semblent toutes être profondément adaptées à chacun de leur milieu : le long cou et les longues pattes de la girafe sont en effet bien adaptés pour attraper les feuilles hautes des acacias des savanes africaines. On pourrait tout aussi bien dire que ce sont les organismes non adaptés qui n'ont pas survécu dans ce milieu.

De plus, certaines variations avantageuses dans un environnement donné peuvent devenir néfastes sous d'autres conditions. Par exemple, dans un milieu enneigé, une fourrure blanche permet de ne pas être vu par ses proies ou ses prédateurs, mais si le milieu devient forestier et plus sombre, il n'y aura plus de camouflage et les individus porteurs de fourrure blanche perdront leur avantage sélectif. La conséquence de ce phénomène est donc qu'au fil des générations, par la sélection naturelle, les caractères observés dans une population seront plus ou moins adaptés aux évolutions de son écosystème.
Autre exemple, chez les humains, la couleur de la peau foncée est une adaptation due à la sélection naturelle, et non à un bronzage qui se serait « fixé » dans certaines populations.
- En zones ensoleillées, les individus à la peau claire ont pu présenter un désavantage sélectif pour plusieurs raisons :
- ils subissent une destruction des folates (vitamine B9), dont l'absence crée des anomalies du développement embryonnaire du système nerveux (spina-bifida) et de la spermatogenèse procurant un désavantage reproductif par rapport aux individus à peau foncée ;
- dans une moindre mesure, l'augmentation du risque de développer un cancer de la peau à cause des rayons UV, puisque les cancers se développent après l'âge de reproduction.
- En zones moins ensoleillées, ces individus à peau claire seraient avantagés car la lumière du soleil permet au corps de produire de la vitamine D ; de plus, le corps économise de l'énergie et des nutriments en fabriquant moins de mélanine, le pigment de la peau.
| Répartition de l'énergie solaire reçue. | Répartition de l'intensité de la couleur de la peau humaine R. Biasutti avant 1940. |
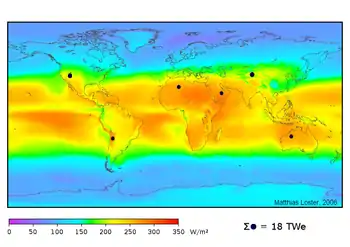 |
 |
Les facteurs de l'environnement qui peuvent donc entraîner une sélection naturelle peuvent être :
- des facteurs physico-chimiques (le biotope) : climat, milieu occupé (terrestre, aquatique, cavernicole, etc.) ;
- d'autres êtres vivants (la biocénose) : présence de prédateurs, de parasites, de microbes, de compétiteurs, etc.
Adaptations convergentes
L'adaptation des espèces à leur niche écologique peut parfois conduire deux espèces qui occupent un milieu similaire, à acquérir des ressemblances qui ne sont alors pas dues à leur éventuelle parenté. On parle dans ce cas d'évolution convergente. Ce phénomène s'interprète comme le fait que les mêmes contraintes du milieu mènent aux mêmes « solutions adaptatives ». Les yeux des vertébrés et des céphalopodes constituent l'un des exemples les plus frappants de convergence alors même que l'ancêtre commun de ces deux taxons ne possédait pas d'yeux complexes. Ces deux lignées ont évolué vers des systèmes optiques qui présentent une très forte similarité, la différence résidant surtout dans l'orientation des cellules sensorielles dans la rétine.
De tels cas de convergence évolutive sont souvent mis en avant pour argumenter en faveur d'une conception adaptationniste de l'évolution par sélection naturelle selon laquelle l'essentiel des caractéristiques observées dans les espèces vivantes ne sont pas dues au hasard mais sont le résultat de diverses pressions de sélection au cours de l'histoire évolutive des espèces.
Origine des variations héréditaires dans une population
Pour qu'il y ait sélection, encore faut-il que plusieurs variations d'un même caractère soient présentes dans la population afin que l'individu le plus adapté l'« emporte » sur les autres. En effet, dans l'exemple ci-dessus, si la totalité des individus sont identiques et porteurs de la variation phénotypique « fourrure blanche », en cas de réchauffement climatique aucun individu ne pourra survivre, et l'espèce s'éteindra. En cas de modification de l'environnement, pour qu'une espèce survive, il faut qu'elle s'adapte par la sélection naturelle. Il est donc indispensable qu'avant le changement du milieu elle présente en son sein une diversité génétique importante.
Information génétique instable

La diversité génétique dans une population d'individus a pour origine des modifications de l'information génétique dans l'ADN des cellules. Il s'agit : premièrement, de mutations ponctuelles dans la séquence d'ADN des gènes, ou de mutations plus importantes comme des réarrangements chromosomiques ; deuxièmement, de recombinaisons génétiques qui se produisent lors des transferts d'ADN des bactéries et lors de la reproduction sexuée des eucaryotes. On peut ajouter aux modifications du génome citées plus haut, une autre source de diversité génétique, soient les migrations par lesquelles le stock génétique dans une population donnée se voit renouvelé par l'arrivée d'autres membres de l'espèce porteuse d'un ensemble génétique différent.
Les modifications génétiques sont aléatoires : ce n'est pas l'environnement qui « dicte » quel gène doit muter, mais bien le hasard. C'est pour cela qu'on observe dans les populations beaucoup de variations inadaptées au milieu de vie (par exemple, les maladies génétiques rares). Une erreur courante consiste à croire que les modifications génétiques sont une conséquence de la sélection naturelle. En revanche la sélection naturelle a bien le pouvoir de cumuler les modifications génétiques adaptées, ce qui aboutit à des adaptations complexes.
Modifications de l'information génétique
C'est donc parmi la grande diversité génétique des individus, que vont ensuite être sélectionnés les phénotypes et les allèles les plus adaptés à l'environnement. Pour en faire la démonstration on pourrait simplement montrer que les phénotypes nouveaux étaient présents avant le changement du milieu. Ceci n'est pas toujours évident et quand bien même ce serait le cas, un mécanisme d'induction d'une mutation donnée par l'environnement n'est pas à exclure. C'est grâce à une astuce mathématique que Luria et Delbrück montrent que ce sont bien les mutations préexistantes dans une population de bactéries qui sont sélectionnées quand on ajoute un virus.
Hypothèse des caractères acquis
La génétique moléculaire n'existant pas à son époque, Darwin ne pouvait prendre en compte les mécanismes moléculaires à l'origine des nouveaux caractères. Sa théorie de la sélection naturelle incluait donc l'hypothèse de la transmission des caractères acquis. Dans son ouvrage de 1868, La variation des animaux et des plantes sous l'effet de la domestication, il alla même jusqu'à proposer une théorie pour cette transmission des caractères acquis[8]. Dans la théorie initiale de Darwin telle qu'il l'expose dans L'Origine des espèces, ces variations entre les individus trouvent leur origine dans le fait que des individus acquièrent des caractéristiques différentes au cours de leur vie. Ces caractères acquis seraient alors transmis à leur descendance et cela expliquerait les variations observées et l'évolution des caractéristiques de l'espèce. Toute autre source de variation reste pourtant acceptable, par exemple le hasard ; en revanche, le problème qui se poserait alors serait de savoir comment empêcher la dilution, puis l'éventuelle disparition de ces caractères s'ils ne sont pas « entretenus ».
La théorie de l'hérédité des caractères acquis a été considérée comme invalidée par August Weismann à la fin du XIXe siècle. En réponse aux néo-lamarckiens qui soutenaient le contraire, il montra que des mutilations n'étaient pas transmises. On en déduisit abusivement qu'aucun caractère acquis ne pouvait se transmettre, alors qu'une mutilation ne peut être assimilée à une acquisition par l'organisme de fonctions nouvelles comme le voulait Lamarck. On ne peut prouver avec certitude l’impossibilité d'hérédité de caractères acquis (une inexistence ne peut être prouvée qu'en mathématiques, par l'absurde). On peut en revanche chercher s'il existe un exemple réel réfutant cette impossibilité. Plusieurs recherches ont été menées en ce sens au début du XXe siècle, notamment par Paul Kammerer. Cela fait aussi l'objet de recherches dans le domaine de l'immunologie.
Les caractères innés sont bel et bien transmis au cours de la reproduction mais avec des variations qui suivent les lois de l'hérédité mendélienne, du nom de leur découvreur, Gregor Mendel (1822-1884) dont les travaux sur les lignées de pois ne furent redécouverts qu'au début du XXe siècle et étaient malheureusement ignorés de Darwin. Mendel apporte la réponse au problème de la dilution : un caractère ne s'affaiblit pas ; il est simplement transmis en tant que dominant, transmis en tant que récessif, ou éliminé ; mais chez les individus qui le portent, il reste totalement présent, ce qui assure sa pérennité s'il est favorable à son porteur (ou, pour être plus précis, à la descendance de son porteur).
En génétique des populations
La convergence entre la théorie darwinienne et la théorie de l'hérédité donnera alors naissance au cours des années 1930 à la génétique des populations, en particulier grâce aux travaux théoriques de Ronald Fisher. À la même période, grâce aux expériences de Thomas Morgan et Theodosius Dobzhansky sur les mouches drosophiles, les mécanismes moléculaires responsables des phénomènes d'hérédité génétique commenceront à être identifiés. L'une des découvertes majeures de la biologie sera alors de montrer que la diversité génétique qui garantit la variété des phénotypes est due à des modifications aléatoires du génotype (mutations, recombinaisons génétiques…) en particulier lors de sa transmission d'une génération à l'autre, au moment de la reproduction.
Même si ce n'est pas le mécanisme qui avait été envisagé par Darwin dans sa théorie de la sélection naturelle, il n'en reste pas moins que ces processus permettent de rendre parfaitement compte de la sélection naturelle dans le cadre de ce qui est considéré comme la théorie centrale de la biologie moderne, la théorie synthétique de l'évolution ou synthèse néo-darwinienne qui fait le lien entre les mécanismes au niveau de la génétique moléculaire et les phénomènes d'évolution à l'échelle des populations. Ainsi la sélection naturelle peut se « mesurer » grâce à des calculs statistiques.
Études scientifiques
 |
 | |
Les deux morphes du phalène du bouleau : le clair (à gauche) et le sombre (à droite). | ||
Il apparaît aujourd'hui évident que tout organe ayant une fonction définie, par exemple la nageoire du poisson, est une adaptation à un milieu et le résultat d'une sélection naturelle. Cependant la démonstration scientifique doit, elle, passer par la mise en évidence d'une corrélation chiffrée entre les variations d'un caractère héréditaire et celles d'un paramètre précis de l'environnement. Parmi les exemples les plus célèbres, peuvent être cités :
- Les pinsons des Galápagos : les épisodes de sécheresse furent suivis par une raréfaction des graines molles et donc par une augmentation de la taille du bec des pinsons leur permettant de briser la coquille des graines restantes, plus dures.
- La sélection naturelle chez les bactéries résistantes aux virus bactériophages a été mise en évidence par l'expérience de Luria et Delbrück. Ils ont en effet démontré pour la première fois que les mutations préexistent à la sélection et qu'elles sont bien aléatoires.
- Le mélanisme industriel de la phalène du bouleau en Angleterre, est un exemple de microévolution mis en évidence par le zoologiste britannique Bernard Kettlewell : dans cette espèce de papillons, la proportion d'individus clairs aurait diminué du fait de la pollution qui détruisait les lichens (blanc) qui se développaient sur les surfaces des troncs d'arbre sur lesquels ils se posaient. En effet, les individus clairs (initialement présents en majorité), étant désormais plus visibles que les autres sur les troncs noir, étaient plus sujets à la prédation que les autres. Les phalènes sombres qui existaient avant la pollution seraient alors devenues majoritaires. Puis, le phénomène se serait inversé lorsque les industries polluantes ont progressivement disparu permettant le retour du développement du lichen sur les troncs. En fait, cette belle histoire est discutable car apparemment les phalènes ne se posent pas sur les troncs mais sous les feuilles des bouleaux. Les modifications des fréquences des morphes sombres et clairs pourraient être des réponses à la toxicité des rejets industriels plutôt qu'à la modification de la couleur du revêtement des troncs.
Espèce humaine
La sélection naturelle produit aussi ses effets dans l'espèce humaine. La capacité chez l'adulte à digérer le lactose du lait a été sélectionnée il y a environ 9 000 ans dans les populations humaines originaires d'Europe du nord ou d'Asie centrale où on élevait du bétail pour son lait, mais pas dans d'autres populations où cet aliment est absent[9]. Récemment, il a été découvert que cette capacité était présente également chez certaines ethnies d'Afrique de l'Est, les Tutsis et Peuls du Soudan, de Tanzanie et du Kenya, mais à partir de trois autres mutations, sélectionnées indépendamment l'une de l'autre[10]. Ces dernières seraient apparues il y a 7 000 à 3 000 ans. Il s'agit là d'un exemple de convergence évolutive.
La persistance d'une maladie génétique comme la drépanocytose est due au fait que l'allèle responsable de la maladie confère aussi un avantage sélectif en augmentant la résistance au parasite responsable du paludisme.
Activités humaines
L'humain peut aussi exercer involontairement une pression de sélection sur certains organismes dont l'évolution, en retour, peut être néfaste pour l'économie ou la santé humaine :
- La résistance aux antibiotiques : par exemple, depuis 1961 les bactéries staphylocoques dorés résistantes à la méticilline se sont multipliées et rendent inefficaces le traitement par cet antibiotique dans un grand nombre de cas. Ces résistances sont à l'origine de nombreuses maladies nosocomiales en milieu hospitalier.
- La résistance des criquets pèlerins aux insecticides : comme dans l'exemple précédent, en Afrique sahélienne l'utilisation massive d'insecticide par les agriculteurs pour éradiquer les criquets qui dévorent régulièrement les cultures et peuvent entrainer une famine s'est traduite par une plus grande résistance de ceux-ci. En effet, les individus survivants au traitement insecticide ont pu engendrer plusieurs milliers de descendants en quelques générations, soit en quelques années, qui eux-mêmes ont hérité de cette capacité de résistance. L'élimination de cette nouvelle population exige alors un traitement encore plus agressif pour l'écosystème.
Coévolution et compétition
Dans les exemples précédents, il s'agit d'espèces évoluant au gré des améliorations techniques humaines mais ce phénomène peut aussi s'observer dans les interactions biologiques entre deux espèces. Un exemple très étudié d'une telle coévolution est le parasitisme de ponte chez les coucous[11]. Dans ces espèces, le parent pond ses œufs dans le nid d'autres oiseaux. Dès sa naissance, le jeune coucou expulse les œufs présents afin d'être le seul à bénéficier des soins prodigués par les parents de l'espèce hôte ainsi trompés. Parmi les espèces parasitées, certaines ont évolué vers des stratégies antiparasitiques, en l'occurrence une aptitude à distinguer les œufs de coucou de leurs propres œufs. Cela a créé une pression de sélection pour certaines espèces de coucou qui ont évolué en développant une forme de mimétisme des œufs de telle sorte que ceux-ci ressemblent étonnamment aux œufs de leurs hôtes. À leur tour, les espèces parasitées (comme la pie-grièche écorcheur) ont développé des capacités accrues de discrimination de leurs propres œufs des œufs mimétiques du coucou, capacité qui est absente chez des espèces non ou moins parasitées.
De tels phénomènes de coévolution sont fréquemment observés dans les cas de parasitisme mais parfois aussi dans certains cas de compétition interspécifique ou dans le cas de compétition intraspécifique de la sélection sexuelle[12]. La coévolution inter-sexes s'observent souvent en réponse à la sélection sexuelle post-copulatoire : par exemple, dans certaines espèces d'oiseaux[13] et d'invertébrés[14], les pénis des mâles ont une forme hélicoïdale qui fonctionne comme un goupillon éliminant la semence des autres mâles et leur permettant de déposer leurs propres gamètes au plus profond du vagin des femelles avec lesquelles ils s'accouplent afin de s'assurer la paternité de la progéniture ; dans ces mêmes espèces, on observe une augmentation proportionnelle de la taille du vagin avec une morphologie tout en sinuosités qui contrecarre les stratégies reproductives des mâles.
Sélections
Sélection artificielle d'origine anthropique
 |
 | |
Rose cultivée et Rose sauvage Rosa canina. | ||
D'autres modes de sélection existent.
« Les débuts de la sélection anthropique sont un des éléments de la révolution culturelle du néolithique. Ils peuvent être identifiés par la période pendant laquelle les humains sont intervenus dans la sélection des espèces vivantes »[15].
L'adjectif naturelle s'oppose chez Darwin au concept de sélection artificielle connue et pratiquée depuis quelques milliers d'années par les éleveurs. En effet les animaux d'élevage domestiques ou les espèces de plantes cultivées (vaches, chiens, roses…) constituent autant de variations « monstrueuses » absentes dans la nature. Elles sont le fruit de la lente sélection d'individus intéressants (pour les rendements, ou du point de vue esthétique) par les éleveurs et les agriculteurs (voir élevage sélectif des animaux). C'est cette observation qui permet à Darwin d'émettre l'hypothèse d'une sélection opérée par la nature sur les espèces sauvages.
Exemples de sélection artificielle d'origine anthropique (exercée par l'homme, notamment sur les espèces domestiques, et ce depuis les débuts de leur domestication) :
- la domestication du chien s'est accompagnée d'une meilleure capacité de son appareil digestif à dégrader, transporter et absorber l'amidon, que celle du loup. Cette domestication au Néolithique correspond à une période où le régime alimentaire humain s'est modifié en incorporant de plus en plus de céréales. En conséquence, les rebuts de nourriture jetés par les groupes humains contenaient plus d'amidon mieux digéré par le chien chez qui le génome exprime plus d'alpha-amylase, de maltase-glucoamylase (en) et d'une protéine qui transporte le glucose vers le petit intestin[16].
- Une rose cultivée résulte de mutations à partir d'une rose sauvage. Ces mutations ont été sélectionnées par les horticulteurs, elles sont répandues dans les jardins : c'est la sélection artificielle. Dans la nature des roses cultivées ne peuvent se reproduire, c'est la sélection naturelle.
- L'intensification du braconnage des éléphants pour leur ivoire favorise la naissance de pachydermes avec des défenses plus petites ou sans défenses[17].
- La sélection artificielle explique la résistance des micro-organismes aux antibiotiques, la résistance aux pesticides d'au moins 500 espèces nuisibles aux cultures, aux denrées stockées et à la santé humaine[18].
On peut aujourd'hui faire apparaître de nouvelles espèces (de drosophile notamment[19]) par sélection artificielle. Mais ce genre de pratique serait contre-productif en agriculture, à cause de la diminution de la diversité génétique de chaque espèce.
Sélections utilitaire et sexuelle
La sélection darwinienne s'appuie sur deux mécanismes conjoints : le premier est la sélection utilitaire (ou sélection de survie ou sélection écologique) ; le second est la sélection sexuelle. Dans le grand public, ce dernier aspect de sélection sexuelle est souvent ignoré et on identifie la sélection naturelle avec la sélection de survie. Or c'est une erreur car ces deux mécanismes sont bien à l'œuvre dans le monde vivant.
La sélection utilitaire correspond à un processus de tri entre individus en vertu de leur capacité à survivre et/ou à être féconds[20]. Ce terme désigne plus spécifiquement le mécanisme qui fait évoluer les espèces sous les pressions extérieures à l'espèce de la compétition interspécifique et de l'environnement ou sous la pression intérieure à l'espèce de la compétition intraspécifique. En effet, elle repose sur l'idée que pour pouvoir se reproduire, il faut d'abord survivre. Ce type de sélection favorise donc les individus capables d'échapper ou de se protéger des prédateurs mais aussi de résister aux parasites : c'est la compétition interspécifique. Cette sélection favorise aussi les individus d'une même espèce étant en compétition entre eux pour trouver des ressources dans l'environnement, qu'il s'agisse de proies ou d'autres ressources non-nutritives comme des abris (terrier, nid…) : c'est la compétition intraspécifique. Enfin, elle favorise les individus les mieux capables de résister à l'environnement biotopique, aux conditions climatiques, etc. : on parle de facteurs abiotiques.
La sélection sexuelle est un phénomène qui a lieu à une étape différente de la vie de l'individu. Elle désigne le fait qu'il y a aussi une compétition au sein de chaque espèce pour accéder aux partenaires sexuels dans le cadre de la reproduction sexuée. Cet aspect de la théorie fut pleinement développé par Darwin dans son ouvrage intitulé La Descendance de l'Homme. Dans le cadre de la sélection sexuelle, il va donc se produire une compétition intrasexuelle, entre les individus d'un même sexe, mais aussi intersexuelle, entre les individus de sexes différents (les individus d'un sexe devant choisir avec quel individu de l'autre sexe ils vont s'accoupler). La sélection sexuelle permet donc d'expliquer des traits de caractère ou des comportements qui pénalisent la survie quand ils sont analysés en dehors du contexte utilitaire, comme la queue du paon ou les bois des mégacéros. Très importante chez les oiseaux (parades nuptiales), elle pourrait être à l'origine des plumes dont elle explique la forme aplatie pour exhiber les couleurs.
Pression de sélection
Rétrospectivement, les modifications successives au cours des générations des populations peuvent sembler orientées, comme si ces modifications étaient « tirées » ou « poussées » dans une certaine direction. Par exemple, en suivant les observations de Darwin sur les pinsons des Galápagos, on peut observer que certaines espèces semblent suivre une tendance vers un élargissement du bec qui devient de plus en plus massif alors que chez d'autres espèces de pinsons, la tendance est plutôt vers un affinement du bec. Ce phénomène qui se manifeste comme une tendance apparente dans l'évolution d'une ou plusieurs espèces a reçu le nom de pression de sélection. Ces pressions de sélection sont « orientées » par les pressions dites intérieures à l'espèce (sélection sexuelle, compétition intraspécifique) et les pressions dites extérieures à l'espèce (compétition extraspécifique, environnement), c'est-à-dire tout ce qui influence la survie et la reproduction des individus.
Les pressions de sélection s'exercent différemment d'une espèce à l'autre ou d'un milieu écologique à un autre, voire d'une sous-population d'individus à une autre. Ainsi, il peut se produire au sein d'une même espèce une divergence si deux sous-populations sont soumises à des pressions de sélection légèrement différentes. Ces deux populations évolueront vers des formes différentes qu'on appelle morphes et si le phénomène se poursuit dans le temps on peut aboutir à la formation de deux espèces distinctes, c'est la spéciation sympatrique. Les deux espèces occupent alors des niches écologiques suffisamment distinctes pour qu'elles n'entrent plus directement en compétition l'une avec l'autre et suivent alors des trajectoires évolutives différentes en réponse aux pressions de sélection spécifiques auxquelles elles sont soumises.
Apparition d'adaptations biologiques complexes
Les systèmes vivants apparaissent comme très complexes et sont des adaptations tellement poussées à un milieu que les humains y trouvent une source d'innovations techniques et industrielles (par exemple, les attaches scratch ou velcro, les industries pharmaceutique et chimique). Cette complexité n'aurait pu voir le jour sans le pouvoir qu'a la sélection naturelle d'accumuler les « bonnes » innovations génétiques :
Chaque innovation évolutive apparaît de manière aléatoire. La sélection naturelle favorise ensuite chacun de ces petits « sauts » évolutifs (A, puis AB, puis ABC…, puis ABCDE). Elle permet ainsi l'apparition d'adaptations de plus en plus poussées (ABCDE). En effet, si le caractère A n'avait pas été sélectionné, le caractère ABCDE ne serait jamais apparu. Car E dépend de A. Un caractère complexe, comme une enzyme, résulte d'une accumulation d'innovations sélectionnées successivement, et non de simples apparitions indépendantes, au hasard des innovations génétiques (même si certains « sauts » évolutifs peuvent être plus ou moins importants ou graduels, voir équilibre ponctué). Donc la sélection naturelle ne fait pas que favoriser les adaptations les plus complexes ; elle permet aussi leur apparition. Cela n'est valable que si la sélection naturelle s'opère de manière continue, ce qui est envisageable par une compétition intra-spécifique, que Malthus et Darwin estiment inévitable dans une population. En effet les êtres vivants ont une tendance naturelle et universelle à se reproduire en plus grand nombre qu'à la génération précédente.
Les simples innovations dues au hasard sur quelques générations ne suffisent pas à rendre compte de la complexité des êtres vivants et de leur adaptation à leur milieu. Il faut la sélection naturelle pour accumuler les petites innovations et pour en arriver à un organe aussi complexe que l'œil de mammifère, par exemple. Cela peut être une réponse aux critiques de certains néo-créationnistes, qui affirment que les systèmes vivants (enzymes…) sont trop complexes et harmonieux pour que leur apparition ne soit due qu'à des mutations aléatoires, et que donc selon eux il n'y aurait pas eu d'évolution.
Limites d'explication dans l'évolution de la lignée humaine
Comme toutes les autres espèces animales, l'espèce humaine est le produit de l'évolution et de la sélection naturelle. Par exemple, on peut penser que la bipédie a offert, dans le passé, un tel avantage sélectif aux individus capables de se déplacer debout, que les gènes associés à ce mode de locomotion se sont répandus dans toute la population humaine. Il reste néanmoins difficile de reconstruire le détail de l'histoire évolutive de notre espèce, et en particulier d'identifier quels facteurs exacts et quels processus précis ont pu intervenir dans l'évolution humaine (e.g., sélection sexuelle, sélection de groupe, sélection culturelle, dérive génétique…) Dans certains cas, il est néanmoins possible d'identifier les pressions de sélection et les adaptations résultant de la sélection naturelle dans les populations humaines. L'un des exemples les plus documentés est la capacité de résistance au paludisme. Les individus porteurs d'un gène entraînant une anomalie de leurs cellules sanguines résistent mieux au parasite qui cause cette maladie. Par contre, leurs enfants courent le risque de souffrir de drépanocytose s'ils héritent de ce gène des deux parents. L'avantage sélectif fourni par la résistance au paludisme permet donc d'expliquer pourquoi, dans les populations humaines où ce parasite est endémique (Afrique), l'allèle d'un gène pouvant entraîner une maladie relativement grave se maintient.
L'une des tentatives les plus décriées d'appliquer la théorie darwinienne à l'espèce humaine fut l'utilisation qui fut faite de la sociobiologie développée par Wilson pour expliquer le comportement des espèces ultra-sociales (fourmis, termites, abeilles…). En effet, même s'il est vrai que l'une des caractéristiques de l'être humain (et aussi d'autres primates) est son mode de vie très social, la théorie de Wilson nécessite aussi un mode de reproduction et d'organisation sociale très particulier dite eusocialité, très différents de ce qu'on observe chez les humains. Comme s'en défend Wilson lui-même, il est donc absurde de vouloir appliquer directement les conclusions des travaux menés sur ces espèces à l'espèce humaine. Toutefois, certains outils théoriques développés initialement dans le cadre de la sociobiologie peuvent se révéler parfaitement pertinents pour l'étude de l'humain.
Le défi majeur de la paléoanthropologie reste néanmoins de parvenir à un cadre théorique pour expliquer l'évolution humaine au moyen de mécanismes plus riches que la seule sélection naturelle. Or, l'importance de phénomènes comme la culture, la sélection sexuelle, la dérive génétique reste difficile à évaluer :
« Ainsi, même dans le cadre des théories modernes de l'évolution, qu'on appelle néodarwinisme ou théorie synthétique de l'évolution, les évolutionnistes s'efforcent de réserver une place à part à l'homme, étant entendu que si son corps a évolué, il reste que ce qui fait l'humain échappe aux lois de l'évolution[21]. »
Un exemple récent d'une telle difficulté concerne le rôle de la sélection de groupe. Alors que ce mécanisme qui « favorise la survie du groupe au détriment de la survie de l'individu » a été très critiqué dans l'évolution animale, il semble que l'une des particularités de l'espèce humaine est que, justement, des phénomènes de sélection multi-niveaux ont pu jouer un rôle important au cours de son évolution[22] et en particulier dans l'évolution de sa psychologie.
Comportement et culture selon la sociobiologie
Tout comportement possède une composante génétique et héréditaire. Chez certains animaux, il a été démontré que l'environnement pouvait agir sur l'évolution d'un comportement héréditaire et inné[23]. Quant à la culture, qui ne se transmet pas par l'ADN mais par l'apprentissage, elle peut également être sujette à une sélection. Par exemple, si j'ai un comportement qui m'apporte de la satisfaction, comme m'habiller à la mode, je le reproduirai et je l'enseignerai ou on m'imitera. Et inversement, ce comportement ne sera pas transmis s'il ne donne pas satisfaction. La théorie de la mémétique émise par Richard Dawkins, désigne ces entités, qu'on appelle mèmes, comme éléments de base de la sélection que subit la culture, au même titre que le sont les gènes pour l'évolution du vivant. En retour, la culture peut entraîner de nouvelles conditions de vie, et donc modifier la sélection naturelle[24]. Par exemple, l'utilisation de lait de vache dans l'alimentation a favorisé les génotypes tolérants à la digestion du lactose (sucre présent dans le lait). Ainsi, l'évolution de la culture serait le produit d'une interaction entre une sélection naturelle et une sélection culturelle. Ainsi on peut envisager que ce modèle d'évolution faisant jouer des rétroactions puisse suivre un cercle vicieux ou vertueux, ce qui entraîne une évolution perpétuelle et continue. À condition que les rétroactions soient positives. Ce modèle est corroboré par les observations : la culture humaine suit bien une évolution continue, et les structures anatomiques qui ont permis cette évolution (volume crânien, structure du cortex) ont aussi évolué de manière continue depuis deux millions d'années, au moins.
La culture humaine, qui fait toute la singularité de notre espèce, pourrait donc être le résultat d'un tel modèle d'évolution, avec une certaine indépendance vis-à-vis de l'environnement, si on ne tient pas compte des modifications engendrées par les humains eux-mêmes. Wilson parle de coévolution[25] des gènes et de la culture. Mais cette approche est critiquée. Cela est expliqué par certains scientifiques dans la théorie de la construction de niche[26] : le comportement ou une autre activité peut influer sur l'environnement immédiat (la niche écologique) et, en retour, modifier la pression de sélection naturelle.
Autres mécanismes de l'évolution
Lorsque les fréquences de certaines variations héréditaires changent uniquement à cause du hasard, on parle de dérive, voire, si le groupe est très réduit, d'effet fondateur. Ces caractères doivent être relativement neutres pour la sélection naturelle (il n'y a ni avantage, ni désavantage sélectif). Si une même évolution se produit de manière répétée dans un même milieu au cours des générations, ce n'est pas de la dérive, mais on peut parler de sélection.
Dimension universelle
Dans le cadre de cette théorie, tout système dans lequel s'observeraient ces trois premiers principes donnerait lieu à un phénomène d'évolution par sélection naturelle. Dans le monde vivant, la transmission héréditaire de l'information génétique, qui obéit à ces trois principes, résulte donc d'une évolution des espèces par sélection naturelle. Cependant, d'un point de vue théorique, l'évolution par sélection naturelle ne dépend pas de la nature précise des mécanismes qui permettent l'apparition de variations, soit, la transmission héréditaire et la traduction de l'information héréditaire en caractères phénotypiques. Le fait que Darwin lui-même ignorait jusqu'à l'existence des gènes illustre bien le distinguo qu'il convient de faire entre le cadre théorique de la sélection darwinienne et ses manifestations observables dans les écosystèmes terrestres.
Par conséquent, les phénomènes évolutionnaires observés dans le monde vivant pourraient tout à fait se manifester dans d'autres systèmes qui mettraient en œuvre les trois principes fondamentaux de la sélection darwinienne. C'est par exemple, l'hypothèse faite en exobiologie selon laquelle des formes de vie extraterrestres pourraient être apparues sur la base de mécanismes fondamentaux différents de ceux que l'on connait de la biologie terrestre. Au sein de telles formes de vie évoluant par sélection naturelle, on devrait donc observer des phénomènes similaires à ceux que l'on connait sur Terre : adaptation, coévolution, reproduction sexuée… Les plus adaptationnistes des biologistes de l'évolution (comme Simon Conway Morris) ont ainsi proposé que des contraintes environnementales extraterrestres suffisamment fortes et similaires aux nôtres pourraient donner des formes de vie présentant d'importantes convergences évolutives avec la vie terrestre ; parmi lesquelles la présence d'yeux, l'eusocialité, ou des capacités cognitives complexes[27].
Plus près, l'application des principes de la sélection naturelle à la sphère culturelle humaine a donné lieu à la mémétique qui cherche à expliquer les variations, la transmission, et la stabilisation des phénomènes culturels par analogie avec les espèces vivantes. Dans ce cadre théorique, les mèmes sont les unités de sélection, ou réplicateurs, des phénomènes culturels. La sélection intervient sur les mèmes en fonction de leur capacité à « survivre », c'est-à-dire à persister dans l'esprit des individus, et de leur capacité à « se reproduire », c'est-à-dire à passer de l'esprit d'un individu à un autre par l'imitation, la communication, l'enseignement, etc. À chaque reproduction, un mème peut donc « muter » : à force d'être racontée, une même histoire sera par exemple déformée, c'est le principe du téléphone arabe. Ainsi, malgré les différences notables entre la nature des réplicateurs biologiques et culturels, certaines analogies peuvent être envisagées : coévolution (y compris entre gène et mème[28]) ou formation de complexes de mèmes. La pertinence de l'application de la théorie darwinienne aux phénomènes culturels reste néanmoins très débattue[29].
Enfin on peut aussi citer l'exemple des algorithmes évolutionnaires utilisés dans le domaine de l'optimisation en ingénierie. Ceux-ci permettent de rechercher une solution à un problème donné en mettant en compétition une population de solutions potentielles dont seules les meilleures sont conservées pour être recombinées et donner naissance à une nouvelle génération de solutions. Étant donné que, dans cette méthode, la sélection est le résultat d'une intervention humaine, ce cas relève plutôt de la sélection artificielle.
Notes et références
- ↑ Voir emballement fisherien, théorie du gène égoïste, conflits génétiques, sélection fréquence-dépendante.
- ↑ Stanislas Lyonnet, Dominique Stoppa-Lyonnet, Les 100 mots de la génétique, Presses universitaires de France, (lire en ligne), p. 65.
- ↑ Charles Darwin (trad. Edmond Barbier), L'Origine des espèces, Paris, Librairie C. Reinwald, Schleicher Frères éditeur, , 604 p., p. 67. L’Origine des espèces, Paris, Librairie C. Reinwald, Schleicher Frères éditeurs,
 (Wikisource).
(Wikisource). - ↑ Jean-Louis de Lanessan, La lutte pour l'existence et l'évolution des sociétés, Paris, Felix Alcan, , 277 p. (lire en ligne), page 15.
- ↑ Aristote Physique II, La nature, chapitre VIII p.81 dans l'édition électronique dirigée par Laurence Hansens-Love Profil Textes philosophiques, édition 1990.
- ↑ Cf. Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, Cambridge, Harvard University Press, 1982 — traduit en français sous le titre de Histoire de la biologie. Diversité, évolution et hérédité, Fayard (1989) : 894 p. (ISBN 2213018944).
- ↑ Thierry Lodé, La Guerre des sexes chez les animaux, une histoire naturelle de la sexualité. 2006, éd. Odile Jacob, Paris, (ISBN 2-7381-1901-8).
- ↑ Cf. André Pichot, Histoire de la notion de vie, éd. Gallimard, coll. TEL, 1993.
- ↑ (en) Genetic Signatures of Strong Recent Positive Selection at the Lactase Gene. Todd Bersaglieri, Pardis C. Sabeti, Nick Patterson, Trisha Vanderploeg, Steve F. Schaffner, Jared A. Drake, Matthew Rhodes, David E. Reich, and Joel N. Hirschhorn. The American Journal of Human Genetics, volume 74 (2004), p. 1111–1120.
- ↑ (en) [PDF] Convergent adaptation of human lactase persistence in Africa and Europe. Sarah A. Tishkoff, Floyd A. Reed, Alessia Ranciaro, Benjamin F. Voight, Courtney C. Babbitt, Jesse S. Silverman, Kweli Powell, Holly M. Mortensen, Jibril B. Hirbo, Maha Osman, Muntaser Ibrahim, Sabah A. Omar, Godfrey Lema, Thomas B. Nyambo, Jilur Ghori, Suzannah Bumpstead, Jonathan K. Pritchard, Gregory A. Wray & Panos Deloukas. Nature Genetics, volume 39 (January 2007), p. 31-40.
- ↑ (en) « Egg mimicry by cuckoos Cuculus canorus in relation to discrimination by hosts. M. de L. Brooke & N. B. Davies. Nature 335, 630-632 », sur nature.com (consulté le ).
- ↑ (en) « Arms Races between and within Species. R Dawkins, JR Krebs. Proceedings of the Royal Society of London. Series B (1979) 205, 1161 », sur journals.royalsoc.ac.uk (consulté le ).
- ↑ Brennan PL, Prum RO, McCracken KG, Sorenson MD, Wilson RE, et al. (2007) Coevolution of Male and Female Genital Morphology in Waterfowl. PLoS ONE 2(5): e418. doi:10.1371/journal.pone.0000418
- ↑ Arnqvist G, Rowe L. (2002) Antagonistic coevolution between the sexes in a group of insects. Nature 415 : 787–789.
- ↑ Théophile Godfraind, Hominisation et transhumanisme, Académie royale de Belgique, , p. 121.
- ↑ (en) M Arendt, K M Cairns, J W O Ballard, P Savolainen & E Axelsson, « Diet adaptation in dog reflects spread of prehistoric agriculture », Heredity, vol. 117, no 5, , p. 301–306 (DOI 10.1038/hdy.2016.48).
- ↑ Pierre Robert, « Des éléphants sans défenses : quand l'homme provoque la sélection naturelle », sur franceculture.fr, .
- ↑ Robert E Ricklefs, Rick Relyea, Écologie. L'économie de la nature, De Boeck Superieur, , p. 166.
- ↑ http://www.genetics.org/content/121/3/527.full.pdf.
- ↑ E. Danchin, L.-A. Giraldeau & F. Cézilly (2005). Écologie comportementale. Dunod.
- ↑ Picq P. L'humain à l'aube de l'humanité p. 41 dans Qu'est-ce que l'humain ?.
- ↑ (en) « Wilson, D. S., & Sober, E. (1994). Reintroducing group selection to the human behavioral sciences. Behav. Brain Sci. 17 (4): 585-654 », sur bbsonline.org (consulté le ).
- ↑ Boyd et Silk, L'Aventure humaine, de la molécule à la culture, De Boeck 2004, partie 1, chapitre 3, p. 68-72 ; (ISBN 2 8041 4333 3).
- ↑ Laland et Coolen, la culture autre moteur de l'évolution, dans La Recherche n° 377 juillet-août 2004.
- ↑ (en) Vidéo d'une conférence sur la relation entre biologie et sciences humaines
- ↑ (en) Site des auteurs de la théorie de la construction de niche
- ↑ (en) Life's Solution Inevitable Humans in a Lonely Universe, Simon Conway Morris, Cambridge University Press, New York, 2003. 486 p. 30 $, 18,95 $. (ISBN 0-521-82704-3).
- ↑ [PDF] On meme--gene coevolution. Bull L, Holland O, Blackmore S. Artif Life. 2000 Summer ; 6(3):227-35. PDF.
- ↑ (en) Atran, S. (2001) The Trouble with Memes : Inference versus Imitation in Cultural Creation. Revue scientifique. Human Nature 12(4):351-381.
Annexes
Articles connexes
- Accommodation génétique
- Anthropologie
- Élevage sélectif des animaux
- Exploitation anthropique : effets sur la sélection naturelle
- Historique des critiques des théories de l'évolution
- Modes de sélection naturelle
- Sélection (biologie), radiation évolutive
- Sélection et adaptation durant les successions végétales
- Sélection fluctuante
- Sélection fréquence-dépendante et Phénomène densité-dépendant
- Sélection température-dépendante
Bibliographie
- Serres M., Picq P., Vincent J.-D. (2003) Qu’est-ce que l’Humain ?, éd. Le Pommier et le Collège de la Cité des sciences (ISBN 2746501309).
- Chayeux J.-P. (2003) Gènes et cultures - Symposium annuel, éd. Odile Jacob, 304 pages (ISBN 2-7381-1310-9).
- Mousseau, T. A., et D. A. Roff. 1987 Natural selection and the heritability of fitness components. Heredity 59: 181–197
- Patrick Tort, Darwin et la science de l’évolution, Gallimard, coll. « Découvertes Gallimard / Sciences et techniques » (no 397), 2000, 160 p.
- Charles Darwin, La Filiation de l’Homme et la sélection liée au sexe, trad. sous la direction de P. Tort, coord. par M. Prum. Précédé de Patrick Tort, « L’anthropologie inattendue de Charles Darwin ». Paris, Champion Classiques, 2013.
- Charles Darwin, La Variation des animaux et des plantes à l’état domestique, trad. sous la direction de P. Tort, coord. par M. Prum. Précédé de Patrick Tort, « L’épistémologie implicite de Charles Darwin ». Paris, Champion Classiques, 2015.
- Patrick Tort, L'Effet Darwin. Sélection naturelle et naissance de la civilisation, Paris, Seuil, « Science ouverte », 2008, 234 p. Réédition Points-Sciences, 2017.
Liens externes
- Ressource relative à la recherche :
- Ressource relative à la musique :
- Ressource relative à la santé :
- (cs + sk) WikiSkripta
- Notices dans des dictionnaires ou encyclopédies généralistes :