

الخلية هي الوحدة الاساسية التركيبية والوظيفية في الكائنات الحية، فكل الكائنات الحية تتركب من خلية واحدة أو أكثر، وتنتج الخلايا من انقسام خلية بعد عملية نموها. وتقسم الخلايا عادة إلى خلايا نباتية وخلايا حيوانية، وهناك تقسيمات أخرى؛ وتسمى مجموعة الخلايا المتشابهة في التركيب والتي تؤدي معًا وظيفة معينة في الكائن الحي عديد الخلايا بالنسيج. وتحتوي الخلية على أجسام أصغر منها تسمى عضيات، مثل أجسام جولجي، وهناك أيضًا النواة التي تحمل في داخلها الشيفرة الوراثية الدي أن إيه (حمض نووي ريبوزي منقوص الأكسجين). كما يحيط بالخلية غشاء يسمى بالغشاء الخلوي، ولدى الخلايا النباتية، جدار من السليلوز يسمى غشاء بلازمي، وهو غير مرن كالغشاء الخلوي.
وتمثل كل خلية من المئة تريليون خلية أو أكثر في جسم الإنسان بنية حية يمكنها أن تبقى على قيد الحياة إلى الأبد، وفي بعض الحالات يمكنها أن تعيد توليد نفسها إذا ما توفرت لها في السوائل المحيطة بها مواد مغذية مناسبة.[1]
تركيبها
لدراسة وظائف أعضاء الجسم وبنياته الأخرى لا بد من دراسة أولًا التنظيم الأساسي للخلية ووظائف كل مكوناتها. تتكون الخلايا حقيقية النواة من أربعة أجزاء رئيسية كما تشاهد بالمجهر الضوئي هي:
القسمان الرئيسيان في الخلية هما النواة nucleus والسيتوبلازم ( الهيولي ) cytoplasm ويفصل النواة عن السيتوبلازم غشاء النواة كما يفصل غشاء الخلية السيتوبلازم عن السائل المحيط الخارجي. في الأجزاء الأخرى في داخل الخلية والتي تكون عادة معلقة في الجبلة فيما يعرف باسم البنية الفيزيائية للخلية؛ وتشمل:
- غشاء الخلية.
- غشاء النواة.
- المتقدرات والجسيمات الحالة lysosomes والمريكزات centrioles.
- الشبيكة بلازمية.
- أجسام جولجي.
- الجسيمات الحالّة
- الميتوكندريا
- البلاستيدات الخضراء (بالنسبة الخلايا النباتية)
فالخلية ليست مجرد محفظة للسوائل والإنزيمات والمواد الكيميائية بل إنها تحوي أيضا بنيات فيزيائية منتظمة يسمى العديد منها العُضيات organelles. وتعطي الخواص الفيزيائية لهذه المواد بمجموعها أهمية وظيفية مهمة للخلية لا تقل عن أهمية مكوناتها الكيميائية؛ فمثلًا بدون إجدى أنواع هذه العضيات - وهي الميتوكوندريا mitochondria - يتوقف أكثر من 95 % من إمداد الخلية من الطاقة وتسمى المواد المختلفة التي تكون الخلية بمجموعها الجِبلة protoplasm التي تتكون بصورة رئيسية من خمس مواد أساسية، وهي: الماء والكهارل electrolytes والبروتينات والشحوم والسكريات.[2]
الماء: يكون الماء الوسط السائل الرئيسي للخلية. وهو يكون بنسبة تتراوح بين 70 و85 % وتوجد مذابة فيه الكثير من المواد الكيميائية في الخلية. كما يوجد البعض الآخر من المواد معلقة فيه بشكل دقائق صغيرة. وتتم العمليات الكيميائية في الخلية بين المواد الكيميائية المذابة في السائل أو عند حدود سطوح الجسيمات المعلقة والأغشية والماء.
الكهارل: أهم الكهارل electrolytes الخلية هي البوتاسيوم والمغنيسيوم والفوسفات والسلفات والبيكربونات وكميات صغيرة من الصوديوم والكلوريد والكالسيوم؛ مما يحفظ العلاقة المتبادلة بين السائلين داخل الخلية وخارجها. توفر الكهارل المواد الكيميائية اللاعضوية الضرورية للتفاعلات الخلوية؛ فمثلًا تساعد كهارل غشاء الخلية في انتقال الدفعات الكهركيميائية في الألياف العصبية والعضلية؛ كما تعين كهارل داخل الخلية العمليات المختلفة المحفزة إنزيميًا والضرورية لاستقلاب الخلية.
البروتينات: هذه هي أكثر المواد وفرة في معظم الخلايا بعد الماء؛ فهي تكون 10 - 20 % من كتلة الخلية. ومن الممكن تقسيم البروتينات هذه إلى بروتينات كريوية globular proteins وهي التي تكون الإنزيمات بصورة رئيسية، وبروتينات بنيوية structural proteins؛ وكمثال هام على البروتينات البنيوية يلاحظ بأن الجلد يتكون بصورة رئيسية من بروتينات بنيوية كما أن الشعر مكون بصورة تامة تقريبًا من نفس هذه البروتينات، ويوجد هذا النوع من البروتينات في الخلية على شكل خيوط طويلة ورفيعة وهي مكونة من مكاثير polymers عديدة من جزيئات بروتينة. وأهم وظائف هذه الخيوط داخل الخلية هو توفير الآلية التقلصية للعضلات. وتنتظم هذه الخيوط بشكل نبيبات مجهرية مكونة هياكل خلوية لبعض العُضيات كالأهداب، ومغازل الانقسام الفتيلي للخلايا التي تنقسم فتيليًا. كما توجد البروتينات الخيطية خارج الخلايا بصورة خاصة في الألياف الكلاجينية والمرنة للنسيج الضام والأوعية الدموية والأوتار والأربطة العضلية وما شابه ذلك. ومن الناحية الأخرى فإن البروتينات الكريوية هي من نوع مختلف تمامًا إذ أنها تتكون عادة من جزيئات بروتينية مفردة أو على الأكثر من تجمع عدد قليل من البروتينات بشكل كريوي بدلًا من الشكل الخيطي. وتقوم هذه البروتينات بصورة رئيسية بتكوين إنزيمات الخلايا - بخلاف البروتينات البنيوية - وهي عادة بروتينات ذؤوبة في سائل الخلايا أو أنها تكون أقسامًا متكاملة أو ملتصقة بالبنيات الغشائية داخل الخلايا. وتوجد الإنزيمات بتماس مباشر مع المواد الأخرى في داخل الخلية، وهي تحفز التفاعلات الكيميائية مثل تلك التي تشطر الجلوكوز إلى مكوناته وتوحدها بعد ذلك مع الأكسجين لتكون ثاني أكسيد الكربون والماء.كما أنهات تجهز في الوقت نفسه طاقة للوظائف الخلوية التي تحفز بسلسلة من الأنزيمات البروتينية.
الشحوم: وهي على أنواع متعددة ومختلفة تبحث كلها سوية بسبب خاصيتها العامة بكونها ذؤوبة في المذيبات الدهنية. وأهم الشحوم lipids الموجودة في معظم الخلايا الشحوم الفسفورية والكوليستيرول، وتكون هذه حوالي 2 % م نالكتلة الكلية للخلية. وتبرز الأهمية الخاصة للشحوم الفسفورية والكوليستيرول في الخلية لأنها بصورة عامة غير ذؤوبة بالماء ولذلك فإنها تكون حواجز غشائية تفصل مختلف الأحياز داخل الخلية. وبالإضافة للشحوم الفسفورية والكوليستيرول تحتوي بعض الخلايا كميات كبيرة من ثلاثيات الجليسريد triglycerides التي تسمى شحمًا متعادلًا. وتصل نسبة ثلاثيات الجليسريد في الخلايا الدهنية حوالي 95 % من كتلتها. ويمثل الدهن المخزون في هذه الخلايا المخزن الرئيسي للجسم للمغذيات المولدة للطاقة حيث يمكن تحليلها واستعمالها عندما يحتاج الجسم للطاقة.
السكريات: للسكريات carohydrates بصورة عامة وظائف ابتنائية قليلة في الخلية، فيما عدا كونه جزء من جزيئت البروتين السكري glycoprotein. ولكنها تقوم بدور رئيسي في تغذية الخلية. ومعظم خلايا الجسم في الإنسان لا تحتفظ بمخزون كبير من السكريات؛ فقد يصل مخزون السكريات فيها إلى 1 % فقط من مجموع كتلتها، ولكن هذا المخزون يزداد إلى 3 % في خلايا العضلات، واحيانًا يصل هذا المخزون إلى 6 % في خلايا الكبد. ومع ذلك توجد السكريات دائمًا بصورة جلوكوز في السائل خارج الخلايا المحيط بالخلايا وبصورة ميسرة لاستعمالها في الخلايا. وفي العادة تخزن كمية صغيرة من السكريات في الخلايا بشكل جليكوجين glycogen، وهو مكثور غير ذؤوب من الجلوكوز ومن الممكن أن يستعمل في الخلية لتوليد الطاقة فيها [3].
تراكيب الخلية


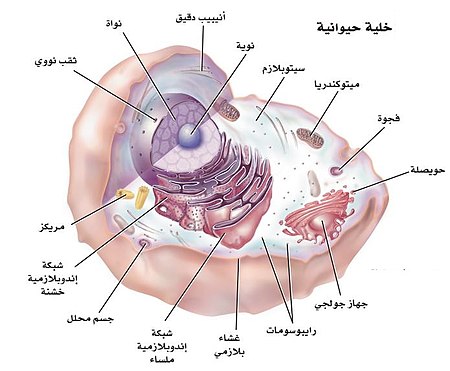
الخلية الحيوانية
| 1. النويّة | 
|
| 2. النواة | |
| 3. الجسيم الريبي | |
| 4. حويصل | |
| 5. الشبكة الإندوبلازمية الخشنة | |
| 6. جهاز جولجي | |
| 7. الغشاء الخلوي | |
| 8. الشبكة الإندوبلازمية الملساء | |
| 9. الميتوكوندريا | |
| 10. فجوة عصارية | |
| 11. السيتوبلازم | |
| 12. الجسيم الهاضمّ | |
| 13. مريكزات (سنتريولات) |
الخلية النباتية


مشاهدة الخلية

بالإمكان النظر إلى الخلية باستعمال المجهر بأنواعه المتعددة، ولكن في الغالب تكون العينة ميتة، وتتعرض للتلف بسبب ضغطها بين الشرائح، ولكن يمكن تفادي هذا الضرر عن طريق استعمال ملاقط ليزر لتثبيت الخلية دون لمسها، أو تحريكها، وتشريحها باستعمال مقص ليزر
حقيقية النواة
جميع الخلايا التي تمتلك نواة حقيقية وغلاف نووي ذا جدار مضاعف تسمي حقيقية النواة.
بدائية النواة


يضم هذا التقسيم الجراثيم (البكتيريا) والطحالب الزرقاء المخضرّة وتنقسم الخلية الغير حقيقية النواة إلى جزئين رئيسيين هما الهيولى (السيتوبلازم) وشبيه النواة ويسمى بعض الأحيان المنطقة النوية، ويحيط بهذين الجزئين الالغشاء الخلوي ويكون هذا الغشاء محاطًا أحيانًا (كما في بعض الجراثيم، وفي الطحالب) بجدار خلوي صلب أو شبه صلب يحافظ على الخلية ويؤمن لها الدعم يتراوح معدل حجم الخلية غير حقيقية النواة بين 1 - 10 ميكرومتر والخلية لا تستطيع الاستمرار في الحياة إذا تلف غشاؤها وينطوي الغشاء البلازمي في بعض غير حقيقيات النواة مكونًا طيات وثنايا، لكن هذه لا تكون منفصلة عن الغشاء البلازمي، لذلك لا تعتبر تراكيب داخلية بعض هذه الطيات الجسميات المتوسطة وتكون حاوية على الأنزيمات الأساسية الضرورية لعملية التنفس الهوائي والتي تحدث في المتقدرة (الميتوكندريا) المنتمية للخلايا حقيقية النواة، ولكن عدم وجود أغشية داخلية دائمة يعني عدم وجود تركيز موضعي للفعاليات والنشاطات محددًا بغشاء وهذا هو الاختلاف الرئيسي بين النوعين كما تختلف الريباسات (الرايبوزومات) في غير حقيقية النواة حيث تكون أصغر حجمًا ويتراوح قطرها بين 150- 200 انجستروم وتكون حرة في السيتوبلازم وتوجد في غير حقيقية النواة، إضافة إلى السيتوبلازم، مناطق كثيفة ذات شكل غير منتظم، وهي المناطق النووية ويشكل فقدان غشاء فاصل بين المادة الوراثية والسيتوبلازم فرقا أساسيا بين هذين النوعين من الخلايا (غير حقيقية النواة وحقيقية النواة).
مقارنة بين خصائص الخلايا حقيقية النواة وبدائية النواة
| غير حقيقية النوى | حقيقية النوى | |
|---|---|---|
| العضويات النموذجية | جراثيم, عتائق | أوليات, فطريات, نباتات, حيوانات |
| المقاييس (في المتوسط) | ~ 1-10 ميكرومتر (ميكرومتر) | ~ 10-100 ميكرومتر (حيوان منوي, من دون ذيله.) |
| نمط نواة (خلية) | منطقة شبيهة بالنواة; لا وجود لنواة حقيقية | نواة حقيقية ذات غشاء مضاعف |
| دي إن إي DNA | دائرية (عادة) | جزيئات خطية(صبغيات) مع هستونات بروتينية |
| RNA آر إن إي-/اصطناع البروتين | يتم في سيتوبلاسم | يتم تصنيع ال(RNA) في النواة اصطناع البروتين في السيتوبلاسم |
| ريبوسومات | وحدتين ريبوسيتين 50S+30S | وحدتين ريبوسيتين 60S+40S |
| البنية السيتوبلاسمية | بنيات قليلة العدد | بنية متينة مزودة بأغشية داخلية وهيكل خلوي |
| حركة خلوية | سوط مكون من الفلاجيلين | سياط وأهداب مكونة من نبيبات دقيقة |
| مصورات حيوية | لا يوجد | من بضعة إلى آلاف الميتوكوندريا) |
| صانعات يخضورية | لا يوجد | في أشنيات ونباتات |
| التشكل | عادة تكون بشكل خلية مفردة | خلية وحيدة، مستعمرات، متعدد خلايا مع وجود تمايز خلوي |
| الانقسام الخلوي | انشطار ثنائي (انقسام بسيط) | انقسام متساو انقسام منصف |
مقارنة البنية بين خلية نباتية وخلية حيوانية
| خلية حيوانية نموذجية | خلية نباتية نموذجية | |
|---|---|---|
| عضيات خلوية |
|
|
| بنى إضافية |
|
- مقالة مفصلة: الفرق بين الخلية الحيوانية والخلية النباتية
المقارنة بين الخلية الحيوانية وأشكال الحياة دون الخلية
يعتقد الكثيرون بأن الخلية هي أصغر أنواع الحياة. ولكن الخلية هي كائن حي شديد التعقيد واحتاجت لعدة ملايين من السنين حتى تطورت لشكلها الحالي بعد ظهور أول أشكال الحياة الأولية. وبمقارنة الحجوم النسبية لكل من أصغر فيروس معروف، وفيروس أكبر نسبيًا، والجرثومة والخلية ذات النواة وُجِد أن قطر الخلية يزيد 1000 مرة على قطر أصغر فيروس، ولذلك يزيد حجمه حوالي بليون مرة على حجم الفيروس الصغير، وبالمقارنة مع ذلك فإن التنظيم الوظيفي والتشريحي للخلية هو أكثر تعقيدًا من تنظيمات الفيروسات. والمكون الرئيسي الضروري المولد للحياة في الفيروس الصغير جدًا هو الحمض النووي المطمور في غلاف بروتيني، وهو مكون من نفس المكونات ( الـ DNA، والـ RNA ) التي توجد في خلايا الثدييات وتتمكن من التناسخ الذاتي عند توفر الظروف المناسبة لها. وبهذا فإن الفيروس يتمكن من الانتشار السلالي من جيل لآخر ولهذا فإنه يكون بنية حية شبيهة بخلية الإنسان وبالبنيات الحية الأخرى. وبعد نشوء الحياة ندمجت مواد كيميائية أخرى مع الحمض النووي والبروتينات البسيطة وأصبحت أجزاء متكاملة من الكائن الحي وبدأت تظهر وظائف متخصصة في مختلف أقسام الفيروس، ثم تكون غشاء حول الفيروس وظهر مطرس سائل داخل ذلك الغشاء. ثم تكونت مواد كيميائية متخصصة في المطرس لتقوم بوظائف خاصة وظهرت إنزيمات بروتينية قادرة على تحفيز التفاعلات الكيميائية فتتمكن بذلك من التحكم في نشاطات الكائن الحي. وفي مراحل لاحقة تطورت عضيات داخل الكائن الحي تمثل البنيات الفيزيائية للتجمعات الكيميائية التي تقوم بعملياتها بكفاءة أكبر من تلك التي يمكن أن تقوم بها مواد كيميائية مبعثرة خلال المطرس السائل matrix. وأخيرًا تطورت في الخلايا المنواة عضيات أكثر تعقيدًا وأهمها جميعًا هي النواة نفسها التي تميز هذا النمط من الخلايا عن كل أنواع الحياة الأقل منها؛ إذ أن هذه البنية توفر مركزًا للتحكم بكل النشاطات الخلوية وتوفر توالدًا مضبوطًا لأجيال متتالية لخلايا جديدة بحيث يكون لكل خلية جديدة منها نفس بنية الخلية المولدة لها[4].
الأنواع الأساسية للخلية
تعَد الخلايا الوحدات الأساسية للمخلوقات الحية جميعها. وتوجد بأشكال وحجوم مختلفة. كما تختلف بناءً على الوظيفة التي تؤديها في المخلوقات الحية. تشترك جميع الخلايا في صفة شكلية هي الغشاء الخلوي كما في الشكل الآتي:

والغشـاء الخلوي، هو حاجز خاص يساعد على ضبط ما يدخل إلى الخلية وما يخرج منها. وللخلايا عادةً عدد من الوظائف المُشترَكة. فمثلًا تحوي جميع الخلايا مادة وراثية تعطي معلومات وتعليمات للخلية لإنتاج مواد تحتاج إليها. كمـا تحلل الخلايـا الجزيئات لإنتاج الطاقـة اللازمة لعمليات الأيض. وقد قسـم العلمـاء الخلايا إلى مجموعتين، همـا: الخلايا البدائية النـواة، كما يتبين من الشكل السابق أن الخلايا الحقيقية النواة أكبر من الخلايا البدائية النواة، بل قد يزيد حجمها عليها مئة مرة. إن العلماء وضعـوا الخلايا في مجموعتين؛ بناءً على تراكيبهما الداخلية. فكلتاهما تحوي غشـاء خلويًّا، إلا أن إحداهمـا تحـوي تراكيـب داخلية مميزة تسـمى العضيـات وهي تراكيب خاصة تقوم بوظائف محددة.
تحوي الخلايا الحقيقية النواة نواة وعضيات أخرى محاطة بأغشية؛ أما النواة فهي عضيـة مركزية مميزة تحـوي المادة الوراثية على شـكل الحمض النووي الريبوزي المنقوص الأكسجين. تسـمح العضيات للخليـة بالقيـام بوظائفها فـي أجزاء مختلفة مـن الخلية في الوقت نفسـه. وتتكون معظـم المخلوقـات الحية من الخلايـا الحقيقية النواة. كمـا أن بعض المخلوقات الحيـة الوحيـدة الخليـة - ومنهـا بعـض الطحالب والخميـرة - مـن المخلوقات الحقيقية النواة. أمـا الخلايـا البدائيـة النواة فهـي خلايا ليـس لها نـواة أو عضيات محاطة بغشـاء. ومعظـم المخلوقات الحية الوحيدة الخلية - ومنها البكتيريا - خلايا بدائية النواة؛ لذا سميت الخلايا البدائية النواة.
البنيات الغشائية للخلية
تبطن كل عضيات الخلية أساسًا بأغشية تتكون مبدئيًا من الشحوم والبروتينات. وتشمل هذه الغشية كلًا من غشاء الخلية، وغشاء النواة، وغشاء الشبكة الهيولية الباطنة، وأغشية المتقدرات والجسيمات الحالة، وجهاز جولجي، بالإضافة لغيرها من الأغشية. و توفر شحميات الأغشية حاجزًا يمنع حركة المياه الحرة والمواد الذؤوبة فيه من حيز خلوي لحيز خلوي آخر. ومن الناحية الأخرى تخترق جزيئات بروتينات الغشاء خلال سمكه كله فتقطع بذلك استمرارية الحاجز الشحمي وتوفر بذلك ممرات على هيئة قنوات لعبور المواد الخاصة خلال الغشاء. كما أن العديد من بروتينات الغشاء هي إنزيمات تحفز العديد من مختلف التفاعلات الكيميائية [5].
غشاء الخلية
غشاء الخلية الذي يحوطها تمامًا هو غشاء رقيق ذو نية مرنة يتراوح سمكه بين 7.5 و10 نانومترات، ويتكون بصورة تامة تقريبًا من بروتينات وشحميات. وتركيبه التقريبي هو 55 % بروتينات، و25 % شحميات فسفورية، و13 % كوليستيرول، و4 % شحميات أخرى، و3 % سكريات [6][7].
وظيفة الغشاء الخلوي
إن عمليـة المحافظة على اتـزان البيئة الداخليـة للمخلوقات الحية تسـمى الاتزان الداخلـي، وهى جزء أساسي من فيزيولوجيا الخلية وهي ضرورية لبقاء الخلية. ويعد الغشـاء البلازمي أحد التراكيب المسـؤولة أساسًا عن الاتزان الداخلي؛ فهو حاجز فاصل رقيق مرن بين الخلية وبيئتها يسمح بمرور المواد المغذية إلى الخلية وخروج الفضلات والمواد الأخرى. تحوي جميع الخلايا البدائية والحقيقية النواة غشـاءً خلويًّا يفصلها عن البيئة السائلة التي توجد فيها. تُعدُّ خاصية النفاذية الاختيارية إحدى الصفات المهمة للغشاء الخلوي؛ إذ يسمح الغشاء الخلوي بمرور بعض المواد إلى الخلية، ويمنع مرور أخرى.

مكونات الغشاء الخلوي
معظم الجزيئات في الغشاء البلازمي دهون. والدهون جزيئات كبيرة من الجلسترول وثلاثة أحماض دهنية. فإذا حلَّ مكان أحد الأحماض الدهنية مجمموعة فوسفات تتكون الدهون المُفسفَرة

والدهون المُفسفَرة جزيئات تكونت من سلسلة أساسية من الجلسترول وسلسلتين من الأحماض الدهنية ومجموعة فوسفات. ويتكوَّن الغشاء البلازمي من طبقتين من الدهون المُفسفَرة المزدوجة تترتب ذيلًا مقابل ذيل، وبطريقة تسمح بأن يبقى الغشاء البلازمي قائمًا في بيئة سائلة.
طبقة الدهون المُفسفَرة المزدوجة
لاحظ شكل الطبقة المزدوجة من الدهون المُفسفَرة سترى أن كل طبقة دهون مُفسفَرة رُسمَت على شكل رأس له ذيلان فيما يعرف باسم الطبقة الشحمية المزدوجة؛ حيث تُكوِّن مجموعة الفوسفات رأسًا قطبيًّا في كل طبقة من الدهون المُفسفَرة. وينجذب الرأس القطبي إلى الماء، لأن الماء قطبي أيضًا. أما ذيلا الأحماض الدهنية فهما غير قطبيان ويتنافران مع الماء. وهو غشاء رقيق من الشحميات بسمك جزيئين يتواصل حول كل سطح الخلية. وتنتشر داخل هذه الطبقة الشحمية جزيئت بروتين كروية الشكل. وتتكون الطبقة الشحمية المزدوجة من الشحميات الفسفورية والكولستيرول بصورة تامة. وأحد أقسام جزيء الشحميات الفسفورية والكوليستيرول ذؤوب بالماء أي أنه أليف للماء hydrophilic، بينما يكون قسمه الآخر كاره للماء hydrophobic [8]. فجذر فسفات الشحم الفوسفاتي أليف للماء بينما الجذر الشحمي للحمض الدهني ذؤوب بالدهن. ويحوي الكولستيرول جذر هيدروكسيل ذؤوب بالماء ونواة ستيروئيدية ذؤوبة بالدهن. ولأن الماء يطرد الأجزاء الكارهة للماء لهذين الجزيئين بينما هي تجاذب مع بعضها البعض بصورة متبادلة؛ فلذلك تكون لها نزعة للاصطفاف كما في الصورة السابقة، وتكون بتماس مع الماء المحيط بالغشاء [9].
تشكل جزيئات طبقتي الدهون المُفسفَرة ما يشبه الشطيرة، حيث تُكوِّن فيها ذيول الأحماض الدهنية الجزء الداخلي (الأوسط) من الغشاء البلازمي، في حين تُكوِّن رؤوس الدهون المُفسفَرة مواجهة للبيئة السائلة داخل الخلية وخارجها، ويُعدُّ التركيب المزدوج مهمًّا في تكوين الغشاء البلازمي وأدائه لوظيفته. تترتَّب الدهون المُفسفَرة بطريقة تجعل الرؤوس القطبية هي الأقرب إلى جزيئات الماء، والذيول غير القطبية هي الأبعد عنها. وعندما تتجمَّع جزيئات الدهون المُفسفَرة معًا بهذا النمط فإنها تُشكِّل حاجزًا سطحه قطبي وأوسطه غير قطبي. وذلك الحاجز الشحمي لغشاء الخلية يمنع نفاذية الماء خلاله ولذلك لا تمرُّ المواد الذائبة في الماء بسهولة عبر الغشاء البلازمي؛ لأن وسط الغشاء غير القطبي يُعيقها. وهكذا يستطيع الغشاء البلازمي فصل بيئة الخلية الداخلية عن بيئتها الخارجية[5]. ويكون الغشاء مزدوج الطبقة الشحمية حاجزًا رئيسيًا غير نفوذ للمواد الاعتيادية الذؤؤوبة بالماء كالأيونات والجلوكوز واليوريا وغيرها. ومن الناحية الأخرى تتمكن المواد مثل الأكسجين والكحول من اختراق هذا الجزء من الغشاء بسهولة. وهناك سمة خاصة بهذه الطبقة الشحمية المزوجة وهي أنها سائلة وليست صلبة، ولذلك فإن أجزاء من الغشاء تتمكن من الجريان من نقطة لنقطة أخرى في الغشاء نفسه. كما تحاول البروتينات أو المواد الأخرى المذابة أو العائمة في الطبقة الشحمية المزدوجة من الانتشار في كل مناطق غشاء الخلية.
مُكوِّنات الغشاء الخلوي الأخرى
يبين شكل الغشاء الخلوي كتلًا عائمة في الطبقة الشحمية المزودجة. وهذه هي بروتينات الغشاء ويتكون معظمها من بروتينات سكرية. ويوجد منها نوعان وهما: البروتينات المندمجة التي تخترق كل سمك الغشاء، والبروتينات المحيطية الملتصقة بسطح الغشاء من دون اختراقه. ويوف العديد من البروتينات المندمجة integral proteins قنوات ( أو مسام ) تمر خلالها المواد المذابة في الماء وخاصة الأيونات التي يمكنها الانتشار بين السائل خارج الخلايا والسائل داخل الخلايا. ولكن لهذه البروتينات خواص انتقائية تفضل انتشار بعض المواد أكثر من غيرها. كما يعمل البعض منها كبروتينات حمالة carrier proteins لنقل المواد الكبيرة التي هي أكبر من أن تمر خلال القنوات. وبالإضافة لذلك يعمل البعض منها كإنزيمات. وتوضع البروتينات المحيطية بصورة كاملة أو شبة كاملة تقريبًاعلى داخل غشاء الخلية، وهي عادة ما تكون ملتصقة بأحد البروتينات المندمجة وهي تعمل عادة كإنزيمات أو كأنماط أخرى لضوابط الوظيفة داخل الخلايا [10].
يوجد على السطح الخارجي للغشاء البلازمي بروتينات، تُسمَّى المُستقبِلات، تُرسِل إشارات إلى داخل الخلية. كما تقوم بروتينات الغشاء البلازمي الموجودة على السطح الداخلي له بربطه مع تراكيب الدعم الخلوية الداخلية، مما يُعطي الخلية شكلًا مُميَّزًا. كما تخترق بروتينات أخرى الغشاء كله فتكوِّن قنوات تدخل من خلالها بعض المواد الخلية أو تخرج منها. وتنقل البروتينات الناقلة المواد التي تحتاج إليها الخلية أو الفضلات عبر الغشاء البلازمي. ومن المواد التي تنتقل عبر طبقة الدهون المُفسفَرة في الغشاء البلازمي الكولسترول، والبروتينات، والكربوهيدرات. فتُلاحظ أن البروتينات تُسهم في خاصية النفاذية الاختيارية للغشاء البلازمي[11].
يُساعِد الكوليسترول على منع التصاق ذيول الأحماض الدهنية في طبقة الدهون المفسفرة المزدوجة بعضها مع بعض، مما يُسهم في سيولة الغشاء البلازمي. وعلى الرغم من التوصية بعدم تناول المواد الغنية بالكوليسترول بكثرة، إلَّا أن الكوليسترول يُؤدي دورًا مهمًّا في تركيب الغشاء البلازمي، ويُعدُّ مُكونًا مُهمًّا أيضًا في الحفاظ على الاتزان الداخلي للخلية.
وهناك مواد أخرى في الغشاء البلازمي، ومنها الكربوهيدرات المرتبطة مع البروتينات لتُحدِّد خصائص الخلية وتٌساعدها على معرفة الإشارات الكيميائية. فمثلًا، تُساعِد الكربوهيدرات الموجودة على الغشاء البلازمي الخلايا المُقاوِمة للمرض على تمييز الخلية الضارة وتُهاجمها. تُكوِّن الدهون المُفسفَرة المزدوجة "بحرًا" تعوم فيه الجزيئات. ومفهوم البحر هذا هو أساس النموذج الفسيفسائي السائل في الغشاء البلازمي. وتتحرَّك الدهون المُفسفَرة جانبيًّا داخل الغشاء البلازمي. وفي الوقت نفسه، تتحرك مكونات أخرى ومنها البروتينات خلال الدهون المُفسفَرة. وبسبب وجود مواد مُختلِفة في الغشاء البلازمي، يتكوَّن نمط فُسيفسائي على سطح الخلية؛ كما أنَّ مكونات الغشاء البلازمي في حركة دائمة وثابتة، وينزلق بعضها فوق بعض.

سكريات غشاء الخلية
توجد سكريات الغشاء بصورة عامة تقريبًا متحدة مع البروتينات والشحوم بشكل بروتينات سكرية وشحوم سكرية. وفي الحقيقة إن معظم البروتينات المندمجة هي بروتينات سكرية وحوالي عُشر ( 1/ 10 ) الجزيئات الشحمية هي شحميات سكرية. ويبرز الجزء السكري glyco- من هذه الجزيئات بصورة عامة تقريبًا إلى خارج الخلية متدليًا من سطحها إلى خارجه [6][7]. كما يوجد العديد من مركبات السكريات proteoglycans، وهي مواد سكرية مرتبطة معًا بواسطة ليف بروتيني ومتصلة بلطف بالسطح الخارجي للخلية. وبهذا يكون لكل سطح الخلية غلاف سكري رخو يسمى الكأس السكري glycocalyx. ولهذه الأجزاء السكرية الملتصقة بالسطح الخارجي للخلية وظائف مهمة عديدة منها:
- العديد منها مشحون بشحنات سالبة مما يعطي معظم الخلايا شحنات سطحية سالبة تنفر منها المواد الأخرى السالبة الشحنة.
- ياتصق الكأس السكري لبعض الخلايا بالكؤوس السكرية للخلايا الأخرى وهكذا تلتصق الخلايا ببعضها البعض.
- تعمل بعض السكريات كمواد استقبالية لربط الهرمونات كالأنسولين الذي يحفز أنواعًا معينة من الأنشطة في داخل الخلايا.
- تدخل بعض السكريات في التفاعلات المناعية في الجسم [12].
الجِبلة والهيكل الخلوي
الجبلة هي البيئة شبة السائلة داخل الغشاء الخلوي. وقد اكتشف علماء الأحياء أن العضيات لا تسبح في الخلية، ولكن تدعمها تراكيب داخل الجبلة، كما في الشكل الآتي:

أما الهيـكل الخلوي فهو شبكة مُكوَّنة من خيوط بروتينية طويلة تدعم الخلية وتعطيها شكلها، وتثبِّت العضيات داخل الخلايا. كما يساعد الهيكل الخلوي على حركة الخلية وأنشطتها الأخرى. يتكون الهيكل الخلوي من تراكيب ثانوية تسمى الأنيبيبات الدقيقة والخيوط الدقيقة. والأنيبيبات الدقيقة تراكيب أسطوانية طويلة مجوفة من البروتين تكوِّن هيكلًا صلبًا للخلية، وتساعد على حركة المواد داخل الخلية. أما الخيوط الدقيقة فهي خيوط بروتينية رفيعة تساعد على إعطاء الخلية شكلها، وتمكِّن الخلية كاملة أو جزءًا منها من الحركة. وتتجمع الأنيبيبات والخيوط الدقيقة أو تتفرق لينزلق بعضها فوق بعض، مما يسهم في حركة الخلية.
الهيولي مملوءة ببعض الجسيمات الدقيقة والكبيرة وبالعضيات التي تتراوح أحجامها من بضعة نانومترات إلى عدة ميكرونات. ويسمى السائل الصافي من الهيولى الذي تنتشر فيه الجسيمات العصارة الخلوية cytosol، وهو يحوي قسم الهيولي الموجود تحت الخلية أعدادًا كبيرة من الخيوط المجهرية المكونة من لييفات أكتينية توفر إسنادًا شبه صلب لغشاء الخلية، وتسمى هذه المنطقة من الهيولى باسم القشرة cortex أو الهيولي الظاهرة ectoplasm. أما الهيولى الموجودة بين القشرة وغشاء النواة فهي سائل يسمى الهيولى الباطنة endoplasm. وتنتشر في الهيولى الكريات الدهنية المتعادلة وحبيبات الجليكوجين والريبوسومات والجبيبات الإفرازية وخمسة غضيات مهمة بصورة خاصة وهي الشبكة الهيولية الباطنة وجهاز جولجي والمتقدرات والجسيمات الحالة والجسيمات البيروكسيدية [11][13].
تراكيب الخلية
الخلية الحقيقية النواة لها مناطق مختلفة للقيام بالمهام. ويؤدي وجود العضيات المحاطة بالغشاء إلى القيام بعمليات كيميائية مختلفة في الوقت نفسه وفي أجزاء مختلفة من الجبلة. كما تقوم العضيات بالعمليات الخلوية الضرورية، ومنها بناء البروتين، وتحويل الطاقة، وهضم الغذاء، وإخراج الفضلات، وانقسام الخلية. ولكل عضية تركيب ووظيفة مميزان.
النواة
تحتاج الخلية إلى عضية لتنظيم عملياتها؛ النواة كما في الشكل الآتي:

فالنواة هي التركيب الذي ينظم عمليات الخلية، وهي مركز التحكم في الخلية، وهي التي تتحكم بالتفاعلات الكيميائية التي تحدث في الخلية وبعمليات توالدها. وتحوي النواة معظم الحمض النووي الريبوزي منقوص الأكسجين في الخلية الذي يخزن المعلومات التي تستخدم في بناء البروتينات اللازمة لنموها، ووظيفتها وتكاثرها ؛ وتحدد الجينات خواص إنزيمات بروتين الهيولى فتتحكم بهذه الطريقة في نشاطات الهيولى. وعند تحكمها في تكاثر الخلية تبدأ الجينات أولًا بتكثير نفسها، وبعد إنجاز ذلك تنشطر الخلية بعملية تسمى التفتل mitosis لتكون خليتين وليدتين تستلم كل واحدة منها إحدى مجموعتي الجينات. إن مظهر النواة تحت المجهر لا يعطي دلالة كبيرة على الآلية التي تستعملها في عملية التحكم الخلوي ؛ فمظهر الطور البيني للنواة ( الفترة بين التفتلين ) كما تظهر بالمجهر الإلكتروني المادة الكروماتينية غامقة الانصباغ خلال جبلة النواة. ومن السهولة التعرف على مادة الكروماتين أثناء فترة التفتل كجزء من الصبغيات المتميزة البناء والتي يمكن رؤيتها بسهولة بالمجهر الضوئي. وتحاط النواة بغشاء مزدوج يسمى الغلاف النووي nuclear envelope يتكون من غشائين منفصلين أحدهما داخل الثاني. فالغشاء الخارجي متواصل مع الشبكة الهيولية الباطنة، كما يتواصل أيضًا الحيز بين الغشائين النوويين مع الحيز داخل الشبكة الهيولية الباطنة. وهو مشابه للغشاء الخلوي إلا أن للغلاف النووي ثقوبًا تسمح للمواد الأكبر حجمًا بدخول النواة والخروج منها ؛ حيث تخترق الغلاف النووي عدة آلاف من المسامات النووية nuclear pores، وهي كبيرة جدًا إذ تصل أقطارها لحوالي 100 نانومتر. وتوجد معقدات من جزيئات بروتينية ملتصقة حول حواف المسامات بحيث يبقى قطر الباحة المركزية للمسام حوالي 9 نانومترات.[14][15] وحتى هذا الحجم يعتبر كبيرًا لكي يسمح لجزيئات البروتين الكبيرة ذات الـ 44000 وزن جزيئي من المرور خلالها بسهولة كما تسمح للبروتينات ذات الوزن الجزيئي الذي يقل عن 15000 لتمر خلالها بسرعة كبيرة. كما تنتشر المادة الكروماتينية داخل النواة وهي عبارة عن حمض نووي ريبوزي منقوص الأكسجين معقد يرتبط ببروتين
النويات
تحوي نوى العديد من الخلايا بنية أو أكثر خفيفة الانصباغ تسمى النويات nucleoli. وبعكس معظم العضيات التي لا تحوي النويات غشاءً محددًا بل هي عوضًا عن ذلك بنية بسيطة تحوي كمية كبيرة من الحمض الريبي النووي RNA، وبروتينات من الأنواع التي توجد في الريبوسومات. وتصبح النويات كبيرة جدًا عندما تقوم الخلية بالتركيب الفعال للبروتينات. وتركب جينات خمسة أزواج متفرقة من الصبغيات الـ RNA وتخزنه في النويات مبتدئة بـ RNA ليفي رخو يتكايف ليكون وحدات ثانوية حبيبية من الريبوسومات [16]. وتنقل هذه بدورها خلال مسامات غشاء النواة إلى الهيولى حيث تتجمع سوية لتكون ريبوسومات ناضجة تلعب دورًا ضروريًا في تكوين البروتينات إما في العصارة الخلوية cytosol أو بالترافق مع الشبكة الهيولية.[17]
رايبوسومات
من وظائف الخلية إنتاج البروتين. وتسمى العضيات التي تساعد الخلية على صنع البروتين الرايبوسومات. تلتصق بالسطوح الخارجية للأقسام العديدة من الشبكة الهيولية الباطنة أعداد كبيرة من حبيبات صغيرة تسمى الرايبوسومات ribosomes. وتسمى المواقع التي توجد فيها هذه الرايبوسومات باسم الشبكة الهيولية الباطنة الحبيبية granular endoplasmic reticulum. تتكون الرايبوسومات بصورة رئيسية من مكونين رئيسين، هما: حمض نووي ريبوزي RNA الذي يقوم بتركيب البروتين في الخلايا، وبروتين ؛ ولا تحاط الرايبوسومات بغشاء كباقي العضيات في الخلية. ويتم إنتاج الرايبوسومات في النوية داخل النواة. تحوي الخلايا الكثير من الرايبوسومات التي تنتج بروتينات مختلفة تستخدمها الخلية أو تنتقل إلى خارج الخلية لتستخدمها خلايا أخرى. كما تسبح بعض الرايبوسومات بحرية في الجبلة، في حين يرتبط بعضها الآخر مع عضيات أخرى تسمى الشبكة الهيولية الباطنة. وتنتج الرايبوسومات الحرة بروتينات تستخدم داخل جبلة الخلية[18]. أما الرايبوسومات المرتبطة فتنتج بروتينات يتم إحاطتها بغشاء أو تستخدمها خلايا أخرى.
الشبكة الاندوبلازمية الباطنة
هي نظام يتكوَّن من أكياس وقنوات متصلة ومتداخلة محاطة بغشاء مزدوج تعمل بوصفها مواقع لبناء البروتين والدهون؛ حيث تزودها الانثناءات والصفائح التي داخلها بمساحة سطح أكبر لكي تُنجز الوظائف الخلوية. وعندما ترتبط الرايبوسومات مع منطقة على الشبكة الهيولية الباطنة فإن هذه المنطقة تسمى شبكة هيولية باطنة خشنة لاحظ الشكل الآتي:
حيث تبدو الشبكة الاندوبلازمية الباطنة الخشنة كثيرة النتوءات والبروزات. وهذه البروزات هي الرايبوسومات التي تنتج البروتين تمهيدًا لنقله إلى الخلايا الأخرى أيضًا وجود مناطق على الشبكة الهيولية الباطنة لا ترتبط بها رايبوسومات. هي شبكة من النبيبات والبنيات الحويصلية المسطحة في الهيولي تسمى الشبكة الهيولية الباطنة. وتترابط هذه النبيبات والحويصلات مع بعضها البعض وتتكون جدرانها من أغشية شحمية ثنائية تحوي كميات كبيرة من البروتين شبيهة بغشاء الخلية. ويمكن أن تكون المساحة الكلية لسطح هذه البنية في بعض الخلايا - كخلايا الكبد مثلا ً - مساوية 30 - 40 ضعفًا من مجموع سطح غشاء الخلية. كما يوضح الشكل تفاصيل بنية جزء صغير من الشبكة الهيولية الباطنة حيث يبيبن أن الحيز داخل النبيبات والحويصلات يمتلأ بالمَطرِس الهيولي الباطن endoplasmic matrix، وهو وسط سائل يختلف عن السائل الموجود خارج الشبكة الهيولية الباطنة.
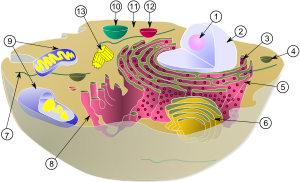
وتبين الصور المجهرية الإلكترونية بأن الحيز الموجود داخل الشبكة الهيولية الباطنة مرتبط بالحيز بين غشائي الغشاء النووي المزدوج. والأجزاء من الشبكة الهيولية الباطنة التي لا ترتبط معها رايبوسومات تسمى الشبكة الهيولية الباطنة الملساء[19]. وعلى الرغم من عدم وجود رايبوسومات في الشبكة الهيولية الباطنة الملساء إلا أنها تقوم بوظائف مهمة في الخلية. منها بناء الكربوهيدرات والدهون المعقدة، ومنها الدهون المُفسفَرة. كما تعمل الشبكة الهيولية الباطنة الملساء في الكبد على إزالة السموم الضارة من الجسم. وتدخل المواد التي تتكون في الأقسام المختلفة من الخلية إلى حيز الشبكة الهيولية وتنقل بعد ذلك إلى أقسام الخلية الأخرى. كما توفر المساحة السطحية الواسعة للشبكة والأجهزة الإنزيمية المتعددة المنتشرة على سطح أغشيتها النصيب الأكبر من الآلية اللازمة لفعالية الخلية الاستقلابية [20].
الشبكة الاندوبلازمية الملساء
هناك قسم من الشبكة الاندوبلازمية لا يحوي رايبوسومات ملتصقة بها؛ ويسمى هذا القسم باسم الشبكة الاندوبلازمية الملساء، وتقوم هذه الشبكة بتركيب المواد الشحمية في الكثير من العمليات الإنزيمية الأخرى في الخلية.
الميتوكوندريا
تسمى الميتوكوندريا mitochondria محطات توليد الطاقة للخلية. وبدونها لا تتمكن الخلايا من استخلاص كميات مناسبة من الطاقة من الغذيات والأكسجين؛ وكنتيجة لذلك تتوقف كل الوظائف الخلوية الأساسية. وكما هو مبين في الشكل السابق توجد هذه العضيات أساسًا في كل أقسام الهيولى ولكن يختلف عددها الكلي في الخلية الواحدة من أقل من مئة ميتوكوندريا إلى عدة مئات منها حسب كمية الطاقة التي تحتاجها الخلية. وبالإضافة لذلك تتركز الميتوكوندريا في أقسام الخلية المسؤولة عن أكبر جزء من استقلاب الطاقة في الخلية [21]. كما أنها تختلف في أشكالها وأحجامها إذ يبلغ طول قطر بعضها بضع مئات من النانومترات ؛ وهي كروية الشكل بينما يكون بعضها كبيرًا ويبلغ قطره 1 - 7 ميكرومترات ويكون متفرعًا أو خيطي الشكل.

1) الغشاء الداخلي
2) الغشاء الخارجي
3) الأعراف
4) المتركس
ويبين الشكل البنية الأساسية للميتوكوندريا ويظهر بأنها تتكون بصورة رئيسية من غشائين بروتينيين مزدوجي الطبقة الشحمية، أحدهما داخلي والآخر خارجي. وتكون عدة طيات من الغشاء الداخلي رفوفًا تلتصق عليها الإنزيمات المؤكسدة. وبالإضافة لذلك يمتلأ التجويف الداخلي للميتوكوندريا بمَطرِسٍ يحوي كميات كبيرة من الإنزيمات المذابة فيه والضرورية لاستخلاص الطاقة من الغذيات. وتعمل هذه الإنزيمات بالترافق مع الإنزيمات المؤكسدة على الفوف الداخلية في الميتوكوندريا لتؤكسد الغذيات مكونة ثاني أكسيد الكربون والماء. وتستعمل الطاقة المحررة لصنع مواد عالية الطاقة تسمى ثلاثي أدينوزين الفوسفات ATP، ثم ينقل الـ ATP إلى خارج الميتوكوندريا وينتشر في أنحاء الخلية ليحرر الطاقة كلما دعت الحاجة إليها للقيام بوظائف الخلية. والميتوكوندريا هي عضيات ذاتية التناسخ؛ وهذا يعني أن الميتوكوندريا تتمكن من توليد واحدة أخرى مثلها وثالثة أيضًا وهكذا كلما دعت الحاجة في الخلية لتوليد كميات إضافية من الميتوكوندريا وانتاج الطاقة. كما تحوي الميتوكوندريا الحمض النووي الريبي منقوص الكسجين DNA شبيه بذلك الذي يوجد في النواة ؛[22] فالـ DNA هو المادة الأساسية التي تحكم تناسخ الخلية، ولكن ليس لدرجة تامة لأنه في عملية تناسخ الميتوكوندريا يدخل الكثير من البروتينات والشحميات التي سبق تكونها إلى داخل الميتوكوندريا التي تتضخم عند ذاك وتتبرعم لتولد منها ميتوكوندريا جديدة وهكذا.
وظيفة الميتوكوندريا
تعتبر الوظيفة الرئيسية للميتوكوندريا هي استخلاص الطاقة من المواد الغذائية الرئيسية التي تستخلص الخلايا طاقتها منها، وهي الأكسجين وواحد أو أكثر من المواد الغذائية كالسكريات والدهون والبروتينات. وتتحول كل السكريات في جسم الإنسان بصورة أساسية إلى جلوكوز قبل وصولها إلى الخلية، وتتحول البروتينات إلى أحماض أمينية، وتتحول الدهون إلى أحماض دهنية؛ ثم تتفاعل تلك المواد الغذائية داخل الخلية كيميائيًا مع الأكسجين بتأثير الإنزيمات المختلفة التي تتحكم بسرعة تفاعلاتها وتوجه الطاقة التي تتحرر من ذلك بالاتجاه المناسب [23]. وتتم كل هذه التفاعلات المؤكسدة تقريبًا داخل الميتوكوندريا وتستعمل الطاقة المحررة لتوليد ثلاثي أديونوزين الفوسفات ATP الذي يستعمل هو نفسه وليست الأغذية الأصلية نفسها في أنحاء الخلية لتزويد الطاقة لكل التفاعلات الاستقلابية فيها.
جهاز جولجي

بعد بناء بعض البروتينات بواسطة الرايبوسومات على سطح الشبكة الهيولية الباطنة ينتقل بعضه بواسطة حويصلات تنفصل عن الشبكة الهيولية الباطنة لتصل إلى جهاز جولجي Golgi apparatus ؛ حيث يتعلق جهاز جولجي بصورة وثيقة بالشبكة الهيولية الباطنة وله أغشية شبيهة بأغشية الشبكة الهيولية الباطنة اللاحبيبية. كما في الشكل الآتي:
إن جهاز جولجي مكوّن من مجموعة من الأغشية المتراصة في أربع طبقات أو أكثر تعد ِّل البروتينات وترتبها وتغلفها داخل أكياس تسمى الحويصلات تنبثق عن جهاز جولجي، ثم تلتحم الحويصلات بالغشاء الخلوي لتحرر البروتينات إلى بيئة الخلية الخارجية. حيث تكون هذه الحويصلات رقيقة ومسطحة ومكدسة قرب النواة. ويكون هذا الجهاز بارزًا في الخلايا الإفرازية حيث يتوضع على جهة الخلية التي تقذف منها المواد الإفرازية [24]. ويعمل جهاز جولجي بالاشتراك مع الشبكة الهيولية الباطنة. كما يوجد حويصلات نقلية transport vesicles صغيرة، تسمى أيضًا حويصلات الشبكة الهيولية الباطنة، تنقطع باستمرار من الشبكة الهيولية الباطنة وتندمج بعد فترة قصيرة مع جهاز جولجي. وتنقل المواد بهذه الطريقة من الشبكة الهيولية الباطنة إلى جهاز جولجي، ومن ثم تعامل المواد المنقولة في جهاز جولجي لتكون الجسيمات الحالة والحويصلات الإفرازية أو المكونات الهيولية الأخرى.[25]
تركيب وتكوين البنيات الخلوية بواسطة الشبكة الإندوبلازمية الباطنة وجهاز جولجي
الوظائف النوعية للشبكة الإندوبلازمية الباطنة
يتسع انتشار الشبكة الإندوبلازمية الباطنة وجهاز جولجي وخصوصًا في الخلايا الإفرازية. وتتكون هاتان البنيتان بصورة مبدئية من أغشية مزدوجة الطبقة الشحمية وجدرانها مشبعة بإنزيمات البروتين التي تتمكن من تحفيز تركيب العديد من الموا التي تحتاجها الخلية. وبصورة عامة يبدأ معظم التركيب في الشبكة الإندوبلازمية الباطنة ثم تمرر المواد التي تصنع فيها إلى جهاز جولجي حيث تعالج بدرجة أكبر قبل تحريرها إلى الهيولى. ويمكن دور الشبكة الإندوبلازمية الباطنة الحبيبية في تكوين البروتينات؛ حيث تتصف الشبكة الإندوبلازمية الباطنة بوجود عدد كبير من الريبوسومات الملتصقة بالسطوح الخارجية للغشاء الشبكي، وتتركب جزيئات البروتين داخل بنيات الريبوسومات. كما أن الريبوسومات نفسها تطرح العديد من جزيئات البروتين المركبة لا إلى العصارة الخلوية cytosol بل عوضًا عن ذلك إلى مطرس الهيولى الباطنة خلال جدار الشبكة الهيولية الباطنة. و بنفس السرعة التي تدخل بها جزيئات البروتين إلى مطرس الهيولى الباطنة تسبب الإنزيمات في جدار الشبكة الإندوبلازمية الباطنة تغييرات سريعة في هذه الجزيئات إذ أن جميعها يُسكِر فورًا، أي أنها تقرن بأجزاء سكرية لتكون بروتينات سكرية glycoproteins. ولهذا فإن كل بروتينات الهيولى الباطنة هي أساسًا بروتينات سكرية بعكس البروتينات التي تولدها الريبوسومات في العصارة الخلوية والتي تكون بصورة عامة بروتينات حرة غير مرتبطة. وترتبط بعد ذلك البروتينات تعابريًا وتتطوى لتكون جزيئات أكثر اكتنازًا.
تركيب الشحوم بواسطة الشبكة الإندوبلازمية الباطنة وخصوصًا بالشبكة الإندوبلازمية الباطنة الملساء
تقوم الشبكة الإندوبلازمية الباطنة بتركيب الشحوم خصوصًا الشحوم الفسفورية والكولستيرول. وتُضَمن هذه بسرعة في الطبقة الشحمية المزدوجة للشبكة الإندوبلازمية الباطنة نفسها. ويؤدي ذلك إلى النمو المستمر لهذه الشبكة ويتم ذلك بصورة خاصة في القسم الأملس من الشبكة الإندوبلازمية الباطنة. و للمحافظة على عدم نمو الشبكة الإندوبلازمية الباطنة إلى أبعد من حدود الخلية تنفصل باستمرار حويصلات صغيرة تدعى حويصلات الشبكة الإندوبلازمية الباطنة أو الحويصلات الناقلة transport vesicles من الشبكة الملساء، ومعظم هذه الحويصلات تهاجر بسرعة إلى جهاز جولجي.
الوظائف الأخرى للشبكة الإندوبلازمية الباطنة
إن الوظائف المهمة الأخرى للشبكة الإندوبلازمية الباطنة، وخصوصًا للشبكة الملساء منها هي:
- أنها تحوي إنزيمات تتحكم في تحلل الجليكوجين عندما يستعمل لتوليد الطاقة.
- أنها تحوي أعدادًا كبيرة من الإنزيمات القادرة على نزع سموم المواد التي تؤذي الخلايا مثل العقاقير ؛ وهي تحقق ذلك بالتخثير [26] أو بالأكسدة أو بالهضم أو بالتضمين مع حمض الجليكورونيك أو بطرق أخرى غير ذلك.
الوظائف النوعية التركيبية لجهاز جولجي
بالرغم من أن الوظيفة الرئيسية لجهاز جولجي هي معالجة المواد التي سبق تكوينها في الشبكة الإندوبلازمية الباطنة، لكنه يمتلك أيضًا القدرة على تركيب بعض السكريات التي لا يمكن تكوينها في الشبكة الإندوبلازمية الباطنة. ويصدق هذا بصورة خاصة على حمض السياليك sialic acid والجلاكتوز. وبالإضافة لذلك فإنه يتمكن من تكوين مكاثير السكريد الكبيرة جدًا والمرتبطة بكميات قليلة من البروتين. وأهم هذه المواد هي حمض الهيالورنيك وسلفات الكوندرويتين. ومن خواص هاتين المادتين:
- أنهما المكونان الرئيسيان للبروتيوجليكانات porteoglycans التي تفرز في المخاط في الإفرازات الغدية الأخرى.
- وأنهما المكونان الرئيسيان للمادة الأساسية الإسنادية في الأحياز الخلالية والتي تعمل كحشوة بين ألياف الكُلاجين والخلايا الحية في الجسم.
- وأنهما المكونان الرئيسيان للمطرس العضوي في الغضاريف والعظام.
معالجة إفرازات الهيولى الباطنة بواسطة جهاز جولجي بتكوين الحويصلات
عندما تتكون المواد في الشبكة الإندوبلازمية الباطنة - وخصوصًا البروتينات - فإنها تنقل خلال النبيبات باتجاه أقسام الشبيكة الملساء داخل الهيولى التي تقع بالقرب من جهاز جولجي. وتنفصل عند هذه النقطة باستمرار حويصلات نقل صغيرة تنتشر إلى الطبقات الأعمق من جهاز جولجي. وتوجد داخل هذه الحويصلات البروتينات المركبة والنتاجات الأخرى. وتندمج هذه الحويصلات مباشرة مع جهاز جولجي وتفرغ محتوياتها من المواد إلى أحيازه الحويصلية، ثم تضاف لهذه الإفرازات أجزاء سكرية. وإضافة إلى ذلك فإن أهم وظيفة لجهاز جولجي هي تكثيف إفرازات الشبكة الإندوبلازمية الباطنة في هيئة رزم مركزة صغيرة. وعندما تمر هذه الإفرازات إلى الطبقات الدنيا من جهاز جولجي تتواصل عمليات التكثيف والمعالجة. وأخيرًا تنفصل الحويصلات الكبيرة والصغيرة منها وباستمرار عند الطبقة الدانية من جهاز جولجي حاملة معها المواد الإفرازية المكثفة التي تنتشر خلال الخلية. و لتحديد توقيت هذه العمليات فعندما تسبح الخلية الغدية في أحماض أمينية مشعة يكون بالإمكان الكشف عن جزيئات بروتينية مشعة حديثة التكوين في الشبكة الإندوبلازمية الباطنة الحبيبية خلال 3 إلى 5 دقائق، ويظهر في خلال 20 دقيقة وجود البروتينات حديثة التكوين في جهاز جولجي وتفرز في خلال 1 - 2 ساعة بروتينات مشعة من سطح الخلايا.
أنماط الحويصلات الإفرازية التي يكونها جهاز جولجي والأجسام الحالة
تكون الحويصلات التي يكونها جهاز جولجي في خلية إفرازية نشطة هي حويصلات إفرازية بشكل رئيسي، وتحوي هذه الحويصلات بصورة خاصة المواد البروتينية التي تفرز من سطح الخلية. وتنتشر هذه الحويصلات إلى غشاء الخلية وتندمج فيه وتفرغ محتوياتها إلى الخارج بآلية تسمى الالتفاظ الخلوى exocytosis ( الإيماس )، وهو عكس عملية الالتقام الخلوي endocytosis. وتزيد معدلات الإفراز الخلوي في أغلب الحالات بدخول أيونات الكالسيوم إلى داخل الخلية حيث تتفاعل هذه مع الغشاء الحويصلي بطريقة غير مفهومة لتولد اندغامها بالغشاء الخلوي. ومن الناحية الأخرى تخصص بعض الحويصلات للاستعمال داخل الخلية، فمثلًا تكوِن أجزاء متخصصة من جهاز جولجي الأجسام الحالة المذكورة سابقًا. ومن المعتقد أن أغشية هذه الأقسام المتخصصة تحوي مستقبلات كيميائية تسبب التصاق إنزيمات الهضم الحمضية. وتركز هذه الإنزيمات بهذه الطريقة ثم تُحرر من جهاز جولجي بشكل حويصلات الأجسام الحالة[27]. ويتكون بنفس الطريقة نوع آخر من الحويصلات وهي الجسيمات البيروكسيدية peroxisomes. ويعتقد بأن هذه الجسيمات تولدها الشبكة الإندوبلازمية الباطنة الملساء مع تكوين حويصلات النقل عوضًا عن تكوينها في جهاز جولجي. ومن المحتمل أن تكون هناك أيضًا مستقبلات خاصة في غشاء الشبكة الإندوبلازمية الباطنة تقوم بجلب الإنزيمات المؤكسدة التي تنطلق بعدئذ وتلتصق بشكل مركز بالجسيمات البيروكسيدية.
استعمال الحويصلات داخل الخلية لسد عوز الأغشية الخلوية
يندمج العديد من الحويصلات في النهاية مع غشاء الخلية أو مع أغشية البنيات الأخرى داخل الخلية كالميتوكوندريا، وحتى مع الشبكة الإندوبلازمية الباطنة نفسها. وتزيد هذه طبيعيًا من سعة الأغشية فتعوض بذلك عما يحصل فيها من نقص عند حدوث أي تدمير فيها هي نفسها؛ فمثلًا يفقد غشاء الخلية الكثير من مادته كل مرة تولد فيها حويصلة بلعمية أو احتسائية. وتعوض حويصلات من جهاز جولجي باستمرار كل ما يفقده غشاء الخلية. وباختصار يمثل الجهاز الغشائي للشبكة الإندوبلازمية الباطنة ولجهاز جولجي عضوًا استقلابيًا كبيرًا قادرًا على تكوين بنيات خلوية جديدة ومواد إفرازية تفرز من الخلية.
الفجوات

يوجد في الخلية حويصلات محاطة بغشاء، تُسمَّى الفجوات، تقوم بتخزين المواد بصورة مؤقتة في الجبلة. والفجوة - كالموجودة في خلية النبات - كيس يُستخدم في تخزين الغذاء والإنزيمات والمواد الأخرى التي تحتاج إليها الخلية، انظر الشكل الآتي:
تُخزِّن بعض الفجوات الفضلات. ومن المثير للاهتمام أن الخلية الحيوانية عادةً لا تحوي فجوات، وإذا حدث ذلك فإن الفجوات تكون أصغر كثيرًا مما هي عليه في الخلية النباتية.
الأجسام الحالّة
يوجد في الخلية حويصلات تحوي مواد تهضم، أو تحلل العضيات وجزيئات المواد المغذية الزائدة، تُسمَّى الجسيمات الحالة lysosomes، وهي عضيات حويصلية يولدها جهاز جولجي، وتنتشر خلال الهيولى. وتوفر هذه الجسيمات جهاز هضم داخل الخلايا يمكنه هضم البنيات والمواد غير المرغوبة خصوصًا الأجسام المعطوبة أو الغريبة عن الخلية مثل الجراثيم للتخلص منها. انظر الشكل الآتي:

يهضم اليحلول أيضًا البكتيريا والفيروسات التي تدخل الخلية.[28] ويمنع الغشاء المحيط باليحلول الإنزيمات الهاضمة داخلها من تحليل الخلية. وقد يلتحم اليحلول مع الفجوات، تم تطرح إنزيماتها في هذه الفجوات لتهضم الفضلات داخلها. وتتباين الجسيمات الحالة من خلية لأخرى وتتراوح أقطارها في العادة بين 250 و750 نانومترًا. وهي محاطة بغشاء نمطي مزدوج الطبقة الشحمية ومملوء بأعداد كبيرة من الحبيبات الصغيرة التي تترواح أقطارها بين 5 و8 نانومترات، وهي تجمعات بروتينية من إنزيمات هضمية. ويتمكن الإنزيم الهضمي من شطر المركب العضوي إلى قسمين أو أكثر باتحاد هيدروجين من جزيء ماء مع قسم من المركب، وباتحاد جزء الهيدروكسيل من جزيء الماء نفسه مه القسم الآخر من المركب؛ فمثلًا يهضم البروتين ليولد أحماضًا أمينية ويهضم الجليكوجين ليولد الجلوكوز. وهناك أكثر من 50 نوعًا من الإنزيمات الحمضية المختلفة التي وجدت في الجسيمات الحالة. أما المواد الرئيسية التي تهضمها فهي البروتينات، والأحماض النووية وعديدات السكريد المخاطية والشحوم والجليكوجين. ويمنع عادة الغشاء الذي يحيط بالجسيمات الحالة الإنزيمات الهضمية الموجودة في داخلها من ملامسة المواد الأخرى الموجودة في الخلية. ومع ذلك فهناك حالات مختلفة في الخلية تحطم فيها أغشية بعض الجسيمات الحالة فتسمح بتحرير الإنزيمات. وتشطر هذه الإنزيمات المواد العضوية التي تلامسها إلى مواد صغيرة عالية النفاذية كالأحماض الأمينية والجلوكوز [29][30].
فالوظيفة الرئيسية للأجسام الحالة هي هضم الأجسام الاحتسائية والبلعمية الغريبة في الخلية؛ حيث يتعلق بالحويصلة البلعمية أو الاحتسائية بعد ظهورها داخل الخلية جسيم lysosome واحد أو أكثر ويفرغ فيها هيدرولازاته الحمضية، فتتكون ذلك حويصلة هضمية digestive vesicle. وتبدأ الهيدرولازات hydrolases بهضم البروتينات والجليكوجين والأحماض النووية وعديدات السكريد المخاطية والمواد الأخرى التي توجد في تلك الحويصلة. وتتكون نتاجات هذا الهضم من جزيئات صغيرة من الأحماض الأمينية والجلوكوز والفوسفات وما شاكل ذلك من المواد التي يمكنها أن تنتشر خلال غشاء الحويصلة إلى الهيولى [31]. ويمثل باقي الحويصلة الهضمية - والذي يسمى الجسم الثمالي residual body - المواد غير المهضومة. وتفرغ هذه المواد في الغالب خلال غشاء الخلية بعملية تسمى الالتفاظ الخلوي exocytosis ( الإيماس )، وهي عكس عملية الالتقام الخلوي endocytosis. ولذا تسمى الأجسام الحالة أعضاء الهضم في الخلية.
نكوص الأنسجة والانحلال الذاتي للخلايا
غالبًا ما تتنكص أنسجة الجسم ويصغر حجمها، ويحدث ذلك مثلًا في الرحم بعد الولادة، أو في العضلات بعد فترة طويلة من عدم الاستعمال، أو في غدد الثدي بعد انتهاء فترة الإرضاع. والأجسام الحالة هي المسؤولة عن عمليات التنكس هذه؛ ومع ذلك لم تعرف حتى الآن الآلية التي تدفع بالأجسام الحالة لزيادة نشاطها عند قلة نشاط الأنسجة.
وهناك وظيفة أخرى للأجسام الحالة وهي إزالة الخلايا التالفة أو الأجزاء المخربة من الخلايا والأنسجة ؛ كالخلايا المخربة بالحرارة أو بالبرودة أو بالرضح أو بالكيميائيات أو بالعوامل الأخرى. ويؤدي تخريب الخلايا إلى تفجير الأجسام الحالة وتحرير الهيدرولازات ( الإنزيمات الحالة ) التي تبدأ فورًا بهضم المواد العضوية في محيطها. فإذا كان التخريب صغيرًا أُزيلت منطقته وأُعيد ترميم الخلية. أما إذا كان التخريب كبيرًا فتزال الخلية كلها بعملية تسمى الانحلال الذاتي autolysis ؛ وبهذه الطريقة تزال الخلية كلها وتتولد في العادة محلها خلية جديدة من نوعها نفسه بعملية التكاثر التفتلي للخلية المجاورة لها لتحل محل الخلية المخربة. كما تحوي الأجسام الحالة عوامل مبيدة للجراثيم يمكنها قتل الجراثيم المبلعمة قبل أن تتمكن من تخريب الخلايا [32]. وتشمل هذه العوامل الليزوزيم lysozyme الذي يقوم بإزابة أغشية خلايا الجراثيم، والليزوفرين lysoferrin الذي يربط الحديد والفلزات الأخرى الضرورية لنمو الجراثيم، والحمض عند درجة باهاء pH 5 تقريبًا بحيث ينشط الإنزيمات الحالة ويثبط بعض أنظمة استقلاب الجراثيم. كما تُخزن في الأجسام الحالة إنزيمات تتمكن من تحرير كُداسات الشحم وحبيبات الجليكوجين فتوفرها للاستعمال في مواقع أخرى في داخل الخلية أو في الجسم. وعند غياب هذه الإنزيمات الذي ينتج أحيانًا من اضطرابات جينية، تتجمع في الغالب كميات كبيرة من الشحوم والجليكوجين في خلايا العديد من الأعضاء وخاصة في الكبد ويؤدي ذلك إلى الموت المبكر.
الجسيمات البيروكسيدية
الجسيمات البيروكسيدية peroxisomes هي جسيمات شبيهة فيزيائيًا بالجسيمات الحالة، ولكنها تختلف عنها في أمرين مهمين: أولهما أنها تتكون بالتبرعم من الشبكة الهيولية الباطنة الملساء وليس من جهاز جولجي كما في حالة الأجسام الحالة ؛ وثانيهما أنها تحوي إنزيم الأكسيداز بدلًا من الهيدرولاز hydrolase ( الإنزيم الحال ). ويتمكن العديد من الاكسيدازات من توحيد أيونات الأكسجين مع أيونات الهيدروجين من مختلف المواد الكيميائية من داخل الخلية لتكوين ثاني أكسيد الهيدروجين H2O2. وهذه المادة نفسها وبدورها هي مادة مؤكسدة قوية تستعمل بالتزامن مع إنزيم الكاتالاز catalase، وهو أنزيم أكسيدازي آخر يوجد بكميات كبيرة في الجسيمات البيروكسيدية فيؤكسد مواد عديدة يمكن أن تكون سامة للخلية لولا أكسدتها ؛ فمثلًا يزال بهذه الطريقة نصف كمية الكحول الذي يتناوله الشخص بواسطة الجسيمات البيروكسيدية لخلايا الكبد.
الحويصلات الإفرازية
إحدى الوظائف المهمة للعديد من الخلايا هي إفراز بعض المواد الخاصة. وتتكون كل هذه المواد الإفرازية تقريبًا في منظومة الشبكة الهيولية الباطنة وجهاز جولجي، وتتحرر بعد ذلك من جهاز جولجي إلى الهيولى في داخل حويصلات التخزين المسماة الحويصلات الإفرازية secretory vesicles أو الحبيبات الإفرازية. و من الأمثلة الهامة عليها هي الحويصلات الإفرازية النمطية داخل الخلايا العنيبية للبنكرياس التي تخزن سليفات إنزيمات البروتين proenzymes ( وهي إنزيمات لم تنشط بعد ) والتي تفرز بعد ذلك خلال غشاء الخلية الخارجي إلى قناة البنكرياس ومن ثم إلى الإثنا عشر حيث تنشط وتقوم بوظائفها الهضمية.
المريكزات
يتكوَّن المُريكز من مجموعة من الأُنيبيبات الدقيقة، كما في الشكل الآتي:

تعمل في أثناء انقسام الخلية. وتوجد المُريْكزات في جبلة الخلايا الحيوانية ومعظم الطلائعيات، وهي قريبة من النواة.
الخيوط والبنيات النبيبية للخلية
تنتظم البروتينات الليفية للخلية عادة على شكل خيوط أو نبيبات وتتأصل هذه البروتينات كجزيئات بروتين طليعي تصنعها الريبوسومات التي تعوم بحرية في الهيولى. وتوجد هذه الجزيئات في أول الأمر مذابة في الهيولى، وتتكوثر polymerize بعد ذلك مكونة الخيوط filaments. وغالبًا ما توجد أعداد كبيرة من الخيوط المجهرية في المنطقة الخارجية من الهيولى التي تسمى الهيولى الظاهرة لتكون سندً مرنًا لغشاء الخلية. كما تنتظم الخيوط المجهرية في الخلايا العضلية بشكل آلة قلوصة خاصة وهي أساس التقلص العضلي في كل أنحاء الجسم. و يستعمل في كل الخلايا نمط خاص من الخيوط المكونة من جزيئات نبيبين tubulin مكوثرة تبني بنيات نبيبية تسمى النبيبات المجهرية microtubules [33]. وتحوي هذه دائمًا تقريبًا 13 سليفة خيط توجد مطروحة بطريقة متوازية وبترتيب دائري مشكلة أسطوانة طويلة مجوفة يبلغ قطرها حوالي 25 نانومترًا، وطولها من ميكرون واحد إلى عدة ميكرونات. وتترتب هذه الاسطوانات على شكل جزم مكونة كتلًا بنيوية قوية. ومع ذلك فالنبيبات المجهرية هي بنيات جاسئة تتكسر عند ثنيها بشدة. و مثال آخر على النبيبات المجهرية هي البنيات الآلية الأنبوبية للأهداب التي تعطيها صلابة بنيوية. وهي تتشعع إلى الأعلى ابتداء من هيولى الخلية حتى ذروة الهدب. كما تتكون أيضًا المريكزات والمغازل التفتلية للخلايا المنقسمة تفتليًا من النبيبات المجهرية [34]. ويتضح من هذا أن الوظيفة الأساسية للنبيبات المجهرية هي العمل كهيكل خلوي cytoskeleton يوفر بنيات صلبة فيزيائيًا لبعض أقسام الخلايا. كما يلاحظ بأن الهيولى غالبًا ما يجري بجوار النبيبات المجهرية ويمكن أن ينتج ذلك من حركة أذرع تبرز خارجة من النبيبات المجهرية[35].
الحُبَيبَات الخَيطِيَّة
تحتوي الخلايا على عضيات تنتج الطاقة تسمى الحُبَيبَات الخَيطِيَّة، وهي تحول جزيئات المواد المغذية (وخصوصًا السُّكَّريات) إلى طاقة قابلة للاستخدام. ومن الشكل الآتي:

يتبين أن للحبيبات الخيطية غشاء داخلي وغشاء خارجي كثير الطيات والانثناءات ليزودا الحبيبات الخيطية بمساحة سطح كبيرة تساعده على تكسير الروابط بين جزيئات السكر. وتُخزِّن الطاقة الناتجة ضمن روابط كيميائية في جزيئات أخرى لتستخدمها الخلية لاحقًا. ولهذا السبب، غالبًا ما تسمى الحبيبيات الخيطية بـ"مصانع الطاقة" في الخلايا.
الصانعات اليخضورية
للخلايا النباتية طريقتها الخاصة في استخدام الطاقة الشمسية. فبالإضافة إلى الحبيبيات الخيطية تحتوي خلايا النباتات وبعض الخلايا الأخرى الحقيقية النواة على الصانعات اليخضورية وهي عضيات تمتص الطاقة الضوئية وتحولها إلى طاقة كيميائية بواسطة عملية البناء الضوئي، تفحص الشكل الآتي:

ستلاحظ في هذا الشكل وُجود حجرات صغيرة وعديدة على شكل أقراص تسمى الثايلاكويدات داخل الغشاء الداخلي. حيث يتم امتصاص الطاقة الشمسية وتجميعها في الثايلاكويدات بواسطة صبغة الكلوروفيل التي تعطي الأوراق والسيقان اللون الأخضر.
الجدار الخلوي
هو تركيب آخر يوجد في الخلايا النباتية، والجدار الخلوي هو شبكة من الألياف السميكة الصلبة تحيط بالغشاء الخلوي من الخارج لتحمي الخلية وتوفر لها الدعامة. ويساعد الجدار الخلوي الصلب النباتات على الوصول إلى ارتفاعات مختلفة تتراوح بين أنصال الحشائش وغابات الشجر الأحمر. كما تتكون الجدران الخلوية في النباتات من كربوهيدرات معقدة تسمى السيليلوز والتي يعطي الجدار خاصية الصلابة. لاحظ الشكل الآتي:

الأهداب والأسواط
يغطي سطوح بعض الخلايا الحقيقية النواة تراكيب خاصة تُسمّّى الأهداب والأسواط، تمتد خارج الغشاء الخلوي وكما في الشكل الآتي:

فالأهداب زوائد قصيرة كثيرة العدد تُشبه الشعر وحركتها تُشبه حركة المجاديف في القارب أما الأسواط فهي أطول من الأهداب وأقل عددًا. وتتكوَّن الأهداب والأسواط من الأنيبيبات الدقيقة؛ المرتبة في نمط 2+9 حيث تترتب في صورة محيط دائرة، أي أن تسعة مجموعات مزدوجة من الأُنيبيبات تحيط بأنيبيبين منفردين في المركز. وعادة ما يكون للخلية سوط أو اثنان فقط.
وتحوي الهُديبات والأسواط في الخلايا البدائية النواة جبلةً محاطةً بغشاء خلوي. ويتكوَّن كل منها من وحدات بنائية من البروتين. وعلى الرغم من أن الأهداب والأسواط تُستخدَم في حركة الخلية إلا أن الأهداب تُوجد أيضًا في خلايا ثابتة وغير متحركة، ومنها الخلايا المبطنة للجهاز التنفسي في الإنسان والتي تغطيها الأهداب كما في الشكل السابق.
الأجهزة الوظيفية للخلايا
الأجهزة الوظيفية للخلية هي التي تجعل من الخلية كائنًا حيًا.
الابتلاع بالخلايا - الالتقام الخلوي
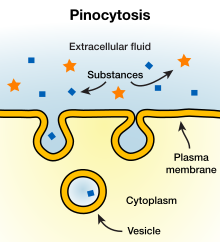
لكي تعيش الخلية وتنمو لا بد لها من الحصول على المواد المغذية والمواد الأخرى من السوائل المحيطة بها. وتمر معظم المواد خلال غشاء الخلية بالانتشار diffusion، وبالنقل الفعال active transport. ولكن الجسيمات الكبيرة تدخل إلى الخلية بآلية خاصة بالغشاء الخلوي تسمى الالتقام الخلوي endocytosis. والطريقتان الرئيسيتان للالتقام الخلوي هما الاحتساء pinocytosis، والبلعمة phagocytosis. والاحتساء يعني ازدراد حويصلات صغيرة جدًا تحوي السائل خارج الخلايا ؛ أما البلعمة فإنها تعني ازدراد جسيمات كبيرة كالجراثيم والخلايا والأجزاء المتنكسة من الأنسجة.
الاحتساء الخلوي
يحدث الاحتساء الخلوي pinocytosis باستمرار في أغشية معظم الخلايا ولكنه يتم بسرعة عالية خاصة في البعض منها[36] ؛ فهو يحدث مثلًا بسرعة عالية جدًا في البلعميات macrophages بحيث يبلع 3 % من غشائها بشكل حويصلات الاحتساء الخلوي صغيرة جدًا ( بقطر يصل إلى 100 - 200 نانومتر ) بحيث أن معظم خواصها لا يمكن تفريقها إلا بالمجهر الإلكتروني فقط.
والاحتساء الخلوي هو الوسيلة الوحيدة التي تمكن الجزيئات الكبيرة - مثل معظم البروتينات - من الدخول إلى الخلايا. وفي الواقع تزداد سرعة تكوْن حويصلات الاحتساء الخلوي كثيرًا عندما تلتصق الجزيئات الكبيرة بغشاء الخلية. ويوضح الشكل المقابل الخطوات المتتالية للاحتساء الخلوي، وهو يبين جزيئات بروتينة ملتصقة بالغشاء الخلوي، وتلتصق هذه الجزيئات عادة بالمستقبلات الموجودة في غشاء الخلية، وتكون هذه ذات نوعية خاصة بأنماط البروتينات التي تمتص. وتتركز هذه المستقبلات بصورة عامة في حفر صغيرة في غشاء الخلية تسمى الوهدات المغلفة coated pits. وتوجد على داخل غشاء الخلية تحت هذه الوهدات شبيكة من بروتين لييفي يسمى الكلاثرين clathrin بالإضافة إلى خيوط الأكتين والميوزين. وبمجرد اتحاد جزيئات البروتين مع المستقبلات تتغير رأسًا الخواص السطحية للغشاء الخلوي بطريقة تؤدي إلى انغمار الوهدة كلها للداخل، ثم تغلق البروتينات المتقلصة حوافها فوق البروتينات الملتصقة وعلى القليل من السائل خارج الخلايا وينقطع بعد ذلك مباشرة الجزء المنغمد من الغشاء من سطح الخلية مولدًا حويصلة احتساء خلوي pinocytic vesicle. وتحتاج هذه العملية إلى طاقة من داخل الخلية يوفرها لها ثلاثي فوسفات الأديونوزين ATP ؛ وهو مادة عالية الطاقة. كما أنها تحتاج إلى أيونات الكالسيوم في السائل خارج الخلايا الذي يحتمل تفاعله مع الخيوط المجهرية المتقلصة التي توجد تحت الوهدات المغلفة لتوفير القوة اللازمة لقطف الحويصلات بعيدًا عن غشاء الخلية.
البلعمة
تحدث البلعمة phagocytosis بنفس طريقة الاحتساء الخلوي عدا أنها تشمل مادة دقائقية كبيرة عوضًا عن الجزيئات [37]. وتمتلك بعض الخلايا الخاصة فقط هذه القدرة على البلعمة وأهمها بلعميات الأنسجة وبعض خلايا الدم البيضاء. وتبدأ البلعمة عندما ترتبط بالمستقبلات الموجودة على سطح البلعمية بروتينات أو عديدات سكريات كبيرة توجد على سطح الجسيم الذي يبتلع مثل الجراثيم أو الخلية الميتة أو حطام الأنسجة [38]. ففي حالة الجراثيم تلتصق هذه في العادة بأجسام مضادة نوعية، وتلتصق الأضداد بدورها بمستقبلات البلعمية. ويسمى هذا التوسط للأجسام المضادة الطهاية opsonization [39].
البلعمة في ثلاث خطوات: 1. مستقبلات البلْعمٌ السطحية الغير محدودة لا تؤدي البلعمة. 2. ربط المستقبلات يسبب لهم التعنقد. 3. يتم إطلاق عملية البلعمة ويتم أخذ الجسيمات من قبل البلعم.

1. مستقبلات البلْعمٌ السطحية الغير محدودة لا تؤدي البلعمة.
2. ربط المستقبلات يسبب لهم التعنقد.
3. يتم إطلاق عملية البلعمة ويتم أخذ الجسيمات من قبل البلعم.
و تتم مراحل البلعمة حسب الخطوات التالية [40]:
- تلتصق مستقبلات غشاء الخلية بالربائط السطحية للجسيم.
- تبرز حواف الغشاء حول نقطة الالتصاق للخارج، ويحدث ذلك خلال جزء من الثانية فتحيط بالجسيم ؛ ومن ثم تلتصق مستقبلات الغشاء بربائطه تدريجيًا وبتكاثر متزايد، ويتم كل ذلك بصورة مفاجئة وبأسلوب الزمام المنزلق.
- يحيط الأكتين والخيوط المتقلصة الأخرى الموجودة في الهيولى بالجسيم المزدرد، وتتقلص حول حوافه الخارجية دافعة به إلى الداخل.
- تقطع البروتينات المتقلصة بعد ذلك الحويصلة البلعمية تاركةً إياها في داخل الخلية بنفس الطريقة التي تتكون بها حويصلات الاحتساء الخلوي.
العضيات عندما تعمل
في ضوء الفهم الأساسي للتراكيب الموجودة في الخلية يُصبح فهم كيفية عمل هذه التراكيب معًا، وكيفية قيامها بوظيفتها الخلوية أسهل. فلو أخذنا مثلًا بناء البروتين فإنه يبدأ في النواة بحسب المعلومات التي يحويها الحمض النووي الريبوزي المنقوص الأكسجين حيث يتم نسخ هذه المعلومات الوراثية وينقلها إلى جزيء وراثي آخر يُسمَّى الحمض النووي الريبوزي ينتقل هذا الحمض الآخر وكذلك الرايبوسومات التي تنتج في النوية، من خلال ثقوب في الغلاف النووي إلى الجبلة. وتُسهِم كل من الرايبوسومات والحمض النووي الريبوزي في إنتاج البروتينات. ولكل بروتين يتكون على سطح الشبكة الهيولية الباطنة الخشنة وظيفة محددة؛ فربما يصبح بروتينًا يكوِّن جزءًا من الغشاء الخلوي، أو بروتينًا يُنقل خارج الخلية، أو بروتينًا يَنتقل إلى عضيات أخرى. وتعمل الرايبوسومات الأخرى الحرة في الجبلة على بناء البروتينات أيضًا
تنتقل معظم البروتينات التي تُصنع على سطح الشبكة الهيولية الباطنة الخشنة إلى جهاز جولجي؛ حيث تُغلف البروتينات في حويصلات لنقلها إلى العضيات الأخرى أو إلى خارج الخلية. وتَستخدم العضيات الأخرى البروتينات للقيام بالعمليات الخلوية. فمثلًا، يَستخدم اليحلول البروتينات، وخصوصًا الإنزيمات؛ لتهضم الغذاء والفضلات. وتَستخدم الحبيبيات الخيطية الإنزيمات لإنتاج الطاقة اللازمة للخلية.
حركات الخلية
إن أهم أنواع حركات الخلية التي تحدث في الجسم هو حركة الخلايا العضلية المتخصصة في العضلات الهيكلية والعضلات القلبية والعضلات الملساء ؛ وتكون هذه حوالي 50 % من كتلة الجسم كله. ولكن هناك نوعان آخران من الحركات التي تقوم بها الخلايا وهما:
ويعني التحرك الأميباني للخلايا أنه حركة خلية كاملة بالنسبة لمحيطها مثل حركة خلايا الدم البيضاء خلال أنسجة الجسم. وقد اكتسب هذا الاصطلاح اسمه من حركة الأميبا التي تتحرك بنفس هذا الأسلوب وهي التي وفرت أحسن وسيلة لدراسة هذه الحركة. و يبدأ التحرك الأميباني نموذجيًا ببروز قدم كاذبة بعيدة عن جسم الخلية ثم تتلو ذلك حركة باقي الجسم باتجاهها حيث يتحرك غشاء هذه النهاية من الخلية إلى الأمام باستمرار ويتبعه أيضًا باستمرار غشاء النهاية المقابلة للخلية عندما تتحرك هي الأخرى.
التحرك الأميباني للخلايا
إن الأساس العام للحركة الأميبانية هو أنها تتولد في الأساس من الالتفاظ الخلوي ( الإيماس ) المستمر الذي يولد غشاءً خلويًا جديدًا عند الحافة المتقدمة من القدم الكاذبة والالتقام الخلوي المستمر للغشاء في قسمي الخلية الوسطي والخلفي. و هناك تأثير آخر ضروري لحركة الخلية للأمام ألا وهو التصاق القدم الكاذبة بالأنسجة المحيطة بحيث تصبح مثبتة في موقعها المتقدم بينما يسحب باقي جسم الخلية للأمام نحو نقطة الالتصاق. ويتولد هذا الالتصاق بالبروتينات المستقبلة التي تبطن داخل حويصلات الالتفاظ الخلوي. وعندما تصبح هذه الحويصلات جزءًا من غشاء القدم الكاذبة فإنها تتفتح وينقلب داخلها للخارج وتبرز عند ذاك المستقبلات للخارج أيضًا لتلامس ربائط ligands الأنسجة المحيطة بها. وتسمى إجدى هذه الربائط المهمة بصورة خاصة الفبرونكتين fibrionectin، وهي المرتبطة بالألياف الكُلاجينية لألياف الأنسجة [43] . و عند النهاية المقابلة للخلية تجذب الفعالية الالتقامية الخلوية المستقبلات بعيدًا عن ربائطها لتكون حويصلات الالتقام الخلوي ؛ ومن ثم تجري هذه الحويصلات داخل الخلية نحو نهاية قدمها الكاذبة حيث تستعمل لتكوين غشاء جديد آخر للقدم الكاذبة. و إحدى الحقائق المجهولة في عملية الحركة الأميبانية هي مصدر الطاقة التي تسبب جريان الحويصلات من نهاية الالتقام الخلوي نحو ذروة القدم الكاذبة في الخلية. ومن الممكن أن يتسبب جزء من ذلك في تقلص خيوط الأكتين والميوزين في الهيولى الظاهرة للخلية فيولد ذلك تقلص مؤخرتها ويدفع ذلك عمليًا الحويصلات والهيولى نحو نهاية القدم الكاذبة.
أنواع الخلايا التي تتحرك تحركًا أميبانيًا
إن أكثر الخلايا التي تتحرك تحركًا أميبانيًا في الجسم هي خلايا الدم البيضاء التي تتحرك خارجة من الدم إلى الأنسجة على شكل بلعميات أو بُلَيْعمات. ولكن هناك أنواع عديدة أخرى من الخلايا التي تتمكن من التحرك الأميباني في بعض الظروف الخاصة ؛ فمثلًا تتحرك الأرومات الليفية fibroblasts تحركًا أميبانيًا إلى أية منطقة مخربة في الجسم لتساعد في ترميم الضرر اللاحق بها. وبالرغم من أن بعض خلايا الجلد الإنتاشية هي خلايا لاطئة تمامًا لكنها تتحرك عادة نحو المناطق المقطوعة من الجلد لترميمها [44]. و أخيرًا فإن التحرك الخلوى مهم بصورة خاصة في تطوير الجنين لأن الخلايا الجنينية غالبًا ما تهاجر إلى مسافات طويلة من مواقع بدئية الأصل إلى مناطق جديدة متكونة في الجنين لتطوير بعض البنيات الخاصة فيه.
التحكم في التحرك الأميباني عن طريق الانجذاب الكيميائي
إن أهم عامل لبدء التحرك الأميباني في العادة هو العملية التي تسمى الانجذاب الكيميائي chemotaxis الذي يتولد من ظهور بعض المواد الكيميائية في الأنسجة والتي تسمى مواد الانجذاب الكيميائي. وتتحرك معظم الخلايا التي تتحرك تحركًا أميبانيًا نحو مصدر مواد الانجذاب الكيميائي؛ ويسمى هذا الانجذاب الكيميائي الموجب. كما أن بعض الخلايا تتحرك بعيدًا عن مصدر مواد الانجذاب الكيميائي ويسمى ذلك الانجذاب الكيميائي السالب. ولكن الكيفية التي يتحكم بها الانجذاب الكيميائي باتجاه التحرك الأميباني هي غير مؤكدة، ولكن المعروف أن حهة الخلية المعرضة لدرجة أكبر إلى مواد الانجذاب الكيميائي تولد تغييرات غشائية تؤثر على بروز الأقدام الكاذبة من الخلية.
الأهداب والحركات الهدبية للخلايا
هناك نوع ثان من الحركات الخلوية وهي الحركة الهدبية ciliary movement. وهي حركة سوطية الشكل للأهداب على سطح الخلية. وتحدث هذه الحركات في موقعين محددين في جسم الإنسان هما:
- على السطوح الداخلية للمجاري التنفسية
- على السطوح الداخلية لأنبوبة قناة فالوب في الجهاز التناسلي للأنثى.
و تسبب الحركات السوطية للأهداب في تجويف الأنف والمجاري التنفسية السفلى حركة طبقة المخاط الموجودة بها بسرعة 1 سم / دقيقة نحو البلعوم. وتنظف المجاري التنفسية بهذه الطريقة من المخاط ومن أية مواد [45] أخرى تتعلق بها. وتسبب الأهداب في أنبوبة الرحم ( قناة فالوب ) حركة بطيئة للسائل من فوهته نحو تجويف الرحم. وتنقل حركة السائل هذه البويضة من مبيض الأنثى إلى رحمها.
و يدل مظهر الأهداب على وجود شعرة مقوسة ذات نهاية مدببة تبرز على بعد 2 - 4 ميكرونات من سطح الخلية. ويبرز العديد من هذه الأهداب من سطح الخلية الواحدة فيوجد مثلًا حوالي 200 هدب على سطح كل خلية طلائية موجودة في مجرى الجهاز التنفسي. وتغطى الأهداب عادة ببروز من غلاف الخلية، وهو عادة ما يكون مسنودًا بأحد عشر نبيبًا مجهريًا وبتسعة نبيبات مزدوجة موضوعة حول محيطه ونبيبين مفردين في مركزه، وذلك عند أخذ مقطع عرضي في الأهداب. و يمثل كل هدب نموًا خارجيًا لبنية تقع تحت غشاء الخلية مباشرة تسمى الجسم القاعدي للأهداب basal body [46]. و يكون سوط الحيوان المنوي sperm شبيه بالأهداب، وله في الواقع نفس نمط بنيته ونفس نمط آلية تقلصه، ولكنه أكثر طولًا ويتحرك بموجات شبه جيبية quasi - sinusoidal بدلًا من الحركات السوطية whiplike. و تظهر حركة الأهداب بأنها تتحرك للأمام بضربات سريعة، بما يعادل 10 - 20 ضربة في الثانية تقريبًا، وتنثني بشدة عند نقطة البروز من سطح الخلية، ومن ثم تتحرك الأهداب ببطء شديد بطريقة الحركة السوطية. وتقوم الحركات الأهداب السريعة للأمام بدفع السائل المجاور لسطح الخلية باتجاه حركة الأهداب السريعة، ولكن ليس هناك أي تأثير لحركاتها البطيئة بالاتجاه المعاكس على حركة السائل. ونتيجة لذلك يفع السائل باستمررا بالاتجاه الأمامي لضربات الأهداب. ولما كانت لمعظم الخلايا الهدبية أعداد كبيرة من الأهداب على سطحها، ولما كانت كل الأهداب موجهة باتجاه واحد لذلك تصبح هذه الحركات فعالة جدًا في تحريك السائل من أحد أقسام السطح إلى أقسامه الأخرى[47].
آلية الحركة الهدبية
بالرغم من عدم وضوح كل نواحي الحركة الهدبية لكن المعروف عنها هو الآتي:
- ترتبط النبيبات المزدوجة التسعة والنبيبين المفردين مع بعضها بارتباطات عابرة مكونة من معقد بروتيني، ويسمى هذا المعقد المكون من النبيبات والروابط البروتينية العابرة الخيط المحوري axoneme.
- تستمر الأهداب بالتحرك تحت الظروف المناسبة حتى بعد إزالة الغشاء منه وتدمير عناصره الأخرى الموجودة بجانب الخيط المحوري [48].
- هناك شرطان ضروريان لاستمرار ضرب الخيط المحوري بعد إزالة بنيات الأهداب الأخرى ؛ وهما:
- وجود الـ ATP
- وجود ظروف أيونية مناسبة تشمل بصورة خاصة التركيز المناسب للماغنسيوم والكالسيوم.
- تنزلق نبيبات الحافة الأمامية للأهداب المنثنية للأمام نحو قمة الأهداب بينما تبقى نبيبات الحافة الخلفية في موضعها.
- تبرز ثلاثة أذرع بروتينية مكونة من البروتين داينين dynein الذي يمتلك فعالية إنزيم الـ ATPase من كل واحد من مجموعة النبيبات المحيطة لتمر نحو المجموعة التالية لها [49].
فمع تطور لمعرفة بكل هذه الحقائق الأساسية أصبح من الممكن معرفة أن إطلاق الطاقة من الـ ATP الملامسة لإنزيم الـ ATPase الموجود على أذرع الداينين يسبب زحف الأذرع على سطح زوج النبيبات المجاورين لها، فإذا زحفت النبيبات الأمامية للخارج وبقيت النبيبات الخلفية ثابتة أدى ذلك طبعًا إلى الانثناء. و لا تعرف حتى الآن الطريقة التي تحكم بها تقلصات الأهداب ؛ ولكن المعروف هو أن أهداب الخلايا الشاذة وراثيًا لا تحوي النبيبين المركزيين المفردين ولا تتحرك هذه الأهداب أبدًا. ولذلك يفترض بأن بعض الإشارات - ولربما كانت إشارة كهربية كيميائية - تنتقل في هذين النبيبين لتنشيط أذرع الداينين[50].
علماء الخلية
- روبرت براون: أول من شاهد كرة صغيرة داخل الخلية النباتية واطلق عليها اسم النواة.
- فيرشو: اثبت أن الخلايا تأتي من الانقسام غير المباشر للخلايا.
مرادفات مكونات الخلايا
هنا أسماء مكونات الخلية مع أسماء لمرادفاتها
| المُسمَّى الرئيس | مُسميات أخرى |
|---|---|
| جدار خلوي | لا يوجد |
| مُريكز | جسم مركزي |
| غشاء خلوي | غشاء بلازمي |
| صانعة | بلاستيد |
| صانعات يخضورية | بلاستيدات خضراء |
| صانعات مُلونة | بلاستيدات ملونة |
| صانعات عديمة اللون | بلاستيدات عديمة اللون |
| نواة | لا يوجد |
| نوية | لا يوجد |
| ريبوسوم | رايبوسوم، أو جسيم ريبي |
| حويصلات | لا يوجد |
| شبكة هيولية باطنة (الأمر ذاته بالنسبة للشبكة الهيولية الباطنة الملساء أو الخشنة وبالنسبة للرابطة الهيولية) | شبكة هيولية داخلة، أو شبكة إندوبلازمية، أو شبكة بلازمية داخلية |
| جهاز جولجي | جسيم جولجي، أو شبكة جولجي |
| حبيبيات خيطية | متقدرات، أو مصورات حيوية، أو ميتوكندريون، أو ميتوكندريا |
| فجوة | لا يوجد |
| الجِبْلة | هيولى الخلية، أو سيتوبلازم |
| اليحلول | جسم (جسيم) حال أو محلل أو هاضم أو ليسوسوم |
| أنيبيب دقيق | لا يوجد |
| ثقب نواة | لا يوجد |
| جسيم تأكسدي | لا يوجد |
| محفظة | لا يوجد |
| أسواط | لا يوجد |
| أهداب | لا يوجد |
| الحمض النووي الريبوزي المنقوص الأكسجين | دي إن أي |
| الحمض النووي الريبوزي | آر إن أي |
مقالات ذات صلة
- خلية حيوانية
- فيزيولوجيا الخلية
- زرع الخلايا
- سمية الخلايا
- خط الخلايا الجذعية
- المجهر
- مناعة داخلية
- متعددة الخلايا
- مستقبل ضوء
- حبيبة (أحياء)
- تحلل استماتي
مراجع
- Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. . مؤرشف من الأصل في 1 نوفمبر 2018.
- Karp, Gerald (19 October 2009). Cell and Molecular Biology: Concepts and Experiments. John Wiley & Sons. صفحة 2. .
Hooke called the pores cells because they re- minded him of the cells inhabited by monks living in a monastery.
- "Cell". Online Etymology Dictionary. مؤرشف من الأصل في 4 يوليو 201731 ديسمبر 2012.
- Ménétret JF, Schaletzky J, Clemons WM, et al. (December 2007). "Ribosome binding of a single copy of the SecY complex: implications for protein translocation". Mol. Cell. 28 (6): 1083–92. doi:10.1016/j.molcel.2007.10.034. ببمد 18158904
- Microbiology : Principles and Explorations By Jacquelyn G. Black
- Kimball's Biology pages, Cell Membranes نسخة محفوظة 20 فبراير 2018 على موقع واي باك مشين.
- Singleton P (1999). Bacteria in Biology, Biotechnology and Medicine (الطبعة 5th). New York: Wiley. .
- Lodish H, Berk A, Zipursky LS, et al. (2004). Molecular Cell Biology (الطبعة 4th). New York: Scientific American Books. . مؤرشف من في 16 ديسمبر 2019.
- Jesse Gray; Shana Groeschler; Tony Le; Zara Gonzalez (2002). "Membrane Structure". Davidson College. مؤرشف من الأصل في 13 أكتوبر 201611 يناير 2007.
- Doherty GJ, McMahon HT (2008). "Mediation, Modulation and Consequences of Membrane-Cytoskeleton Interactions". Annual Review of Biophysics. 37: 65–95. doi:10.1146/annurev.biophys.37.032807.125912. PMID 18573073. مؤرشف من الأصل في 08 مارس 2020.
- Budin, Itay; Devaraj, Neal K. (December 29, 2011). "Membrane Assembly Driven by a Biomimetic Coupling Reaction". Journal of the American Chemical Society. 134 (2): 751–753. doi:10.1021/ja2076873. مؤرشف من الأصل في 31 مايو 201518 فبراير 2012.
- Alberts B, Johnson A, Lewis J, et al. (2002). Molecular Biology of the Cell (الطبعة 4th). New York: Garland Science. . مؤرشف من الأصل في 14 فبراير 2009.
- Staff (January 25, 2012). "Chemists Synthesize Artificial Cell Membrane". علم يوميا. مؤرشف من الأصل في 5 نوفمبر 201818 فبراير 2012.
- cell nucleus Dr.Jastrow's EM-Atlas - تصفح: نسخة محفوظة 31 ديسمبر 2016 على موقع واي باك مشين.
- The cell nucleus and its DNA on a human scale – I | Chemiotics II - تصفح: نسخة محفوظة 13 أكتوبر 2016 على موقع واي باك مشين.
- Thiry M, Lafontaine DL (2005). "Birth of a nucleolus: the evolution of nucleolar compartments". Trends Cell Biol. 15 (4): 194–9. doi:10.1016/j.tcb.2005.02.007. PMID 15817375. as PDF - تصفح: نسخة محفوظة 23 أكتوبر 2013 على موقع واي باك مشين.
- Beven AF, Lee R, Razaz M, Leader DJ, Brown JW, Shaw PJ (1 June 1996). "The organization of ribosomal RNA processing correlates with the distribution of nucleolar snRNAs". J. Cell. Sci. 109 (6): 1241–51. PMID 8799814.
- Jones, Daniel (2003) [1917], Peter Roach, James Hartmann and Jane Setter (المحررون), English Pronouncing Dictionary, Cambridge: Cambridge University Press,
- Walter, Peter. "Peter Walter's short talk: Unfolding the UPR". iBiology. مؤرشف من الأصل في 12 يوليو 2017.
- "Endoplasmic Reticulum (Rough and Smooth)". مؤرشف من الأصل في 24 مارس 201921 نوفمبر 2015.
- Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. . مؤرشف من الأصل في 1 نوفمبر 2018.
- "mitochondria". قاموس علم اشتقاق الألفاظ. مؤرشف من الأصل في 2 يوليو 2017.
- Voet, Donald; Judith G. Voet; Charlotte W. Pratt (2006). Fundamentals of Biochemistry, 2nd Edition. John Wiley and Sons
- Campbell, Neil A (1996). Biology (الطبعة 4). Menlo Park, CA: Benjamin/Cummings. صفحات 122, 123. .
- William G. Flynne (2008). Biotechnology and Bioengineering. Nova Publishers. صفحات 45–. . مؤرشف من الأصل في 3 يناير 202013 نوفمبر 2010.
- del Río LA, Sandalio LM, Palma JM, Bueno P, Corpas FJ (Nov 1992). "Metabolism of oxygen radicals in peroxisomes and cellular implications". Free Radical Biology & Medicine. 13 (5): 557–80. doi:10.1016/0891-5849(92)90150-F. PMID 1334030.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). "Chapter 12: Peroxisomes". Molecular Biology of the Cell (الطبعة Fourth). New York: Garland Science. . مؤرشف من الأصل في 5 فبراير 2009.
- Kuehnel W (2003). Color Atlas of Cytology, Histology, & Microscopic Anatomy (الطبعة 4th). Thieme. صفحة 34. .
- Mindell JA (2012). "Lysosomal acidification mechanisms". Annual Review of Physiology. 74 (1): 69–86. doi:10.1146/annurev-physiol-012110-142317. PMID 22335796.
- Ishida Y, Nayak S, Mindell JA, Grabe M (Jun 2013). "A model of lysosomal pH regulation". The Journal of General Physiology. 141 (6): 705–20. doi:10.1085/jgp.201210930. PMC . PMID 23712550.
- Lodish H, et al. (2000). Molecular cell biology (الطبعة 4th). New York: Scientific American Books. . مؤرشف من في 16 ديسمبر 2019.
- J. Robinson (ed) The Oxford Companion to Wine Third Edition p 54 Oxford University Press 2006
- "Microtubules" en. مؤرشف من الأصل في 19 نوفمبر 201608 مارس 2020.
- Vale RD (Feb 2003). "The molecular motor toolbox for intracellular transport". Cell. 112 (4): 467–80. doi:10.1016/S0092-8674(03)00111-9. PMID 12600311.
- Wayne, R. 2009. Plant Cell Biology: From Astronomy to Zoology. Amsterdam: Elsevier/Academic Press, p. 165. نسخة محفوظة 15 ديسمبر 2016 على موقع واي باك مشين.
- Alberts, Johnson, Lewis, Raff, Roberts, Walter: "Molecular Biology of the Cell", Fourth Edition, Copyright 2002 P.748
- The Immune System, Peter Parham, Garland Science, 2nd edition
- ( كتاب إلكتروني PDF ) https://web.archive.org/web/20170311163850/http://www.colorado.edu/intphys/iphy3700/vitCHemila92.pdf. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 11 مارس 2017.
- Parham, P. (2005). The Immune System," Garland Science Publishing, New York, NY.
- Definition: opsonization from Online Medical Dictionary - تصفح: نسخة محفوظة 16 سبتمبر 2008 على موقع واي باك مشين.
- rosenbaum, JL; Witman G (November 2002). "Intraflagellar transport". Nature Reviews Molecular Cell Biology. 3 (11): 813–25. doi:10.1038/nrm952. PMID 12415299.
- Scholey, JM (2008). "Intraflagellar transport motors in cilia: Moving along the cell's antenna". Journal of Cell Biology. 180: 23–29. doi:10.1083/jcb.200709133. PMC . PMID 18180368.
- Alberts, Bruce. "Molecular Biology of the Cell". مؤرشف من الأصل في 8 مارس 2019.
- Hao, L; Thein M; Brust-Mascher I; Civelekoglu-Scholey G; Lu Y; Acar S; Prevo B; Shaham S; Scholey JM (2011). "Intraflagellar transport delivers tubulin isotypes to sensory cilium middle and distal segments". Nature Cell Biology. 13 (7): 790–798. doi:10.1038/ncb2268. PMC . PMID 21642982.
- Johnson, KA; Rosenbaum, JL (1992). "Polarity of flagellar assembly in Chlamydomonas". Journal of Cell Biology. 119 (6): 1605–1611. doi:10.1083/jcb.119.6.1605. PMC . PMID 1281816.
- Pugacheva EN, Jablonski SA, Hartman TR, Henske EP, Golemis EA (June 2007). "HEF1-dependent Aurora A activation induces disassembly of the primary cilium". Cell. 129 (7): 1351–63. doi:10.1016/j.cell.2007.04.035. PMC . PMID 17604723.
- Of cilia and silliness (more on Behe) - The Panda's Thumb - تصفح: نسخة محفوظة 07 مايو 2016 على موقع واي باك مشين.
- "Functional characterisation of the centrosomal protein Cep170". مؤرشف من الأصل في 29 يوليو 2017.
- Ishikawa H, Kubo A, Tsukita S, Tsukita S (May 2005). "Odf2-deficient mother centrioles lack distal/subdistal appendages and the ability to generate primary cilia". Nature Cell Biology. 7 (5): 517–24. doi:10.1038/ncb1251. PMID 15852003.
- "Ciliary Rootlet". Gene Ontology. مؤرشف من الأصل في 25 مارس 202013 يونيو 2012.