Le système immunitaire[1] d'un organisme est un système biologique complexe constitué d'un ensemble coordonné d'éléments de reconnaissance et de défense qui discrimine le soi du non-soi. Ce qui est reconnu comme non-soi est détruit.
Il protège l'organisme des agents pathogènes : virus, bactéries, parasites, certaines particules ou molécules « étrangères » (dont certains poisons), mais est responsable du phénomène de rejet de greffe[2].
Il est hérité à la naissance, mais autonome, adaptatif et doué d'une grande plasticité, il évolue ensuite au gré des contacts qu'il a avec des microbes ou substances environnementales étrangères au corps[3].
On dénombre plusieurs types de systèmes immunitaires parmi les espèces animales, et généralement plusieurs mécanismes immunitaires collaborent au sein d'un même organisme.
Pour de nombreuses espèces, dont les mammifères, le système est constitué de 3 couches. Ses principaux effecteurs sont les cellules immunitaires appelées leucocytes (ou globules blancs) produites par des cellules souches, au sein de la moelle osseuse rouge. Cette séparation en trois couches n’empêche pas une interaction très importante des couches entre elles:
- La barrière épithéliale comme la protection de la peau et les muqueuses, l'acidité gastrique ;
- Les mécanismes de défense non spécifique ou innée ou naturelle dérivant des cellules immunitaire de la lignée myélocytaire .
- Les mécanismes de défense spécifique ou adaptative dérivant des cellules immunitaires de la lignée lymphocytaire. L' immunité humorale s'oppose aux agents pathogènes extracellulaires grâce à des anticorps spécifiques, protéines sécrétés grâce aux lymphocytes B. L'immunité cellulaire s'oppose aux pathogènes intracellulaires, pris en charge par les lymphocytes T . Cette dernière couche n’existe que chez les vertébrés.
On appelle réponse immunitaire l'activation des mécanismes du système immunitaire face à la reconnaissance de non-soi, agressive ou pas, face à une agression ou à une dysfonction de l'organisme.
L'ensemble de ces systèmes (y compris lors de la vaccination) permet la résilience immunitaire, notion qui recouvre la somme des mécanismes efficaces de défense d’un organisme vis-à-vis d’un agent pathogène (du grec pathos : souffrance) ; il se dégrade avec l'âge (Immunosénescence).
Le système peut entraîner un dysfonctionnement autoimmune.
Cellules du système immunitaire
Toutes les cellules du système immunitaire dérivent d'une cellule souche présente dans la moelle osseuse. Cette cellule souche donne deux lignées de cellule : la lignée lymphocytaire et la lignée myélocytaire.
Les cellules de l'immunité innée sont produites par la lignée myélocytaire. Les cellules de l'immunité adaptative sont produites par la lignée lymphocytaire.
Un seul type de cellule est produite par les deux lignées : la cellule dendritique.

Lignée lymphocytaire
La cellule souche multipotente donne le progéniteur lymphoïde (lymphoid progenitor) qui se divise en trois types de cellules :
- le progéniteur cellulaire B (B cell progenitor). Ces cellules se différencient en cellule à mémoire B (Memory B cell) et en plasmocyte qui secrète des anticorps. Ces cellules sont un composant majeur du système immunitaire adaptatif ;
- le progéniteur cellulaire T (T cell progenitor) donne naissance aux cellules à mémoire T, aux lymphocytes T cytotoxiques et aux lymphocytes T auxiliaires. Les cellules à mémoire T ont les mêmes fonctions que les cellules à mémoire B. Les cellules cytotoxiques reconnaissent les antigènes des agents pathogènes et les détruisent. Les lymphocytes T auxiliaires secrètent des cytokines qui stimulent les cellules de la lignée B ;
- les cellules lymphocytaires tueuses (ou lymphocytes NK, sigle de l'anglais Natural Killer, signifiant « tueur naturel ») sont les cellules tueuses du système immunitaire inné. Elles détectent les cellules infectées par les virus et les détruisent ;
- la lignée lymphocytaire produit aussi des cellules dendritiques.
Lignée myélocytaire
Responsable de la production des hématies et plaquettes, cette lignée donne naissance à des cellules impliquées dans le système immunitaire inné et dans le système immunitaire adaptatif en produisant des cellules portant les antigènes des agents pathogènes pour les présenter aux cellules du système immunitaire adaptatif :
- les neutrophiles sont des cellules du système immunitaire inné s’attaquant aux agents pathogènes et les détruisant. Cette destruction peut aboutir à la destruction de la cellule elle-même aboutissant à la formation de pus ;
- les éosinophiles libèrent des cytokines en présence de parasite ;
- les monocytes vont se différencier en macrophages et en cellules dendritiques. La cellule dendritique permet une communication entre les cellules responsables de l'immunité innée et les cellules responsables de l'immunité adaptative. Les macrophages sont des cellules « poubelles » ;
- un dernier type de cellules impliquées dans les réactions allergiques, les cellules mastocytaires et les basophiles.
Mécanismes de défense
L'organisme se défend contre les dysfonctions de ses cellules et les agressions, c'est-à-dire des processus qui ont pour conséquence de détruire des êtres vivants. Ces agressions peuvent revêtir différentes formes :
- des agressions dites physico-chimiques :
- mécaniques (frottements, chocs, chutes, etc.),
- facteurs climatiques (altitude, changement brusque de température, rayonnements, sécheresse, poussières, etc.),
- agression par des agents chimiques (acides, bases, etc.) ou autres éléments (aluminium, notamment) ;
- des agressions par d'autres êtres vivants : un organisme constitue pour un autre organisme un endroit idéal pour le développement de ses propres cellules et aussi pour un certain nombre de microorganismes qui pourraient y proliférer, il s'agit alors d'une infection. Les agresseurs dans ce cas peuvent être : des virus, des bactéries, des champignons, des levures, des helminthes ou des prions.
 Les cellules du système immunitaire selon leur appartenance aux deux types de réponses immunitaires.
Les cellules du système immunitaire selon leur appartenance aux deux types de réponses immunitaires.
Mécanismes de défense non spécifique (innée)
Le système inné est un mécanisme très rapide de défense aux infections: il permet souvent d'arrêter un agent pathogène ou, du moins, de permettre la mise en route du système adaptatif qui a des armes plus puissantes et plus spécifiques pour arrêter l'agent pathogène. Il a longtemps été considéré comme un système non spécifique, mais la découverte de récepteurs cellulaires spécifiques de plusieurs familles de pathogènes (comme les bactéries gram-négatives) dans les années 2000 a changé notre regard sur le système inné.
Barrières physiques

- La peau est le premier, le plus grand et le plus important élément du système de défense : il empêche l'entrée de la plupart des corps étrangers. Les épithéliums cutanés secrètent des peptides anti-microbiens dès la pénétration d'un agent pathogène. Ils secrètent aussi des cytokines et des chimiokines.
- Les personnes qui perdent trop de peau, par brûlure par exemple, risquent de succomber à des infections. Pour éviter cela, elles sont placées dans des chambres hospitalières maintenues aussi stériles que possible.
- Les muqueuses (ou tissus continus) qui recouvrent les voies oropharyngiennes et digestives, ainsi que les voies respiratoires et urogénitales, constituent également une barrière physique. Les cellules très étroitement juxtaposées sont imperméables à la plupart des agents infectieux. En surface de certaines muqueuses, un film de mucus animé par les battements de cils vibratiles permet de fixer, enrober puis évacuer vers l'extérieur la plupart des particules ou êtres vivants intrus.
- Les cellules possèdent divers systèmes « passifs » de défense chimique et biochimique (enzymes, acides gras, acide lactique, pH du corps, etc.).
- La peau et les muqueuses sont recouvertes d'une flore bactérienne dite « normale » qui protège, souvent par concurrence, des micro-organismes pathogènes.
Système de défense inné
Le système immunitaire inné est déclenché par des récepteurs cellulaires reconnaissant des structures moléculaires uniques aux agents pathogènes ou par des molécules signifiant les dégâts.
Cellules effectrices de l'immunité innée
Elles dérivent toutes de la lignée myélocytaire de l'hématopoïèse. Elles sont parfois regroupées sous le terme de leucocytes phagocytaires ou phagocytes. Ce terme est très réducteur car il laisse à penser que la seule fonction de ces cellules est la phagocytose, alors qu'elles ont d'autres fonctions essentielles.
Ce sont des cellules immunitaires qui reconnaissent les microorganismes grâce à de nombreux récepteurs cellulaires présents à leur surface. Ces récepteurs permettent aux phagocytes de reconnaître certaines structures présentes à la surface des microorganismes infectieux et d'internaliser ces derniers à l'aide d'une vacuole digestive. Par la suite, ils fusionnent la vacuole contenant les microbes avec un lysosome. Les lysosomes peuvent contenir des formes toxique d'oxygène comme du monoxyde d'azote (NO) ou du peroxyde d'hydrogène (H2O2), et ils peuvent aussi contenir du lysozyme et d'autres enzymes digestives qui dégradent des structures microbiennes[4].
Cellules résidentes dans le tissu sous épithéliales
Les cellules résidentes sont les premières activées en cas de franchissement de la barrière épithéliale (cutanée, respiratoire ou intestinales) par un microbe.
Macrophage
Les macrophages ont une plus grande capacité de phagocytose que les granulocytes neutrophiles, et, lorsqu'ils phagocytent un microorganisme, des voies cellulaires internes les stimulent, ce qui les rend plus efficaces dans leurs tâches.
Mastocyte
Ces cellules contiennent des granules contenant des substances vasodilatatrices comme l'histamine. Cette substance en vasodilatant le vaisseau entraîne une diminution de la vitesse de circulation du sang permettant au leucocyte neutrophile de traverser la paroi vasculaire.
Cellule dendritique
Les cellules dendritiques qui dérivent aussi des monocytes sont des cellules présentatrices d'antigènes. Leur rôle est de capturer un microbe au site d'infection, de migrer vers les tissus lymphoïdes et de présenter les antigènes du microbe aux lymphocytes T à l'aide d'une molécule de CMH. Ce type de molécule joue un rôle très important dans la réaction immunitaire primaire[4].
Cellules présentes dans le sang
Leucocyte neutrophile
Les granulocytes neutrophiles représentent 60 à 70 % des leucocytes. Ils pénètrent dans les tissus infectés pour phagocyter les microbes présents et les détruire. Généralement, les granulocytes neutrophiles s'autodétruisent en même temps qu'ils détruisent les microbes. Ils ont normalement une espérance de vie de seulement quelques jours. Ce sont des cellules présentes dans le sang et capables de migrer vers un site où se produit une infection.
Leucocyte éosinophile
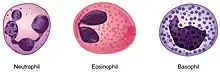
Les granulocytes éosinophiles sont présents en très petite quantité dans l'organisme. Ils ont une faible capacité de phagocytose, mais ils sont essentiels dans le combat contre les parasites présents dans l'organisme. Ils se lient à la paroi du parasite et libèrent des enzymes qui vont causer des dommages importants à celui-ci.
Leucocyte basophile
Les leucocytes basophiles sont les plus rares des leucocytes. Leur taux est si faible que l'absence de leucocyte basophile au cours d'une numération formule sanguine ne doit pas être considérée comme anormale.
Monocyte
Les monocytes représentent 5 % des leucocytes. Ils circulent dans le sang et migrent vers un tissu où ils se transformeront par la suite en macrophages[5]. Les macrophages et les cellules dendritiques sont des cellules résidentes dans les tissus sous-épithéliaux.
Molécules de l'immunité innée
Il existe quatre grands groupes de molécules intervenant dans l'immunité innée : les peptides anti-microbiens, le système du complément, l'interféron I alpha et I beta et les protéines de la phase aiguë dont la plus utilisée en pratique médicale est la protéine C reactive.
Mécanisme général du système inné
Première étape ; le franchissement de la barrière épithéliale
L'introduction d'un agent infectieux, comme une bactérie gram négatif, au cours d'une piqûre à travers la peau déclenche dans les minutes qui suivent la libération de peptides anti-microbiens et de cytokines par les cellules de l'épithélium cutané[6].
Deuxième étape ; la stimulation des cellules résidentes
Les cellules résidentes de l'immunité innée (macrophage, mastocyte, cellule dendritique) reconnaissent le pathogène par des récepteurs[7] appelés pattern recognition receptor (PRR) ou en français récepteurs de reconnaissance de motifs moléculaires, dont il existe 4 types principaux. Pour les bactéries gram-négatives, il s'agit d'un récepteur de type Toll[8] (TLR ou Toll Like Receptor). La bactérie contient sur la surface de sa paroi des lipopolysaccharides spécifiques aux bactéries gram-négatives qui sont reconnus par les TLRs. Les structures biochimiques reconnues par le TLR sont appelées motifs moléculaires associés aux pathogènes.
La liaison TLR-PPR va déclencher des événements qui diffèrent selon le type cellulaire. Au niveau des mastocytes, elle entraînera la libération d'histamine, celle-ci déclenchant la dilatation des vaisseaux aboutissant à un ralentissement de la circulation sanguine. Au niveau des macrophages et des cellules dendritiques, elle entraînera la libération des cytokines et des chimiokines; les chimiokines vont attirer les leucocytes une fois que ceux-ci ont traversé l’endothélium de la paroi vasculaire. La liaison TLR-PPR active une voie de signalisation qui va déclencher une synthèse de protéine anti-microbienne.
Troisième étape ; le recrutement des cellules immunitaires sanguines
Le ralentissement du débit sanguin secondaire à la vasodilatation permet aux leukocytes de traverser la paroi. Outre les leucocytes, les facteurs du complément traversent la paroi participant aussi à la réaction du système inné. Au niveau cutané, la manifestation clinique de l'infection se traduit par quatre signes : rougeur, chaleur, douleur et œdème. Ces quatre signes caractérisent la réaction inflammatoire.
Si l'infection n'est pas contenue localement
Si le système inné n'arrive pas à contenir l'infection, la cellule dendritique va se diriger vers un ganglion lymphatique par les canaux lymphatique. Elle va se maturer au cours du voyage. Dans le ganglion, elle présentera à la cellule T CD4 + auxiliaire des petits morceaux de 30 à 40 acides aminés de la bactérie phagocytée. Cette présentation de l’antigène se fait par son complexe majeur histocompatibilité de classe II.
Mécanismes de défense spécifique (adaptative)
L'immunité adaptative repose sur 3 acteurs : les organes lymphoïdes, les lymphocytes B et les lymphocytes T. Ces 3 acteurs vont permettre de reconnaître un agent pathogène, de le signaler et de déclencher soit l'immunité humorale soit l'immunité cellulaire. Que ce soit l'immunité humorale ou l’immunité cellulaire, l'immunité adaptative ne se déclenchera que si cet antigène porte aussi un récepteur cellulaire de pathogénicité montrant bien la complexité et l'interaction des deux immunités.
- Spécificité : Les lymphocytes T ou B sont porteurs de récepteurs spécifiques. Chaque lymphocyte ne va reconnaître qu'un seul agent pathogène. L'ensemble des lymphocytes est appelé le répertoire des lymphocytes. La multiplicité des lymphocytes pour chaque agent pathogène est possible grâce à la recombinaison V(D)J. Cette recombinaison somatique pourrait produire plus de 10 000 000 000 de type de récepteurs différents aboutissant à un répertoire gigantesque.
- Reconnaissance du soi et du non-soi :
- le soi d'un individu est défini par des récepteurs du complexe majeur d'histocompatibilité (CMH, nommé HLA chez l'humain) présents sur la membrane de ses cellules, associés aux fragments peptidiques qu'ils présentent. Il existe deux types de molécule de CMH : le CMH de type I qui est présent chez presque toutes les cellules de l'organisme et le CMH de type II qui est retrouvé principalement chez les cellules dendritiques, les macrophages et les lymphocytes B ;
- le non-soi d'un individu est défini par des récepteurs cellulaires ou toute autre molécule différente du soi qui sont ainsi reconnus comme étrangers par notre organisme. Le non-soi déclenche une réaction immunitaire. La reconnaissance du non-soi se calque sur celle du soi, y compris au sein des liquides circulants extracellulaires, lymphatiques, veineux, artériels mais aussi des différents mucus (voir ci-dessus). Les immunoglobulines portés par les membranes des globules blancs et les immunoglobulines dissoutes se fixent sur les molécules présentes dans ces différents liquides. En l'absence de molécules HLA, ou si cette molécule est inconnue, le système immunitaire la reconnaît comme du non-soi et déclenche la mort programmée ou apoptose. Une des limites d'efficacité et de sensibilité du système immunitaire repose donc sur la spécificité de la distinction entre le soi et le non-soi :
- par exemple, les lymphocytes et les plasmocytes ne pénètrent pas dans certains tissus comme ceux du cerveau ou de la thyroïde, ils ne les reconnaissent donc pas comme du soi. Qu'une inflammation s'y installe, ou que ces cellules entrent à leur contact, et ils les identifieront comme du non-soi, puis ils secréteront des anticorps qui s'y fixeront pour les détruire, ce qui initiera une réaction dite auto-immune qui ira en s'amplifiant,
- de même, si les molécules présentes à la surface des agents pathogènes ou des cellules cancéreuses proviennent du soi ou en sont suffisamment proches, le système immunitaire les considèrera comme du soi et ne déclenchera pas de réaction immunitaire,
- le problème est similaire pour les muqueuses où la frontière entre le soi et le non-soi est très ténue. Des molécules habituellement bien tolérées peuvent donc y devenir allergisantes quand elles pénètrent dans des espaces d'où elles devraient être absentes.
Organes lymphoïdes
Les organes lymphoïdes comprennent le thymus, la moelle osseuse, la rate, les amygdales, l'appendice et les ganglions lymphatiques.
Immunité humorale
Le système immunitaire humoral agit contre les bactéries et les virus avant leur pénétration dans les cellules. Les cellules responsables de la destruction des pathogènes extra-cellulaires sont les lymphocyte B agissant en sécrétant des anticorps.
Lymphocyte B
La production et la maturation des lymphocytes B se fait dans la moelle osseuse.

Les lymphocytes B sont le support de l'immunité humorale. Ils possèdent à leur surface des récepteurs, nommés BCR, B Cell Receptor ou récepteurs des cellules B. Chaque lymphocyte B possédée plusieurs BCRs mais pour un seul agent pathogène. Ce BCR est une immoglobuline membranaire formée de 2 chaînes légères et de 2 chaînes longues. Il existe autant de lymphocytes B que de pathogènes. L'ensemble des lymphocytes B est appelé le répertoire des lymphocytes B. Chaque BCR possède 2 sites de fixations à l'antigène.
Le lymphocyte B avant d'être activé est appelé naïf. L'activation du lymphocyte B par l'intermédiaire des BCR déclenche une expansion clonale du lymphocyte activé, avec production de cellule mémoire, et déclenche à distance des cellules produisant des anticorps. Ces cellules produisant des anticorps sont appelés plasmocytes. L'activation du lymphocyte B par un antigène nécessite l'implication des cellules lymphocytaires T CD4.
Les lymphocytes B sont nommés B car ces lymphocytes ont été découverts chez l'oiseau dans la bourse de Fabricius ; par la suite le « B » fut conservé car c'est l'initiale de bone marrow (l'anglais de moelle osseuse) qui correspond au lieu de maturation de ces cellules à la suite d'une exposition à une interleukine (molécule chimique permettant le clonage des lymphocytes B et leur différenciation) produite par les lymphocytes T4.
Anticorps ou immunoglobulines
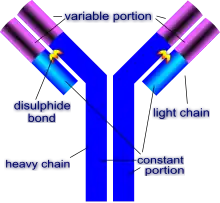
Structure
Ses principaux moyens d'action sont les immunoglobulines, aussi appelées anticorps. Les anticorps sont des molécules ayant une forme de « Y » formées de quatre chaînes polypeptidiques : deux chaînes légères (environ 200 acides aminés chacune) et deux chaînes lourdes (environ 450 acides aminés chacune)[4].
Les chaines légères ont des régions constantes et des régions variables. Les régions variables dépendent de la régulation somatique. Les anticorps ont une forme en Y. La barre verticale du Y est constituée de deux chaînes lourdes constantes qui vont déterminer la fonctionnalité de l’immunoglobuline. Les deux barres inclinées du Y sont formées chacune d'une chaîne lourde et d'une chaîne légère, chacune ayant une partie constante et une partie variable qui est responsable de la spécificité de l'anticorps.
Fonctions
Il existe 5 classes d'anticorps : les IgM, les IgG, les IgA, les IgE et les IgD. Les IgM sont les premiers anticorps à être produits lorsque le corps reconnaît un nouvel antigène. Ceux-ci se retrouvent dans le corps sous forme de pentamère et ils sont très efficaces pour activer le complément. Les IgG sont la classe d'anticorps la plus retrouvée dans le sang, c'est aussi la seule classe d'anticorps qui peut traverser le placenta et donner au fœtus une immunité passive. Les IgA se retrouvent dans les sécrétions (salive, larme, mucus, etc.) sous la forme de dimères. De plus, la présence de ce type d'anticorps dans le lait de la femme permet aux nouveau-nés de recevoir une immunité passive durant la période d'allaitement. Les IgE sont les anticorps impliqués dans les réactions allergiques puisqu'ils provoquent la libération d'histamine et d'autres substances impliquées dans ce genre de réaction par les granulocytes basophiles. Finalement, les IgD sont retrouvés à la surface des lymphocytes B dits « naïfs » (qui n'ont pas encore été exposés à un antigène) et servent de récepteurs cellulaires à ceux-ci. Contrairement aux quatre autres classes d'anticorps, les IgD ont une région transmembranaire qui leur permet de se fixer à la membrane cellulaire des lymphocytes B[4].
Les quatre fonctions principales des anticorps sont :
- la neutralisation et l'agglutination d'antigènes : la neutralisation est le fait d'empêcher les microbes de se lier au cellules de l'hôte ; l'agglutination est le fait de former des agrégats de microbes qui sont facilement phagocytés ;
- l'opsonisation (recouvrement de la cellule cible pour qu'elle soit phagocytée). L'anticorps, après s'être fixé sur un agent pathogène, le présente à un macrophage pour que celui-ci le détruise. Cette reconnaissance par les macrophages se fait par l’intermédiaire de récepteurs FCR qui se lient à la région constante de la chaîne lourde et activent la phagocytose ;
- l'activation du complément ;
- l'élimination d'agents pathogènes trop gros pour être phagocytés, comme un ver. Les anticorps se lient au niveau du ver et activent la libération de substances toxiques par les cellules éosinophiles. C'est la cytotoxicité dépendante des anticorps ou ADCC (Antibody Dependent Cytotoxycity).
Immunité cellulaire
La fonction principale de l'immunité cellulaire est de détruire les agents infectieux intracellulaires. Les cellules responsables de la destruction des pathogènes intra-cellulaires sont les lymphocytes T qui agissent directement en injectant des substances toxiques dans les cellules infectées.
Lymphocyte T
La formation et la maturation des lymphocytes T se fait dans le thymus où le lymphocyte prend le nom de thymocyte. Le lymphocyte T est aussi porteur d'un récepteur pour reconnaître les antigènes pathogènes : les récepteurs des cellules T ou TCR. À la différence des récepteurs des cellules B, le récepteurs des cellules T ne reconnaissent qu'un seul type de molécules : les peptides.
La reconnaissance de la présence d'un agent infectieux intracellulaire par les lymphocyte T se fait par l'intermédiaire du complexe majeur d'histocompatibilité, nommés aussi CMH ou MHC, présent sur les cellules.
Ce complexe majeur d'histocompatibilité a été découvert lors des transplantations d'organes. Ces MHC recueillent en permanence les peptides formés continuellement par la cellule par la dégradation protéique intracellulaire ; elles sont spécifiques à un individu[4].
Les peptides formés en permanence par la cellule par la dégradation protéique intracellulaire et portés par les MHC à l'extérieur de la cellule permettent aux lymphocytes T de vérifier la « santé » de la cellule. En cas d'infection virale, les MHC vont présenter à l'extérieur des peptides viraux qui vont être reconnus par les lymphocytes T. Il en est de même lors d'une greffe d'organe après laquelle les MHC produits seront reconnus comme n’appartenant pas à l'organisme (au soi) risquant de déclencher un rejet de greffe[4].
Le système immunitaire cellulaire s'occupe des cellules infectées par des virus, bactéries, et les cellules cancéreuses. L'action s'effectue via les lymphocytes T. Les lymphocytes T sont capables d'interagir avec les cellules de l'organisme grâce à leurs récepteurs cellulaires T ou TCR (T Cell Receptor) formés de deux chaînes polypeptidiques: la chaîne α (alpha) et la chaîne β (bêta). Ces récepteurs sont tout aussi spécifiques aux antigènes que les anticorps ou que les récepteurs de lymphocytes B, mais, contrairement aux anticorps et aux récepteurs de lymphocytes B, les récepteurs de lymphocytes T ne reconnaissent que de petits antigènes qui doivent être présentés par une molécule de CMH à la surface d'une cellule infectée[4].
Aux lymphocytes T s'ajoutent aussi les lymphocytes NK (natural killer). Ces cellules sont impliquées dans une réponse à mi-chemin entre spécifique et non spécifique, selon les situations. Ils jouent notamment un rôle en début de grossesse, le fœtus devant se protéger contre eux pour pouvoir survivre dans le ventre de sa mère.
Les deux types de complexe majeur d'histocompatibilité

Lorsqu'un agent pathogène pénètre dans une cellule, il reste dans le cytoplasme ou infecte les vacuoles. Les mécanismes pour détruire l'agent différent selon sa localisation et expliquent en partie l'existence de deux familles de MCH, les MCH I et MCH II.
Le complexe majeur d'histocompatibilité de type I
Les MHC I sont produites par les infections cytoplasmiques. Ils activent les lymphocytes T CD 8 qui possèdent le récepteur CD 8. Ces cellules jouent un rôle prédominant dans l'infection virale. Les lymphocytes T CD 8 sont nommés lymphocytes cytotoxiques ou CTL. En effet, la liaison de CD8 sur la molécule de CMH permet de garder le lymphocyte T et la cellule infectée liés plus longtemps, ce qui favorise l'activation du lymphocyte. Une fois activé, le lymphocyte T cytotoxique libère des protéines, comme la perforine ou des granzymes qui provoquent la formation de pores dans la paroi cellulaire de la cellule infectée, entraînant sa mort. Cela a pour effet de priver le pathogène d'un lieu de reproduction et de l'exposer aux anticorps et aux leucocytes phagocytaires qui circulent dans la région infectée[4]. Les MHC I sont présents sur toutes les cellules nuclées de l'organisme. Les hématies ne possèdent donc pas de MHC I.
Le complexe majeur d'histocompatibilité de type II
Les MHC II sont présents sur un nombre très restreint de cellules : cellules dendritiques, macrophages et lymphocytes B.
Les MHC II sont produits par les infections vacuolaires ou la phagocytose. Ils activent les lymphocytes T CD 4 qui possèdent le récepteur CD 4. Les lymphocytes T CD 4 sont nommés lymphocytes helpers ou auxiliaires. En activant les TCD4, ceux-ci libèrent des cytokines transformant les lymphocytes B en plasmocytes sécrétant des immunoglobulines.
Mécanisme général de l'immunité adaptative
Vidéo de la réponse adaptative
Pour que la cellule lymphocytaire B produise des anticorps spécifiques et que le lymphocyte T CD8+ naïf se transforme en lymphocyte T CD8 tueur, il faut deux signaux :
- un signal reçu directement par le lymphocyte B naïf ou le lymphocyte T CD8+ lorsque le microbe se fixe directement sur les récepteurs du lymphocyte B ou T ;
- un signal donné par le lymphocyte T CD4+ helper après activation par la cellule dendritique.
Première étape : la présentation de l’antigène par la cellule dendritique
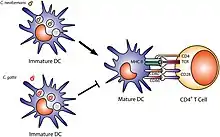
La cellule dendritique est une cellule immunitaire résidant dans le derme ou dans le tissu conjonctif sous-épithélial des bronches ou de l'intestin. Dès l’introduction d'un pathogène, elle va être activée par les molécules émises par les cellules de l'épithélium (les peptides anti-microbiens et les cytokines pro-inflammatoires : l' interleukine-1, l' interleukine-6 et l'interféron-1 alpha et beta).
La cellule dendritique immature possède des récepteurs de reconnaissance de motifs moléculaires qui reconnaissent la famille du microbe porteur d'un motif moléculaire associé aux pathogènes. Elle va internaliser le microbe, le transporter vers un ganglion lymphatique par la lymphe. Au cours du transport, elle va devenir une cellule dendritique mature avec apparition de molécules qui vont lui permettre de se fixer à un lymphocyte T CD4+ auxiliaire naïf.
Durant son transport et dans le ganglion, la cellule dendritique coupe le microbe en morceaux compris entre 30 et 50 acides aminés. Ces morceaux vont être présentés aux lymphocytes T CD4+ auxiliaires grâce au complexe majeur d'histocompatibilité de type II (MCH II). C'est la présentation de l'antigène.
Deuxième étape : l'activation de la cellule T CD4+ auxiliaire par la synapse immunologique
L'activation de la cellule T CD4+ se fait grâce à la synapse immunologique. Des molécules d'adhésion immobilisent la cellule dendritique au lymphocyte TCD4+. La cellule dendritique présente un peptide à la cellule T CD4+. La protéine CD4 se fixe sur un domaine du MCH II. Enfin, des co-récepteurs sont produits par la cellule dendritique après stimulation des récepteurs de reconnaissance de motifs moléculaires. L'ensemble représente la synapse immunologique : la cellule T CD4+ est avertie d'une espèce particulière de microbe par le MCH II de la cellule dendritique à travers les récepteurs de reconnaissance de motifs moléculaires de cette cellule dendritique.
En fonction du signal de famille de pathogène donné par la cellule dendritique lors de la présentation de l'antigène, des cytokines de types différentes vont être émises par la cellule dendritique et activent de façon spécifique les lymphocytes T CD4+ notamment en Th1, Th2. Chaque groupe est spécialisé d'une famille de pathogènes (virus, ver, bactérie, etc.).
Troisième étape : l’activation des lymphocyte B par la reconnaissance de l’antigène ayant activé la cellule T CD4+ auxiliaire
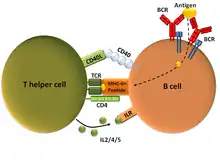
Le même microbe qui a été reconnu par la cellule dendritique se fixe sur les récepteurs des cellules B. Cette fixation va entraîner une activation et un processus aboutissant à la présentation de peptides microbiens par les complexes majeurs d'histocompatibilité de type II (MCH II) au récepteur du T CD4+ : c'est le premier signal.
Le lymphocyte T CD4+ va reconnaître que ce peptide est le même que celui présenté par la cellule dendritique : c'est le deuxième signal.
En fonction du type de CD4+ (Th1, Th2), le TCD 4+ va synthétiser des cytokines, principalement des interleukines, qui à leur tour vont déterminer le type d’anticorps sécrétés. Le lymphocyte B naïf se transforme en lymphocyte B activé. Il va commencer à produire des anticorps A, G ou E. Ces anticorps vont rejoindre le site de l'infection par les canaux lymphatiques aboutissant au canal thoracique se jetant dans l'aorte et vont atteindre le site de l'infection. Un groupe de lymphocytes à mémoire est aussi créé.
Quatrième étape : l’activation des lymphocyte T CD8+ tueurs par la cellule T CD4+ auxiliaire
Tableau de synthèse (immunité adaptative)
| Localisation de l'infection | Substance reconnue par leur récepteur | Aide pour reconnaître la substance | Comment le lymphocyte connaît l'infection | Comment le lymphocyte réagit | Aide pour la réaction | Comment s'appelle le lymphocyte qui réagit ? | |
|---|---|---|---|---|---|---|---|
| Lymphocyte B | Extracellulaire | Toutes les substances | Non | Par la cellule dendritique | En sécrétant des anticorps dans le sang | Oui, par les lymphocytes T nommés lymphocytes T helper ou TCD4 | Plasmocyte |
| Lymphocyte T | Intracellulaire | Uniquement les peptides, des substances très petites | La substance doit être portée par un MHC | Par la cellule dendritique | En sécrétant des substances toxiques dans la cellule infectée | Oui, par les lymphocytes T nommés lymphocytes T helper ou TCD4 | Lymphocyte toxique |
Résumé du fonctionnement d'une réaction de l'organisme face à une invasion bactérienne ou virale
Mécanismes de défense non spécifique (innée)
Première étape. Libération des molécules anti-microbiennes du système inné
L'introduction d'un agent infectieux, comme un virus, déclenche dans les minutes qui suivent la libération de peptides anti-microbiens et de cytokines par les cellules de l'épithélium cutané.
Deuxième étape. Mobilisation des cellules présentes dans les vaisseaux sanguins
Les molécules citées ci-dessus vont aller se fixer sur les récepteurs PRR des macrophages, des mastocytes et de la cellule dendritique. Les mastocytes vont relâcher des granules d'histamine qui ont des capacités vasodilatatrices, ce qui va dilater les parois des vaisseaux sanguins et donc ralentir la vitesse de la circulation sanguine pour permettre au granulocytes de traverser la paroi vasculaire. Les macrophages eux, vont relâcher des chimiokines qui vont se fixer sur les récepteurs PRR des granulocytes et vont les attirer dans le vaisseau sanguin. Les cellules dendritiques vont capturer un microbe et migrer jusqu'aux vaisseaux lymphatiques et aux ganglions, où ils vont présenter une molécule MHC II au lymphocyte T4.
Troisième étape. Mise en action des granulocytes présents dans les tissus
Grâce à la dilatation des vaisseaux produite par les mastocytes et grâce aux chimiokines, les granulocytes présents dans les tissus vont traverser la paroi vasculaire. Les granulocytes neutrophiles vont phagocyter, les ganulocytes basophiles vont relâcher de l'histamine, qui va déclencher la réaction inflammatoire, et les granulocytes éosinophiles vont se lier à la paroi du parasite et libérer des enzymes qui vont causer des dommages importants à celui-ci.
Mécanismes de défense spécifique (adaptative)
Première étape. Présentation la molécule MHC II au lymphocyte T4
Les cellules dendritiques capturent un microbe et migrent jusqu'aux vaisseaux lymphatiques et aux ganglions, où ils vont présenter une molécule MHC II au lymphocyte T4.
Deuxième étape. Activation des lymphocytes B par les lymphocytes T4
Si l'inflammation n'est pas contrôlée par le système inné, les lymphocyte T4 vont activer les lymphocytes B spécifiques au microbe à l'aide d'une cytokine, l'interleukine 2. Les lymphocytes B vont alors produire des anticorps.
Troisième étape. Activation des lymphocytes T8
Les peptides anti-microbiens qui sont libérés en premier par les cellules de l'épithélium cutané vont se fixer aux récepteurs du lymphocyte qui va alors s'activer. Il va libérer tout d'abord de la perforine, une protéine qui va créer des pores dans la paroi des cellules infectées. Il va ensuite libérer du granzime, une protéase à sérine qui va pénétrer par ces pores et induire l'apoptose (mort de la cellule).
Quatrième étape. Neutralisation du microbe
Les anticorps vont se fixer sur les antigènes de la bactéries ou de virus. Cela s'appelle l'opsonisation. Ensuite les anticorps présentent le microbe aux macrophages. Les macrophages activent la phagocytose grâce aux récepteurs FCR.
Mémoire immunologique
Chaque individu acquiert en vieillissant une « mémoire immunologique »[3]. Elle conserve un certain temps les traces de « lutte » passée contre des pathogènes ou parasites, et des cellules spécifiques, permettant une réaction immunitaire plus rapide et efficace. Cette mémoire se constitue de manière naturelle, ou à l'aide de vaccins mais semble se dégrader avec l'âge (phénomène d'immunosénescence).
En effet, l'exposition antérieure à un antigène modifie la vitesse, la durée et l'intensité de la réaction immunitaire. La réaction immunitaire première consiste en la production de cellules effectrices des lymphocytes lors d'une première exposition à l'antigène. Lors d'une seconde exposition au même antigène, la réaction immunitaire secondaire sera plus rapide et efficace car l'organisme aura conservé en mémoire certains lymphocytes de la première attaque.
C'est le principe de la vaccination : on injecte un antigène à la personne pour qu'elle se crée une « mémoire humorale », qui sera directement efficace lors d'une éventuelle attaque ultérieure.
Une étude en 2015[3], basée sur la comparaison de la santé de « vrais » et « faux » jumeaux (210 jumeaux au total, de 8 à 82 ans, suivis pour plus de 200 paramètres de leur système immunitaire, ce qui est une première en nombre de paramètres d'intérêt immunologique), confirme qu'après la naissance, l'environnement a plus d'effets que nos gènes sur le fonctionnement et l'efficacité de notre immunité, notamment via l'exposition antérieure de l'organisme à des agents pathogènes (et/ou à des vaccins)[3]. Les réponses différentes des vrais jumeaux à la vaccination anti-grippale montrent aussi que les réactions (production d'anticorps) ne dépendent pratiquement pas des traits génétiques mais presque entièrement de l'éducation immunitaire de chacun, et donc de nos relations antérieures à l'environnement microbien et parasitaire (dans ce cas liées à des contacts précédents avec diverses souches du virus de la grippe[3]). Face au cytomégalovirus, qui sommeille dans une fraction importante de la population humaine (ne causant que rarement des symptômes), les conclusions sont les mêmes[3].
La meilleure compréhension des mécanismes globaux de l'immunité pourrait peut-être à l'avenir permettre de réduire les problèmes de rejet de greffe car la compatibilité entre un receveur et un donneur ne provient pas que de l'ADN, mais aussi d'enzymes et de facteurs d'immunité qu'on commence à rechercher dans le domaine de la biologie adaptative (via l'immunoséquençage notamment[9],[10],[11]). À l'échelle d'une vie, l'évolution du système immunitaire peut être comparée aux mécanismes complexes en jeu à d'autres échelles dans l'évolution adaptative. De même, des vaccins plus personnalisés pourraient être imaginés.
Maladies du système immunitaire
Le système immunitaire peut se dégrader en réagissant excessivement ou insuffisamment.
Maladie aiguë
L'absence de régulation du système inné peut aboutir au choc cytokinitique.
Maladie chronique
S'il s'attaque aux cellules de l'organisme qui ne sont pas pathologiques (par mauvaise reconnaissance), il va alors se créer une maladie auto-immune qui va se caractériser par une inflammation continue de certains tissus ou par la nécrose complète de certains tissus (par exemple le diabète de type I).
S'il y a un défaut du système immunitaire, dans ce cas les pathogènes ou les cancers pourront se développer plus aisément.
Maladie génétique
Notons l'existence d'une maladie impliquant le système immunitaire adaptatif. Il s'agit du Bare Lymphocytes Syndrome (BLS). Les patients souffrant de cette maladie ne peuvent présenter d'antigène à la surface des cellules présentatrices d'antigène et il ne peut donc pas y avoir production d'anticorps. Cette maladie a notamment permis des avancées en biologie moléculaire en permettant l'identification par complémentation d'un facteur de transcription essentiel, le transactivateur de classe II (CIITA).
Notes et références
- ↑ Le mot « immunité » vient du latin : immunitas (« exempté de charge »).
- ↑ Les receveurs de greffon subissent un traitement immunosuppresseur préalable pour augmenter les chances de succès.
- 1 2 3 4 5 6 Conover E (2015) "Environment, more than genetics, shapes immune system", American Association for the Advancement of Science ; Latest news, publié 15 janvier 2015
- 1 2 3 4 5 6 7 8 Neil A. Campbell, Biologie, Québec, ERPI, , 1334 p. (ISBN 978-2-7613-1783-2 et 2-7613-1783-1), p. 977-991
- ↑ (en) Jiyeon Yang, Lixiao Zhang, Caijia Yu et Xiao-Feng Yang, « Monocyte and macrophage differentiation: circulation inflammatory monocyte as biomarker for inflammatory diseases », Biomarker Research, vol. 2, no 1, , p. 1 (ISSN 2050-7771, PMID 24398220, PMCID PMC3892095, DOI 10.1186/2050-7771-2-1, lire en ligne, consulté le )
- ↑ (en) Katherine Radek et Richard Gallo, « Antimicrobial peptides: natural effectors of the innate immune system », Seminars in Immunopathology, vol. 29, no 1, , p. 27–43 (ISSN 1863-2297 et 1863-2300, DOI 10.1007/s00281-007-0064-5, lire en ligne, consulté le )
- ↑ (en) Hajime Kono et Kenneth L. Rock, « How dying cells alert the immune system to danger », Nature Reviews Immunology, vol. 8, no 4, , p. 279–289 (ISSN 1474-1741, PMID 18340345, PMCID PMC2763408, DOI 10.1038/nri2215, lire en ligne, consulté le )
- ↑ (en) T. Kawai et S. Akira, « TLR signaling », Cell Death & Differentiation, vol. 13, no 5, , p. 816–825 (ISSN 1476-5403, DOI 10.1038/sj.cdd.4401850, lire en ligne, consulté le )
- ↑ Monica Heger (2014) Immunosequencing Comes of Age as Researchers Demo Clinical Utility, Aim for FDA-cleared Products ; Genome Web ; 9 décembre 2014
- ↑ Robins, H. (2013). Immunosequencing: applications of immune repertoire deep sequencing. Current opinion in immunology, 25(5), 646-652.
- ↑ DeWitt, W., Lindau, P., Snyder, T., Vignali, M., Emerson, R., & Robins, H. (2014). Replicate immunosequencing as a robust probe of B cell repertoire diversity. arXiv preprint arXiv:1410.0350.
Voir aussi
Bibliographie
En français
- Jacques Thèze, La Force du système immunitaire : Vers de nouveaux traitements des plus grandes maladies, Paris, Éditions Odile Jacob, , 320 p. (ISBN 978-2-7381-3216-1)
- Charles Janeway, Kenneth Murphy, Paul Travers et Mark Walport, Immunobiologie, 3e édition, traduction de Pierre L. Masson, éditions De Boeck, 2009.
- David Male, Immunologie. Aide-mémoire illustré, traduction de la 4e édition anglaise par Paul Fonteneau, éditions De Boeck, 2005 (ISBN 2-8041-4715-0).
En anglais
Articles connexes
- Immunité humaine
- Immunité (médecine)
- Immunité passive
- Déterminant antigénique
- Diapédèse
- Phagocyte
- Immunosénescence
- Vaccination
Liens externes
- Ressources relatives à la santé :
- FMA
- Uberon
- (en) Medical Subject Headings
- (no + nn + nb) Store medisinske leksikon
- (cs + sk) WikiSkripta
- Notices dans des dictionnaires ou encyclopédies généralistes :