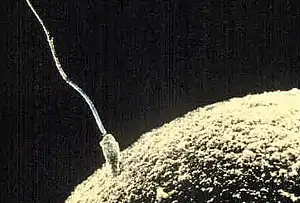
La fécondation est l'étape de la reproduction sexuée où les gamètes, en général un gamète mâle et un gamète femelle, apportent leurs chromosomes et le reste de sa machinerie moléculaire à une cellule unique nommée zygote. Elle a été observée et décrite pour la première fois par Gustave Adolphe Thuret en 1854 chez l'algue brune Fucus. En 1876, Oscar Hertwig publie un article démontrant que la fécondation est l'union du noyau d'un gamète mâle avec un gamète femelle. Il avait alors travaillé sur l'oursin.
La fécondation permet le passage de deux cellules haploïdes, c'est-à-dire les gamètes, en une cellule diploïde qui est le zygote.
Fécondation chez les plantes vertes
Le processus complet de la fécondation est encore mal compris.
Ce n'est que récemment qu'on a compris la manière dont le tube pollinique était chimiquement guidé vers l'ovule[1] en s'enfonçant dans les tissus de reproduction féminin avant de cesser sa croissance, se casser pour libérer deux spermatozoïdes, l'un fécondant l'ovule et l'autre fusionnant avec une cellule femelle pour générer l'endosperme, le tissu de nutrition de l'embryon végétal. Ce n'est que récemment qu'on a identifié le mode de signalisation permettant aux cellules mâles et femelles de se reconnaître et produire l'embryon. Au début des années 2000, Thomas Dresselhaus et ses collègues à l'université de Ratisbonne (Allemagne) trouvaient quatre protéines défensines exclusivement exprimées dans le sac embryonnaire, ce type de protéines étant habituellement impliquées dans le système immunitaire des végétaux, mais aussi des insectes et d'autres animaux[2]. Elles auraient pu être destinées à tuer d'éventuels microbes qui auraient pénétré l'ovaire, mais via d'autres études d'expression des gènes et la création de plantes knock-down, on a montré que l'une de ces quatre protéines (la Defensine-like, ZmES4) est rejetée par le sac embryonnaire au cours du processus de fécondation et provoque la rupture du tube pollinique qui peut alors libérer les spermatozoïdes. La fonction de cette protéine n'était pas la défense, mais la rupture de l'extrémité du tube pollinique ; il pourrait s'agir d'un détournement de fonction originellement de protection immunitaire. La protéine cible du tube pollinique est celle des canaux potassiques. Quand elle est contactée par ZmES4, un flux d'ions potassium et d'eau est brusquement apporté dans les canaux potassiques, rompant l'équilibre osmotique et provoquant une mini-explosion qui libère également les deux spermatozoïdes. La protéine n'agit ainsi que sur le tube de sa propre espèce (La fécondation a même été retardée de 30 min dans le tube pollinique d'une sous-espèce proche de maïs, Tripsacum dactyloides)[2]. Certains généticiens souhaitent mieux comprendre ces mécanismes pour créer des outils biotechnologiques permettant de nouvelles fertilisations croisées voire la création de chimères (ce qui demanderait aussi de résoudre des problèmes d'incompatibilités génomiques). Des outils ou tests de sélection pourraient aussi dériver de cette découverte[2].
Dans le domaine des végétaux, la fécondation se réalise selon deux modalités :
- l'autofécondation (autogamie), ou fécondation par son propre pollen (cas général chez le pêcher). Ce mode de fécondation favorise un taux élevé d'homozygotie. ;
- l'interfécondation (allogamie), ou fécondation croisée (cas général chez le pommier et le poirier), les insectes et particulièrement les abeilles assurant fréquemment la pollinisation. Ce mode de fécondation favorise un taux élevé d'hétérozygotie.
Double fécondation
Chez les angiospermes et les gnétophytes la fécondation est double :
Le grain de pollen produit deux cellules germinales.
- l'une des deux cellules germinales mâle s'associe à l'oosphère. Ceci mène à la formation du zygote-plantule, ou embryon de plante, à l'origine d'une nouvelle plante. Ce zygote est diploïde.
- l'autre fusionne avec les deux noyaux de la cellule centrale et constituent le zygote-albumen, servant de réserve pour la plantule lors de la germination. Ce zygote est triploïde.
Fécondation artificielle
Elle permet de réunir sur un seul individu les qualités possédées par plusieurs. Les soins préalables consistent à enlever, avant la fécondation, toutes les anthères des fleurs qu'on veut féconder puis, quand le stigmate est bien développé, à apporter sur ces fleurs castrées, du pollen d'une variété dont on veut reproduire les caractères.
L'hybride ne sera pas le fruit obtenu mais le fruit issu du semis des graines du fruit obtenu.
Fécondation chez les animaux
Fécondation naturelle
Deux types de fécondation existent chez les animaux : la fécondation interne et la fécondation externe.
Fécondation interne

Lors d'une fécondation interne, le sperme mâle est émis dans les voies génitales femelles, où les spermatozoïdes rencontreront les ovocytes. Ce type de fécondation implique généralement coopération des individus et accouplement.
La fécondation, c'est-à-dire la rencontre des gamètes, peut être facilitée par des comportements sexuels, notamment des parades nuptiales. Chez les espèces à fécondation interne, ces comportements favorisent le rapprochement des deux partenaires et, donc, l'accouplement.
On trouve généralement la fécondation interne chez les espèces vivant en milieu terrestre, principalement chez les mammifères (y compris aquatiques), les oiseaux, les reptiles et les insectes.
Dans le milieu aquatique, la fécondation interne existe chez les poissons cartilagineux (sélaciens comme les requins) et les mollusques céphalopodes.
Fécondation externe
Lors d'une fécondation externe, les gamètes sont émis dans le milieu extérieur.
Il n'y a pas d'accouplement dans ce type de fécondation, et les gamètes libérés vont se rencontrer au gré du hasard dans l'eau où ils vont s'unir. Les gamètes sont émis en très grande quantité, ce qui optimise les probabilités de leur rencontre.
Certaines espèces à fécondation externe présentent aussi des comportements sexuels qui facilitent la rencontre des gamètes. C'est le cas, en particulier, de grenouilles ou de poissons dont la parade nuptiale permet la libération simultanée des gamètes.
La fécondation externe est trouvée généralement chez les espèces vivant en milieu aquatique. Elle est rencontrée principalement chez les batraciens, les poissons osseux ou de nombreux invertébrés aquatiques.
Fécondation artificielle
Observateur et expérimentateur, l'abbé Spallanzani réalise, en 1777, la première fécondation artificielle, en étudiant le mécanisme de la reproduction chez les batraciens. Il réussit, en 1779, la première insémination artificielle d'une chienne.
En 1875, l'embryologiste Oscar Hertwig observe, lors d’une fécondation artificielle d’oursin, la pénétration d’un spermatozoïde dans l’ovule, la fusion des noyaux mâle et femelle et la division de l’œuf en deux cellules.
Fécondation chez l'être humain
Chez les humains, comme chez la plupart des animaux, la fécondation est une des étapes de la reproduction. Elle consiste en la rencontre du gamète mâle, le spermatozoïde avec le gamète femelle, un ovocyte II. Le gamète mâle doit préalablement avoir subi l'étape de capacitation dans les voies génitales femelles et obtenir sa capacité fécondante. La fécondation se déroule en quatre phases distinctes :
- Reconnaissance spécifique
- Le spermatozoïde et l'ovocyte se reconnaissent comme compatibles, de la même espèce. Cette reconnaissance est effectuée entre les protéines composant la zone pellucide (enveloppant l'ovocyte pendant sa maturation) et des récepteurs présents sur la membrane du spermatozoïde. Le spermatozoïde ne subit pas de phénomène de rejet comme corps étranger car il produit à sa surface des cytokines polypeptidiques, des transforming growth factor-bêta (TGFβ2 et TGFβ3) qui agissent comme éléments anti-rejet. Il se produit alors une réaction acrosomique, qui va « dissoudre » la zone pellucide et permettre le passage du noyau de gamète mâle, jusqu'à la membrane plasmique de l'ovocyte. Le spermatozoïde paternel apporte la moitié des chromosomes, leurs molécules épigéniques et un centrosome au zygote. Ce dernier reçoit toutes ses mitochondries, l'autre moitié de ses chromosomes, leurs molécules épigéniques et le reste de sa machinerie moléculaire de son son ovule maternel. Chez l'humain et autres mammifères à fécondation interne, il n'y a a priori pas de problème de reconnaissance, deux espèces différentes ne s'accouplant que rarement ensemble. Les expériences ont tout de même montré qu'une fécondation entre deux espèces différentes n'était pas possible, du fait de la différence des génomes entre les espèces. Ce mécanisme de reconnaissance spécifique est surtout utile pour les animaux à fécondation externe, comme certains poissons ou amphibiens : la femelle pond ses œufs dans le milieu, et le mâle vient y déposer son sperme.
- Fusion du noyau du spermatozoïde et de l'ovocyte
- Afin de garder une quantité 2n de matériel génétique chez le zygote, un seul spermatozoïde doit féconder l'ovocyte : c'est la monospermie. Cette monospermie est premièrement permise quasiment immédiatement par le changement de la polarité électrique de la membrane de l'ovule dès le premier contact avec le spermatozoïde et par la reconnaissance mutuelle de deux protéines spécifiques, Juno et Izumo[3] ; elle est ensuite permise grâce au réveil ovocytaire qu'entraine la fusion des gamètes (caryogamie), qui est une réaction plus lente (une dizaine de minutes environ). Ainsi les granules corticaux (lysosomes synthétisés durant la croissance de l'ovocyte) sont exocytés sous le contrôle d'une augmentation de la concentration en calcium cytosolique et leurs contenus enzymatiques modifieront les glycoprotéines de la zone pellucide qui deviendra "imperméable" à d'autres spermatozoïdes.
- Reprise de la méiose pour l'ovocyte
- celui-ci était bloqué en métaphase II avant la fécondation. Il finit donc sa deuxième division de méiose et expulse son deuxième globule polaire. Cette activation de l'ovocyte est sous le contrôle du calcium cytosolique dont la concentration augmente grâce à une enzyme (une phospholipase C, isoforme zeta) apportée par le spermatozoïde[4]. Une fois cette étape terminée, on trouve dans l'ovocyte deux noyaux, appelés pronuclei : la pronucleus femelle et le pronucleus mâle (provenant du spermatozoïde). On peut alors parler d'ovule et non plus d'ovocyte.
Amphimixie et déclenchement du développement embryonnaire
- il s'agit de la fusion des deux pronuclei. En réalité, les deux pronucléi ne se fusionnent pas à proprement parler, comme on pourrait l'imaginer, mais le matériel génétique se rassemble sur la plaque équatoriale au moment de la métaphase de la toute première division cellulaire du nouveau zygote.
Fécondation in vitro
Jusqu'au XXe siècle, la fécondation, la fusion des gamètes, avait nécessairement lieu dans le corps de la femme. Mais en 1978, naît Louise Brown, le premier nouveau-né obtenu par fécondation in vitro, donc par une fécondation hors du corps de la femme.
Le principe de base est simple : un prélèvement de sperme de l'homme et un ovocyte II de la femme sont mis en contact dans une éprouvette, et un œuf se forme. L'embryon obtenu est alors transféré dans l'utérus de la femme.
En fait, pour augmenter les chances d'avoir un embryon, il faut employer plusieurs ovules. Les ovaires de la femme sont sur-stimulés pour obtenir une dizaine d'ovules. Le sperme de l'homme est mis en contact avec tous ces ovules, ce qui permet d'obtenir cinq ou six embryons. Deux ou trois d'entre eux sont transférés dans le corps de la femme, alors que les autres sont congelés si la division cellulaire le permet. Ils pourront être utilisés pour une autre tentative, si les parents le désirent, ou bien être détruits.
Histoire
- Les Grecs utilisaient la fécondation artificielle du dattier au Ve siècle av. J.-C.
- L'insémination artificielle était pratiquée par les Arabes pour la reproduction des chevaux depuis le XIVe siècle.
- Le procédé de fécondation artificielle de la vanille par Edmond Albius en 1841 a permis à l'Empire colonial français de rompre le monopole de production de la vanille dont jouissait jusque-là le Mexique[5].
Notes et références
- ↑ Identification de deux protéines clés dans le processus de fécondation des plantes à fleur (Source : Ambassade de France au Japon/ ADIT, BE Japon 501 du 18/05/2009)
- 1 2 3 News de Megan Scudellari, à propos de S. Amien et al. Defensin-Like ZmES4 Mediates Pollen Tube Burst in Maize via Opening of the Potassium Channel KZM1 ; PLoS Biol 8(6): e1000388
- ↑ Article relatant la découverte sur Nature.com lire en ligne
- ↑ (en) PLCzeta, a sperm-specific PLC and its potential role in fertilization. - U.S. Department of Health & Human Services, National Center for Biotechnology Information (NCBI), 2007
- ↑ Eric Jennings, « La vanille à la conquête du monde », L'Histoire, no 494, .
Articles connexes
- Caryogamie