|
الضفادع الثعبانية العصر: الطباشيري المتأخر – الآن،100–0 مليون سنة | |
|---|---|
 | |
| المرتبة التصنيفية | رتبة[1][2] |
| التصنيف العلمي | |
| النطاق: | حقيقيات النوى |
| المملكة: | الحيوانات |
| الشعبة: | الحبليات |
| الشعيبة: | الفقاريات |
| غير مصنف: | رباعيات الأطراف |
| الطائفة: | البرمائيات |
| غير مصنف: | البرمائيات الملساء |
| الرتبة: | الضفادع الثعبانية |
| الاسم العلمي | |
| Gymnophiona [1][2] قسطنطين صموئيل رافينسك، 1814 |
|
 | |
الضفادع الثعبانية أو الثعابين العارية أو الديدان العمياء[3] (الاسم العلمي: Gymnophiona)(Caecilians) هي رتبة من البرمائيات التي تشبه ظاهريًا ديدان الأرض أو الثعابين. حيث تعيش هذه الأنواع في الأرض متخفية، مما يجعلها أقل فئة معروفة بين البرمائيات. وقد تم تجميع جميع أنواع الضفادع الثعبانية وأقاربها الموجودة من الحفريات على أنها سلالة أحادية من البرمائيات عديمة الأرجل. كما تُوزع معظم هذه الأنواع في المدارات الاستوائية لجنوب ووسط أمريكا وإفريقيا وجنوب أسيا. لا تتوفر معلومات كافية حول النظام الغذائي لأنواع الضفادع الثعبانية. الاسم العلمي من باللاتينية الجديدة مركب من كلمتين إغريقيتين gymn و تعني "عاري" و ophiona وتعني ثعبان.
الوصف
لا يوجد لدى أنواع الضفادع الثعبانية أية أطراف لذا تشبه الأنواع الصغيرة منها الديدان، بينما تشبه الأنواع الأطول التي يصل طولها إلى 1.5 م (4 قدم 11 بوصة) الثعابين. فذيلها قصير أو قد لا يوجد بها ذيل والمجرور (cloaca) يكون قريبًا من نهاية الجسم.
كما تتميز هذه الأنواع بجلد أملس وعادةً باهت يميل إلى القتامة، ومع ذلك يظهر لدى بعض الأنواع الأخرى ألوان زاهية. يوجد داخل الجلد قشريات قشريات (scales) (calcite) قشريات (حيوي). وبسبب هذه القشريات، كان يعتقد أن أنواع الضفادع الثعبانية ذات صلة بحفريات استجوسفيليا (Stegocephalia)، إلا أنه يعتقد الآن أنها تطور ثانوي، وأن المجموعتين على ما يبدو متصلين ببعضهما.[4] يتميز الجلد أيضًا بالعديد من الحلقات على شكل دوائر، تحيط جزئيًا بالجسم، بحيث تمنح هذه الأنواع مظهرًا مقسمًا. كما يحتوي جلد هذه الأنواع، مثل البرمائيات الحية، على غدد تفرز مادة التكويسين السامة التي تعيق بها فرائسها.[5] وقد ظهر في إفرازات الجلد من سيفونوبس بولينسيس (Siphonops paulensis) خصائص تحلل خلايا الدم.[6]
وتعتبر عوامل الرؤية عند الضفادع الثعبانية محدودة على إدراك الضوء القاتم،[7] حيث يظهر التشريح تكيفها بشكل كبير مع نمط الحياة داخل الحفر. وتتميز هذه الأنواع بوجود رأس قوية، ذات أنف مدببة تستخدمها لشق طريقها خلال الترب القوية أو الطين.[4] وفي معظم الأنواع، يوجد عدد قليل من العظام في الرأس ومرتبطة ببعضها البعض، ويوجد الفم في تجويف تحت الرأس. كما تتميز عضلات هذه الأنواع بقدرتها على الاندفاع في طريقها خلال الأرض، بهيكل وعضلات قوية تعمل كمكبس داخل الجلد والعضلات الخارجية. حيث تتيح للحيوان أن يثبت نهاية مؤخرتها في موضع وتجبر رأسها على التحرك للأمام ثم تقوم بسحب الجسم كله لأعلى حتى تصل في أمواج. في المياه أو الطين الرخو، تستبدل الضفادع الثعبانية السباحة بطريقة الإنكليس.[5] الضفادع الثعبانية في عائلة تيفلونستيدي (Typhlonectidae) عبارة عن أنواع مائية وتعتبر أيضًا الأكبر من نوعها. الأنواع التي تمثل هذه العائلة تتميز بأنها تنفرد بزعانف دهنية على طول الجزء الخلفي من الجسم، والتي تعمل على تحسين حركاتها في المياه.[8]
وتتميز جميع الأنواع خلافًا للأنواع البدائية من الضفادع الثعبانية بمجوعتين من العضلات لقفل الفكّ، بالمقارنة بالزوج المفرد الذي تم العثور عليه في المخلوقات الأخرى. كما تتطور هذه الميزة بشكل كبير في معظم القوارض الحفرية من الضفادع الثعبانية، ويبدوا أنها تساعد في الإبقاء على الرأس والفم في حالة صلبة مرنة.[5]
وللتكيف مع الحياة تحت الأرض، تتميز هذه الأنواع بأعين صغيرة مغطاة بالجلد من أجل الحماية، الأمر الذي أدى إلى سوء فهم حيث كان يقال أن هذه الأنواع عمياء. حيث تبيّن أن هذا الاعتقاد غير صحيح بالمرة، على الرغم من أن الرؤية لديها تقتصر على الإدراك في الضوء البسيط القاتم. وتتميز أنواع الضفادع الثعبانية بزوجين من المجسات، تقع بين العين والأنف. حيث تستخدم هذه المجسات من أجل القدرة على الشم، إضافة إلى ذلك الحاسة الطبيعية للشم التي تعتمد على الأنف.[5]
ويوجد لدى جميع أنواع الضفادع الثعبانية ما عدا النوعين — أتريتوتشونا إيزيلتي (Atretochoana eiselti) [9] وسيسلتيتا أوكرامي (Caecilita iwokramae) [10] — رئات وتستخدم أيضًا الجلد أو الفم لامتصاص الأكسجين. وبالرغم من أن الرئة اليسرى أصغر من اليمنى، فإن يعتبر مناسبًا لشكل الجسم الذي تم العثور عليه أيضًا في الثعابين.
التوزيع
تم العثور على أنواع الضفادع الثعبانية في المناطق الاستوائية الرطبة في شمال جنوب أسيا والهند وسيرلنكا وأجزاء من شرق وغرب إفريقيا وبحيرات سيتشيلس (Seychelles) في المحيط الهندي وجنوب أمريكا في شمال وشرق أمريكا الجنوبية. وفي إفريقيا تم العثور على الضفادع الثعبانية في غينيا بيساو (جيوتريبيتس) (Geotrypetes) جنوب مالاوي (إسكوليكومورفس)، (Scolecomorphus) بسجل غير مؤكد من غرب زيمبابوي. إلا أنه لا يوجد لها أيضًا أية سجلات في المناطق الكبيرة في الغابة الاستوائية في غرب إفريقيا. وفي أمريكا الجنوبية تمتد خلال شرق المناطق غير الاستوائية في البرازيل على طول بلاد الأرجنتين الشمالية المعتدلة. ويمكن رؤية هذه الأنواع أيضًا على الجنوب من بيونس أيريس، عندما تحمل بالمياه المتدفقة من نهر بارانا الذي يرد من أقصى الشمال. كما يمتد النطاق الأمريكي من شمال إلى جنوب المكسيك. ويتركز التوزيع أقصى الشمال على الأنواع اتشثيوفيس سيكيمينسيس (Ichthyophis sikkimensis) شمال الهند. كما تم أيضًا العثور على اتشثيوفيس (اتشثيوفيس) في جنوب الصين وشمال فيتنام. وفي جنوب شرق أسيا، تم العثور عليها أقصى شمال جافا وبوروني وجنوب الفيلبين، إلا أنها لا تعبر خط والاس (Wallace's Line) وغير موجودة في أستراليا أو الجزر القريبة.
التصنيف

يُشتق الاسم caecilian من الكلمة اللاتينينة caecus (caecus)، تعني "أعمى"، تشير إلى الأعين الصغيرة أو غير الموجودة. ويعود تاريخ الاسم إلى الاسم التشريحي للأنواع الأولى الموضحة من قِبل كارلوس ليناوس (Carolus Linnaeus) الذي أطلق عليها الاسم سيسليا تنتاسولاتا (Caecilia tentaculata).[4] كما يشتق الاسم التشريحي للفئات من الكلمة اليونانية γυμνος (جيمنوس، عارِ) وοφις (أوفيس، ثعبان)، حيث كان يعتقد أن الضفادع الثعبانية ذات صلة في الأساس بالثعابين.
أحدث تصنيف لأنواع الضفادع الثعبانية، بواسطة ويكنسون إت آل (Wilkinson et al). (2011)، يقسم جيمنوفيونا (Gymnophiona) إلى 200 نوع و9 عائلات تقريبًا.[11] منذ ذلك الحين، تم اكتشاف عائلة الضفادع الثعبانية العاشرة، برمائيات لا قوائم لها (Chikilidae).[12][13] ويستند التصنيف إلى تعريف شامل للمونوفيلا بناءً على معظم الأدلة المرفولوجية والجزئية الحديثة، وتقوم بحل مشاكل طويلة الأمد لبارافيلي من البرمائيات في التصنيفات السابقة دون اعتماد استثنائي على ترادف المعنى.[11][14]
- رينتريماتيدي (Rhinatrematidae) - جنسان، 11 نوعًا، جنوب إفريقيا
- اتشثيوفيدي (Ichthyophiidae) - 3 أجناس، 50 نوعًا؛ جنوب وجنوب شمال أسيا
- اسكوليكومروفيدي (Scolecomorphidae) - جنسان، 6 نوعًا؛ إفريقيا
- هيربيليدي (Herpelidae) - جنسان، 9 أنواع؛ إفريقيا
- برمائيات لا قوائم لها (Chikilidae) - 1 جنس، 3 أنواع؛ الهند
- البرمائيات (Caeciliidae) - جنسان، 42 نوعًا، وسط أمريكا
- تيفولونيستيدي (Typhlonectidae) - 5 أجناس، 13 نوعًا، جنوب أمريكا
- أندوتفيليدي (Indotyphlidae) - 7 أجناس، 21 نوعًا؛ إفريقيا، الهند، سيتشليس
- سيفونوبيدي (Siphonopidae) - 7 أجناس، 19 نوعًا، جنوب إفريقيا
- ديرموفيليدي (Dermophiidae) - 4 أجناس، 13 نوعًا، إفريقيا ووسط وجنوب أمريكا
تستند النسالة الحالية إلى الدليل الجزيئي التي تمت مراجعته من قِبل ويكينسون إت آل، [11] سان مورو إت آل. (2012),[15] إضافة إلى تشيكيليدي حيث تم تحديد علاقتها بواسطة الشارحين.[12]
التطور

يعرف القليل منها بتاريخها التطوري، حيث تركت سجل ضئيل من الحفريات. الحفرية الأولى، هي فقارة تعود إلى باليوسيني (Paleocene)، التي اكتشفت حتى 1972.[16] تأتي الحفريات السابقة الوهمية من فترة جوراسيك (Jurassic)، والجنس البدائي لها، أيوسيساليا (Eocaecilia) لديها أعضاء صغيرة وأعين متطورة جيدًا.[17] في الوصف الصادر في 2008 لحفريات البرمائيات (batrachian) جيروباتراكس (Gerobatrachus)، [18] اقترح أندرسون ومساعدوه أن الضفادع الثعبانية تأتي من مجموعة ليبوسبونديل (Lepospondyl) من تيترابودس (tetrapods)، وقد تكون على صلة وثيقة بـ حيوان سلوي (amniotes) عن الضفادع والسمندر، الذي يأتي من أسلاف تيمنوسبونديل (Temnospondyl). قد تساعد الأصول المتشعبة من الضفادع الثعبانية والبرمائيات الموجودة في شرح الفرق البسيط الكائن بين تواريخ الحفريات وأصول البرمائيات الحديثة، والتي تقترح أصول بيرميان (Permian) والتواريخ الأسبق، في مكون للفحم (Carboniferou)، التي تم التنبؤ بها من قِبل بعض دراسات الساعات الجزئية لسلاسل DNA. تؤيد معظم الدراسات الجزيئية والمرفولوجية للبرمائيات الموجودة، بالرغم من ذلك، مونوفيلي (monophyly) بالنسبة للسيسليانس والضفادع والسمندر وتستند معظم دراسات الجزيئيات الحديثة على العديد من البيانات الهندسية التي تقترح مكون للفحم–السابقة لأصل بيرميان للبرمائيات الموجودة.[19]
السلوك
التكاثر
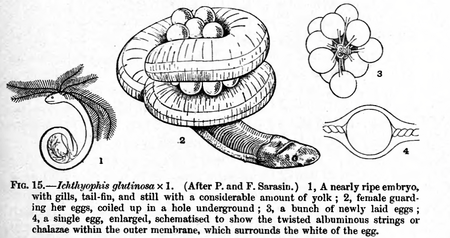
(Ichthyophis)
تعتبر الضفادع الثعبانية هي الأنواع الوحيدة من البرمائيات التي تستخدم التخصيب الداخلي بشكل فريد (على الرغم من أن معظم أنواع السمندر لديها آلية تخصيب داخلية والضفدع ذو الذيل في الولايات المتحدة تستخدم لاحقة تشبه الذيل للتخصيب الداخلي في بيئة تدفع فيها المياه بسرعة. كما يوجد لدى ذكور الضفادع الثعبانية أعضاء تشبه عضو التذكير، فالوديم، الذي يتم إدخاله في مجرور الأنثى لمدة تتراوح من 2 إلى 3 ساعات. حوالي 25% من الأنواع أوفيباروس (oviparous) (وضع البيض)؛ تحرس الإناث البيض. وبالنسبة لبعض الأنواع فإن صغار الضفادع الثعبانية هي بالفعل تكون منسلخة عند الفقس؛ يفقس البعض الآخر على شكل اليرقة. اليرقة ليست مائية بشكل كامل، لكنها تقضي وقتًا في أثناء النهار في التربة قريبة من الماء.[5]
75% من الأنواع تكون ولود (viviparous) بمعنى أنها تلد بالفعل نسل متطور. يتغذى الجنين داخل الأنثى على خلايا قناة المبيض (oviduct)، التي تأكل مستخدمة أسنان خاصة للكشط.
يفقس البيض عن أنواع بولينجيرولا تايتانا (Boulengerula taitana) تغذى صغارها من خلال تطوير الطبقة الخارجية من الجلد، الغنية بالدهون والمواد المغذية التي يستخدم الصغار أسنانهم لتقشريها. حيث تمكنهم من النمو عشرة مرات ضعف حجمهم في غضون أسبوع. ويتم استهلاك الجلد كل ثلاثة أيام، وهو الوقت الذي يكفي لنمو طبقة جديدة من الجلد، ويبدأ الصغار في العثور عليه وأكله ليلًا. وكان يعتقد أن الصغار يتغذون فقط على إفراز سائل مصدره أمهاتهم.[20][21]
بعض اليرقات، مثل الموجودة في تيفلونيكتس (Typhlonectes)، تولد بالعديد من الخياشيم الخارجية التي تختفي في الحال. تعتبر اتشثيوفيس (Ichthyophis) أوفيباروس (oviparous) وتعرف أيضًا بالرعاية الأمومية، حيث تحرس الأم البيض حتى يفقس.[4]
النظام الغذائي
لا تتوفر معلومات كافية حول النظام الغذائي لأنواع الضفادع الثعبانية. يبدو أن معظم الأنواع البالغة من الضفادع الثعبانية تتغذى معظمها على الحشرات وأنواع اللافقاريات الأخرى التي تم العثور عليها في مواطن الأنواع المعنية. تتكون محتويات المعدة من 14 نوعًا من أفروسيسليا تايتانا (Afrocaecilia taitana) من بقايا نباتات ومواد عضوية غير معروفة. حيث كانت البقايا غير المعروفة كثير، حيث تم العثور على ما يقال عليه رؤوس نمل أبيض.[22] حيث اقترح أن المواد العضوية غير المعروفة تظهر أن أنواع الضفادع الثعبانية تأكل الفتات (detritus)، ويعتقد البعض أنها بقايا من ديدان الأرض. يمكن لأنواع الضفادع الثعبانية غير القادرة التغذي بسهولة على ديدان الأرض، حيث تشتهر ديدان الأرض في أماكن عيش العديد من أنواع السيسليانس قد تقوم بعض أمهات الضفادع الثعبانية بإخفاء الجلد الغني بالمواد الغذائية لصغارها لتتغذى عليه.[20]
المراجع
- وصلة : التصنيف التسلسلي ضمن نظام المعلومات التصنيفية المتكامل — تاريخ الاطلاع: 19 سبتمبر 2013 — العنوان : Integrated Taxonomic Information System — تاريخ النشر: 2009
- النص الكامل متوفر في: http://www.mapress.com/zootaxa/2011/f/zt03148p055.pdf — المؤلف: David C. Blackburn و David B. Wake — العنوان : Class Amphibia Gray, 1825 — الصفحة: 39–55 — نشر في: التنوع البيولوجي للحيوان: مخطط تفصيلي للتصنيف العالي المستوى ومسح ثراء التصنيف
- علم الأحياء بواسطة بيتر هـ. ريفن،جورج ب. جونسون،جوناثان ب. لوسوس،كينيث أ. ماسون،سوزان ر. سنجر - تصفح: نسخة محفوظة 01 يناير 2017 على موقع واي باك مشين.
-
 هيو تشيشولم, المحرر (1911). . موسوعة بريتانيكا (الطبعة الحادية عشر). مطبعة جامعة كامبريدج.
هيو تشيشولم, المحرر (1911). . موسوعة بريتانيكا (الطبعة الحادية عشر). مطبعة جامعة كامبريدج.
- Nussbaum, Ronald A. (1998). Cogger, H.G. & Zweifel, R.G. (المحرر). Encyclopedia of Reptiles and Amphibians. San Diego: Academic Press. صفحات 52–59. .
- Elisabeth N. Ferroni Schwartz, Carlos A. Schwartz, Antonio Sebben (1998). "Occurrence of hemolytic activity in the skin secretion of the caecilian Siphonops paulensis". Natural Toxins. 6 (5): 179–182. doi:10.1002/(SICI)1522-7189(199809/10)6:5<179::AID-NT20>3.0.CO;2-M. PMID 10398514.
- S. M. Mohun, W. L. Davies J. K. Bowmaker, D. Pisani, W. Himstedt, D. J. Gower, D. M. Hunt, M. Wilkinson (2010). "Identification and characterization of visual pigments in caecilians (Amphibia: Gymnophiona), an order of limbless vertebrates with rudimentary eyes". The Journal of Experimental Biology. 213 (20): 3586–3592. doi:10.1242/jeb.045914.
- Piper, Ross (2007). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Greenwood Press.
- "Atretochoana eiselti". Natural History Museum. مؤرشف من الأصل في 21 يوليو 201522 فبراير 2012.
- Kaplan, Matt (18 November 2009). "Giant Lungless "Worm" Found Living on Land". National Geographic News. مؤرشف من الأصل في 21 أبريل 201822 فبراير 2012.
- Wilkinson, M. (2011). "A nine-family classification of caecilians (Amphibia: Gymnophiona)". Zootaxa. 2874: 41–64.
- Kamei, R.G. (2012). "Discovery of a new family of amphibians from Northeast India with ancient links to Africa". Proc. R. Soc. B. doi:10.1098/rspb.2012.0150.
- "New amphibian family found in India". CBC News. Associated Press. 21 February 2012. مؤرشف من الأصل في 19 يونيو 2013.
- Frost, Darrel R.; Grant, Taran; Faivovich, Julián; Bain, Raoul H.; Haas, Alexander; Haddad, Célio F.B.; De Sá, Rafael O.; Channing, Alan; Wilkinson, Mark (2006). "The Amphibian Tree of Life". Bulletin of the American Museum of Natural History. 297: 1–370, appendices. doi:10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2. ISSN 0003-0090.
- San Mauro, D. (2012). "Experimental design in phylogenetics: testing predictions from expected information". Systematic Biology. 61 (4): 661–674. doi:10.1093/sysbio/sys028.
- Estes, Richard (22 September 1972). "The First Fossil Record of Caecilian Amphibians". Nature. 239 (5369): 228. doi:10.1038/239228b0. مؤرشف من الأصل في 09 سبتمبر 201618 أغسطس 2009.
- Jenkins, Parish A. (16 September 1993). "An Early Jurassic caecilian with limbs". Nature. 365 (6443): 246. doi:10.1038/365246a0. مؤرشف من الأصل في 07 مايو 201618 أغسطس 2008.
- Anderson, Jason S.; Reisz, Robert R.; Scott, Diane; Fröbisch, Nadia B.; Sumida, Stuart S. (2008). "A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders". Nature. 453 (7194): 515–8. doi:10.1038/nature06865. PMID 18497824.
- San Mauro, D. (2010). "A multilocus timescale for the origin of extant amphibians". Molecular Phylogenetics and Evolution. 56 (2): 554–561. doi:10.1016/j.ympev.2010.04.019. PMID 20399871.
- Kupfer, Alex (2006). "Parental investment by skin feeding in a caecilian amphibian". Nature. 440 (7086): 926–929. doi:10.1038/nature04403. PMID 16612382.
- Vince, Gaia (12 April 2006). "Yummy mummy' worms feed their skin to offspring". New Scientist. مؤرشف من الأصل في 31 مايو 2015.
- Hebrard, J.J. (1992). "Notes on the habitat and diet of Afrocaecilia taitana". J. Herpetol. Society for the Study of Amphibians and Reptiles. 26 (4): 513–515. doi:10.2307/1565136. JSTOR 1565136.
- هيو تشيشولم, المحرر (1911). . موسوعة بريتانيكا (الطبعة الحادية عشر). مطبعة جامعة كامبريدج.
- Himstedt, Werner. Die Blindwühlen (باللغة الألمانية). Magdeburg: Westarp Wistshaften. .
- San Mauro, Diego (2004). "Phylogeny of caecilian amphibians (Gymnophiona) based on complete mitochondrial genomes and nuclear rag1". Molecular Phylogenetics and Evolution. 33 (2): 413–427. doi:10.1016/j.ympev.2004.05.014. PMID 15336675.
- San Mauro, Diego (2009). "Experimental design in caecilian systematics: phylogenetic information of mitochondrial genomes and nuclear rag1". Systematic Biology. 58 (4): 425–438. doi:10.1093/sysbio/syp043. PMID 20525595.