فيروس العوز المناعي البشري (Human Immunodeficiency Virus، يعرف اختصاراً: HIV) هو فيروس يهاجم جهاز المناعة بالجسم ويعطل عمله ويؤدي إلى الإيدز، مما يجعل الإنسان ضعيفاً وبدون أي قوة دفاعية ضد أي مرض، لأنه فقد حماية جهاز مناعة جسمه له، وهنا يتعرض للإصابة بأنواع كثيرة وخطيرة، من الأمراض والسرطانات التي تسمى الأمراض الانتهازية، لأنها انتهزت فرصة عدم قدرة جسم الإنسان عن الدفاع عن نفسه فهاجمته.
| فيروس العوز المناعي البشري | |
|---|---|
 رسم تخطيطي لفيروس العوز المناعي البشري.
| |
| الوبائيات | |
| انتشار المرض | 0.0075 |
| التاريخ | |
| وصفها المصدر | موسوعة لاروس الإلكترونية[1] |
هذا الفيروس ضعيف جداً لا يعيش خارج جسم الإنسان، ولهذا لا يمكن الإصابة به خارج جسم الإنسان بمعنى لا يمكن الإصابة به بالملامسة أو باستعمال أدوات الشراب أو الأكل...إلخ بل ينتقل عن طريق الدم أو عبر الرضاعة الطبيعية أو الإفرازات الجنسية من الجنسين أثناء العلاقة الجنسية.
وفى حالة وصوله إلى داخل جسم الإنسان قد يبقى في حالة كمون، ربما لسنوات، يحدث خلالها تحطيم لجهاز مناعة الإنسان. وهنا نرى أن بعض الناس الذين يظهرون بصحة جيدة إلى الآن هم ينقلون هذا الفيروس لغيرهم وهم لا يعلمون.
وما زال العلماء لا يعرفون كم هي النسبة المئوية % للمصابين بهذا الفيروس، الذين سوف تظهر عليهم الحالة المرضية ولا يعرفون ما هو الزمن الذي يأخذه فيروس العوز المناعي البشري من لحظة دخوله جسم الإنسان إلى أن تظهر عليه أول علامات الإصابة بالإيدز.
ولكن العلماء أيضاً وجدوا أنه بالعلاج المناسب، قليل من المصابين يصلوا إلى حالة الإيدز. ويعتمدوا الآن أيضاً أن كثيراً من المصابين بفيروس العوز المناعي البشري يمكنهم الحياة بالأمراض التي يسببها هذا الفيروس لعدة سنوات وربما في وقت ما في المستقبل يصبح أمراض فيروس العوز المناعي البشري كمثل مرض السكري مرض مزمن يمكن السيطرة عليه.
علم الفيروسات
التصنيف
- مقالة مفصلة: الأنواع الفرعية لفيروس العوز المناعي البشري
| النوع | الضراوة | الإعداء | الانتشار | الأصل المستنتح |
|---|---|---|---|---|
| فيروس الإيدز-1 | عالي | عالي | عالمي | الشيمبانزي الشائع |
| فيروس الإيدز-2 | منخفض | منخفض | غرب إفريقيا | المنجبي الأسخم |
ينتمي فيروس الإيدز إلى جنس الفيروسات البطيئة،[2] وهو جزء من عائلة الفيروسات الراجعة.[3] للفيروسات البطيئة العديد من الأشكال البنيوية والخصائص البيولوجية المشتركة. تُصاب العديد من الأنواع بالفيروسات البطيئة التي تتميز بطول فترة حضانتها وبكونها مسؤولة عن أمراض طويلة الأمد.[4] الفيروسات البطيئة هي فيروسات رنا مفرد السلسلة موجب الاتجاه مغلفة. عند دخولها إلى الخلية الهدف، يحوّل (ينسخ عكسيا) جينوم الرنا الفيروسي إلى سلسلة دنا مزدوجة بواسطة إنزيم مشفٍَّر في جينوم الفيروس وهو الناسخ العكسي الذي يكون موجودا في الفيروس إلى جانب الجينوم الفيروسي. بعد ذلك يُنقل الدنا الفيروسي الناتج إلى نواة الخلية ويُدمج مع الدنا الخلوي بواسطة إنزيم مشفّرٍ في جينوم الفيروس هو الدامج وعوامل مرافقة من الخلية المضيفة.[5] حين يتم دمجه، يمكن أن يصبح الفيروس كامنا لمدة غير محددة من الزمن وذلك لتجنب اكتشافه هو والخلية المضيفة من قبل الجهاز المناعي.[6] يمكن أن يبقى فيروس الإيدز في سبات في جسم الإنسان حتى 10 سنوات بعد الإصابة الأولية، وخلال هذه الفترة لا يُسبب الفيروس أعراضا. ويمكن أن يُنسخ الدنا الفيروسي المدمج وينتج منه جينومات رنا وبروتينات فيروسية جديدة -باستخدام موارد الخلية المضيفة وماكينة النسخ فيها- يتم تجميعها وتحريرها من الخلية كفيروسات جديدة ستبدأ بدورها دورة تضاعف من جديد.
حُددت خصائص نوعين من فيروس الإيدز: فيروس الإيدز-1 وفيروس الإيدز-2. فيروس الإيدز-1 هو الفيروس الذي اكتُشِف في البداية وسُمي بالاسمين: الفيروس المرتبط بتضخم العقد اللمفية (LAV) والفيروس الأليف للمفاويات التائية البشرية 3 (HTLV-III). فيروس الإيدز-1 أشد ضراوة وأكثر إعداءً من فيروس الإيدز-2،[7] ويسبب معظم الإصابات بفيروس الإيدز عالميا. فيروس الإيدز-2 أقل إعداء مقارنة بفيروس الإيدز-1، ويعنى ذلك أن عددا أقل من المعرضين لفيروس الإيدز-2 ستتم إصابتهم في كل تعرض. بسبب قدرته الضعيفة نسبيا في الانتشار، فيروس الإيدز-2 محصور بشكل كبير في غرب إفريقيا.[8]
البنية والجينوم

- مقالة مفصلة: بنية وجينوم فيروس الإيدز
فيروس الإيدز مختلف في البنية عن بقية الفيروسات الراجعة، فهو كروي الشكل تقريبا[9] وقطره حوالي 120 نانومتر، أصغر بحوالي 60 مرة من خلية الدم الحمراء.[10] يتكون الفيروس من نسختي سلسلة رنا مفردة موجبة الاتجاه تشفران جينات الفيروس التسعة وهما محاطتين بقفيصة مخروطية تتكون من ألفي نسخة من البروتين الفيروسي P24.[11] سلسلة الرنا المفردة مرتبطة بإحكام مع بروتينات القفيصة المنواة P7، والإنزيمات المطلوبة لنسخ ودمج وتجميع الفيروسات الجديدة مثل الناسخ العكسي، البروتيازات، الريبونوكليازات والدامج. يحيط مطرس مكون من البروتين الفيروسي p17 بالقفيصة لضمان سلامة الفيروس.[11]
المطرس بدوره محاط بغلاف فيروسي مكون من ليبيد ثنائي الطبقة مأخوذٍ من الغشاء البلازمي الخاص بالخلية المضيفة البشرية حين يتشكل جسيم الفيروس الجديد ويتبرعم منها. يحتوي الغلاف الفيروسي على بروتينات من الخلية المضيفة ونسخا قليلة نسبيا من بروتين غلاف فيروس الإيدز،[11] الذي يتكون من قبعة مكونة من ثلاث جزيئات تعرف بالبروتين السكري 120 (gp 120) وجذع يتكون من ثلاث جزيئات من البروتين السكري gp41 يعمل على إرساء هذه البنية في الغلاف الفيروسي.[12][13] يسمح بروتين الغلاف الفيروسي المشفَّر بواسطة جين فيروس الإيدز Env للفيروس بالارتباط بالخلية الهدف ودمج الغلاف الفيروسي مع الغشاء البلازمي للخلية وتحرير المحتوى الفيروسي داخل الخلية والشروع في دورة التكاثر والإعداء.[12]
لكونه البروتين الفيروسي الوحيد على سطح الفيروس، بروتين الغلاف هدف رئيسي لمجهودات تطوير لقاح لفيروس الإيدز.[14] أزيد من نصف كتلة حسكة الغلاف الثلاثية هي غليكانات مرتبطة بالنيتروجين. كثافة الغليكانات عالية لأنها تقي البروتين الموجود أسفلها من تحييده بواسطة الأجسام المضادة، وهو أحد أكثر الجزيئات المعروفة غلكزةً والكثافة عالية لدرجة تمنع عملية النضوج العادية للغليكانات أثناء التخلق في الشبكة الإندوبلازمية وجهاز غولجي.[15][16] معظم الغليكانات نتيجة لذلك هي غليكانات متوقفة غير ناضجة غنية بالمانوز لا تتواجد عادة في البروتينات السكرية البشرية التي تُفرز أو الموجودة على سطح الخلية.[17] المعالجة غير الاعتيادية والكثافة العالية تعني أن تقريبا جميع الأجسام المضادة التي تم تحديدها (من مجموعة جزئية من المرضى المصابيين بالفيروس منذ شهور أو سنوات) ترتبط بغليكان الغلاف هذه أو تتأقلم لتتكيف معها.[18]
تم تحديد البنية الجزيئية للحسكة الفيروسية بواسطة علم البلورات السيني[19] والمجهرية فائقة البرودة.[20] هذه التقدمات في البيولوجيا البنيوية كانت ممكنة بفضل تطوير نسخ مستقرة مأشوبة من الحسكة الفيروسية عبر إضافة رابطة ثنائي كبريتيد بين الوحدات وطفرة من إيزوليوسين إلى برولين (تبديل جذري لحمض أميني) في البروتين السكري gp41.[21] نسخ الحسكة المطورة هذه المسماة الـSOSIP ثلاثي القسمات لا تحاكي الخصائص المستضدية للحسكة الفيروسية الطبيعية وحسب، بل تحتوي تقريبا على نفس كمية الغليكانات كما هو الحال في الفيروس الطبيعي.[22] الحسكات الفيروسية المأشوبة ثلاثية القسمات هي مرشحات لقاحات واعدة لأنها تُظهر حاتمات غير مسببة للتحييد أقل من البروتين السكري المأشوب أحادي القسمات gp120، الذي يعمل على كبت استهداف الجهاز المناعي للحاتمات.[23]

يتكون جينوم الرنا على الأقل من سبع معالم بنيوية هي (LTR وTAR وRRE وPE وSLIP وCRS وINS) وتسع جينات (gag وpol وenv وtat وrev وnef وvif وvpr وvpu وأحيانا جين عاشر tev وهو اندماج للجينات tat وenv وrev) تشفر 19 بروتينا. تحتوي ثلاثة من هذه الجينات -gag وpol وenv- على المعلومات المطلوبة لتخليق البروتينات البنيوية لبناء فيروسات جديدة.[11] على سبيل المثال يشفِّر الجين env بروتينا يسمى gp160 يُقص بواسطة بروتياز خلوي لإنتاج بروتيني gp120 وgp41. الجينات الستة الباقية: tat وrev وnef وvif وvpr وvpu (أو vpx في حالة فيروس الإيدز-2) هي جينات تنظيمية تنتج بروتينات تتحكم في قدرة فيروس الإيدز على إصابة الخلايا وإنشاء نسخ جديدة من الفيروس (التضاعف) أو إحداث المرض.[11]
بروتينَيْ tat (p16 وp14) هما منشطان-مفروق نسخيان لمحفز LTR ويعملان عبر الارتباط بعنصر الرنا TAR . يمكن أن يُعالَج TAR إلى جزيئات رنا ميكروي تقوم بتنظيم جينَيْ الاستماتة ERCC1 وIER3.[24][25] بروتين Rev (p19) له دور كذلك في تمكيك (نقل وحماية) جزيئات الرنا من النواة والسيتوبلازم عبر الارتباط بعنصر الرنا RRE. يمنع بروتين vif (p23) عمل APOBEC3G (وهو بروتين خلوي يقوم بنزع أمين السيتيدين وتحويله إلى يوريدين في سلسلة الدنا المفردة الفيروسية و/أو التدخل في عمل الناسخ العكسي[26]). يوقف بروتين Vpr (p14) انقسام الخلية في نقطة التحقق G2/M. بروتين nef (p27) ينظم بالإنقاص بروتين CD4 (المستقبل الرئيسي للفيروس)، وكذلك القسم 1 والقسم 2 من معقد التوافق النسيجي الكبير.[27][28][29]
يتآثر بروتين Nef كذلك مع نطاق SH3. بروتين vpu (p16) يؤثر على على تحرير الفيروسات الجديدة من الخلايا المصابة.[11] تسمى نهايتا كل سلسلة من رنا فيروس الإيدز بالتكرار الطرفي الطويل (LTR)، وتعمل مناطقٌ في التكرار الطرفي الطويل كمفاتيح تحويل للتحكم في إنتاج الفيروسات الجديدة ويمكن أن تُثارَ بواسطة بروتينات من الفيروس أو من الخلية المضيفة. عنصر Psi له دور في تجميع الجينوم الفيروسي ويتم تنظيمه بواسطة بروتينات gag وrev. عنصر SLIP (TTTTTT) له دور في انزياح الإطار في إطار القراءة gag-pol المطلوب لإنتاج بروتين pol وظيفي.[11]
الانتحاء

يشير مصطلح انتحاء فيروسي إلى أنواع الخلايا التي يمكن أن يصيبها الفيروس. يمكن لفيروس الإيدز أن يصيب مجموعة متنوعة من الخلايا المناعية مثل الخلايا التائية المساعدة (CD4+)، البلعميات، والخلايا الدبقية الصغيرة. يتوسَّط دخولَ فيروس الإيدز إلى البلعميات والخلايا التائية المساعدة تآثرُ البروتين السكري الخاص بغلاف الفيروس (gp120) مع جزيء كتلة التمايز 4 الموجود على غشاء الخلية المستهدفة وكذلك مع مستقبلات الكيموكين المساعدة.[12][30]
تستخدم سلالات فيروس الإيدز-1 المنتحية للبلعميات، أو غير المحدِثة للملتحم الخلوي (NSI، تسمى الآن فيروسات R5[31]) مستقبل الكيموكين-بيتا -CCR5- للدخول وبذلك تكون قادرة على التضاعف في كل من البلعميات والخلايا التائية المساعدة.[32] يُستخدم المستقبل المساعد CCR5 تقريبا بواسطة جميع فيروسات الإيدز-1 المعزولة بغض النظر عن النوع الجيني الفرعي للفيروس. بالفعل، تلعب البلعميات دورا مفتاحيا في جوانب حاسمة عديدة من الإصابة بفيروس الإيدز. حيث يظهر بأنها أول الخلايا المصابة بالفيروس وربما مصدر إنتاج فيروسات الإيدز حين تصبح الخلايا التائية المساعدة مستنفذة لدى المريض. البلعميات والخلايا الدبقية الصغيرة هي الخلايا التي يصيبها فيروس الإيدز في الجهاز العصبي المركزي. في الزوائد الأنفية ولوزات المرضى المصابين بالفيروس، تندمج البلعميات إلى خلايا عملاقة متعددة النوى تقوم بإنتاج كميات كبيرة من الفيروس.
تتضاعف سلالات فيروس الإيدز-1 المنتحية للخلايا التائية، أو السلالات المحدِثة للالتحام الخلوي (SI تسمى الآن فيروسات X4[31]) في الخلايا التائية المساعدة الأساسية وكذلك في البلعميات، وتستخدم مستقبل الكيموكين-ألفا -CXCR4- للدخول.[32][33][34]
يُعتقد أن سلالات فيروس الإيدز-1 ثنائية الانتحاء هي سلالات انتقالية من فيروس الإيدز-1 وبالتالي تكون قادرة على استخدام كل من CCR5 وCXCR4 كمستقبلات مساعدة للدخول إلى الخلية.
يثبط الكيموكين-ألفا SDF-1 -وهو ربيطة للمستقبل CXCR4- تضاعف فيروسات الإيدز-1 المعزولة المنتحية للخلايا التائية، ويقوم بذلك عبر تخفيض التعبير عن CXCR4 في سطح الخلايا التي يستهدفها الفيروس. فيروسات الإيدز-1 المعزولة المنتحية للبلعميات التي تستخدم المستقبل CCR5 فقط تسمى R5، أما التي تستخدم المستقبل CXCR4 فقط فتسمى X4، والتي تستخدم كلا المستقبلين تسمى X4R5. استخدام المستقبلات المساعدة وحدها لا يفسر الانتحاء الفيروسي، فما كل فيروسات R5 قادرة على استخدام CCR5 الموجود على البلعميات لإحداث إصابة فعالة[32] ويمكن كذلك لفيروس الإيدز إصابة أنواع فرعية من الخلايا المتغصنة النخاعية،[35] التي ربما تكون خزانا يحافظ على الإصابة حين تنخفض أرقام الخلايا التائية المساعدة إلى مستويات دنيا قياسية.
بعض الأشخاص مقاومون لبعض سلالات فيروس الإيدز.[36] على سبيل المثال: الأفراد الذين لديهم طفرة CCR5-Δ32 مقاومون للإصابة بفيروس R5 لأن الطفرة تجعل فيروس الإيدز غير قادر على الارتباط بهذا المستقبل المساعد، وهو ما يخفض قدرته على إصابة الخلايا المستهدفة.
الجماع هو النمط الرئيسي لانتقال فيروس الإيدز، وكلا السلالات X4 و R5 تتواجد في السائل المنوي، وهو ما يسمح للفيروس بالانتقال من الذكر إلى شريكه الجنسي. يمكن لفيروسات الإيدز إصابة العديد من الأهداف الخلوية والانتشار في كافة أنحاء الكائن. مع ذلك، تؤدي عملية انتقاءٍ إلى الانتقال الرئيسي لفيروس R5 عبر هذا النمط والمسار الجنسي.[37][38][39] في المرضى المصابين بالنوع الفرعي فيروس الإيدز-1 B، عادة ما يكون هنالك استبدالٌ للمستقبل المساعد في المرحلة المتقدمة من المرض وعادة ما تنتج مغايِرات منتحية للخلايا التائية يمكنها إصابة صنوفٍ من الخلايا التائية عبر CXCR4.[40] بعدها تتضاعف هذه المغايرات بمعدل أكبر وتشتد ضراوتها مسببة نفاذ الخلايا التائية السريع، وانهيار الجهاز المناعي وحدوث الإصابات الانتهازية التي تكون علامة لمرحلة متقدمة من مرض الإيدز.[41] يصاب المرضى المصابون بفيروس الإيدز بطيف واسع من العداوى الانتهازية والتي كانت سابقا تسبب إشكالات قبل بداية العلاج عالي الفاعلية بمضادات الفيروسات الراجعة (HAART)، مع ذلك، نفس الإصابات تم التبليغ عنها بين المرضى المصابين بفيروس الإيدز المفحوصين بعد الوفاة عقب بداية العلاجات بمضادات الفيروسات الراجعة.[42] لهذا أثناء فترة الإصابة، التأقلم الفيروسي لاستخدام مستقبل CXCR4 بدل CCR5 يمكن أن يكون خطوة رئيسية في استفحال وتطور مرض الإيدز. حددت عدة دراسات لمرضى مصابين بالفيروس الفرعي B أن بين 40-50% من مرضى الإيدز يمكنهم احتواء فيروسات من النمط الظاهري SI والنمط NSI.[43][44]
فيروس الإيدز-2 أقل إمراضا من فيروس الإيدز-1 وهو محصورٌ في انتشاره العالمي في غرب إفريقيا. تبني فيروس الإيدز-2 "للجينات الملحقة" ونمطه الأكثر اختلاطا في استخدام المستقبل المساعد (بما في ذلك عدم الاعتماد على CD4) يمكن أن يساعد الفيروس في تأقلمه لتجنب عوامل التقييد الخاصة بالمناعة الفطرية الموجودة في الخلايا المضيفة. التأقلم لاستخدام الماكينة الخلوية الطبيعية لتمكين انتقال العدوى وفاعليتها ساعد أيضا فيروس الإيدز على التضاعف لدى البشر. استراتيجية البقاء على قيد الوجود الخاصة بأي عامل إعداء هي أن لا يقتل المضيف، وإنما أن يصبح في النهاية كائنا معايشا. مع تحقيقها لإمراضية وضراوة منخفضة -بمرور الزمن- المغايرات الأكثر نجاحا في الانتقال سيتم اصطفاؤها.[45]
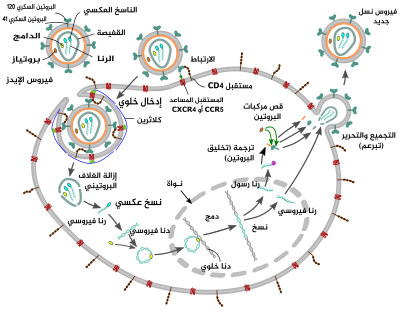
دورة التضاعف
الدخول إلى الخلية

يدخل فيروس الإيدز إلى البلعميات والخلايا التائية المساعدة عبر امتزاز (ارتباط) البروتينات السكرية على سطحه بالمستقبلات الموجودة على الخلية الهدف، يتبع ذلك اندماجٌ للغلاف الفيروسي مع الغشاء الخلوي للخلية المستهدفة وتحرير قفيصة فيروس الإيدز داخل الخلية.[46][47]
يبدأ الدخول إلى الخلية عبر تآثر مركب البروتين السكري ثلاثي القسمات (الحكسة gp160) الموجود على غلاف الفيروس مع كل من كتلة التمايز 4 (CD4) ومستقبل كيموكين مساعد (عادة إما CCR5 أو CXCR4، لكن يمكن أن تتآثر مستقبلات أخرى) الموجودان على سطح الخلية المستهدفة.[46][47] يرتبط Gp120 مع الإنتغرين α4β7 ويُنشِّط LFA-1 -وهو الأنتغرين المركزي الذي له دور في إنشاء مشابك فيروسية- الذي يسهل الانتشار الفعال لفيروس الإيدز-1 من خلية لأخرى.[48] تحتوي الحسكة gp160 على نطاقات ارتباط مع كل من كتلة التمايز 4 ومستقبلات الكيموكين.[46][47]
الخطوة الأولى لدمج الغشائين تتم عبر الارتباط عالي الألفة بين نطاق الارتباط ببروتين كتلة التمايز 4 الخاص بـgp120 وبروتين كتلة التمايز نفسه، عندما يرتبط هذان الأخيران، يخضغ gp120 لتغير هيئوي كاشفا نطاقات الارتباط بمستقبل الكيموكين الخاصة به وهذا يسمح لها بالتآثر مع مستقبل الكيموكين المستهدف.[46][47] يوفر هذا الارتباط المزدوج استقرارا أكبر ويسمح للنهاية الأمينية الخاصة بببتيد الدمج gp41 باختراق الغشاء الخلوي.[46][47] بعد ذلك تتآثر التسلسلات المتكررة في gp41 وHR1 وHR2 مسببة تغير بنية الجزء خارج الخلوي من gp41 إلى حلقة جذعية. تُقرِّب هذه البنية الحلقية غشائي الفيروس والخلية من بعضهما وتسمح باندماجهما ثم يلي ذلك دخول القفيصة الفيروسية إلى الخلية.[46][47]
بعد ارتباط فيروس الإيدز بالخلية المستهدفة، يدخل كل من الرنا الفيروسي ومختلف الإنزيمات بما في ذلك الناسخ العكسي، الدامج، الريبونوكلياز والبروتياز إلى الخلية.[46] وأثناء النقل المعتمد على الأنيبيب إلى النواة تُنسخ سلسلة الرنا الفيروسية عكسيا إلى سلسلة دنا مزدوجة ثم تُدمج في أحد صبغيات الخلية المضيفة.
يمكن لفيروس الإيدز إصابة الخلايا المتغصنة بواسطة الطريق CD4-CCR5، لكن يمكن أن يُستخدم طريق آخر عبر استخدام مستقبلات لكتين نوع سي (CLRs) الخاصة بالمانوز مثل DC-SIGN.[49] الخلايا المتغصنة هي الخلايا الأولى التي يصادفها الفيروس خلال الانتقال الجنسي، ويُعتقد حاليا أنها تلعب دورا مهما عبر نقل الفيروس إلى الخلايا التائية حين يُلتقط الفيروس في الغشاء المخاطي بواسطتها[49]. يُعتقد أن تواجد FEZ1 -الذي يظهر طبيعيا في العصبونات- يقي من إصابة الخلية بفيروس الإيدز.[50]

كان يُعتقد لمدة طويلة أن دخول فيروس الإيدز-1 -وكذلك دخول العديد من الفيروسات الراجعة الأخرى- يحدث حصريا عند الغشاء الخلوي، لكن تم الإبلاغ حديثا كذلك عن إصابة فعالة لفيروس الإيدز-1 للخلايا بواسطة إدخال خلوي متوسَّط بالكلاثرين لا يعتمد على الأس الهيدروجيني واقتُرح مؤخرا أنه يشكل الطريق الوحيد للدخول الفعال.[51][52][53][54][55]
التضاعف والنسخ
بعد فترة قصيرة من دخول القفيصة الفيروسية إلى الخلية، يحرر الإنزيم المسمى الناسخ العكسي جينوم سلسلة الرنا المفردة موجبة الاتجاه من البروتينات المرتبطة بها وينسخها إلى جزيء دنا متمم (cDNA).[56] عملية النسخ العكسي عرضة للأخطاء كثيرا ويمكن أن تسبب الطفرات الناتجة مقاومة الدواء أو تسمح للفيروس بتجنب الجهاز المناعي الخاص بالجسم. للناسخ العكسي نشاط ريبونوكلياز ويقوم بتفكيك الرنا الفيروسي أثناء تخليق الدنا المتمم، وكذلك نشاط بوليميراز الدنا المعتمد على الدنا ويقوم بتخليق سلسلة دنا موجبة الاتجاه من سلسلة الدنا المتممة المضادة للاتجاه.[57] معا، تشكل سلسلة الدنا المتممة والسلسلة المتممة لها سلسلة دنا مزدوجة فيروسية وتُنقل بعدها إلى النواة. دمج الدنا الفيروسي في جينوم الخلية المضيفة يتم بواسطة إنزيم فيروسي آخر يسمى الدامج.[56]
يمكن أن يبقى الدنا الفيروسي المدمج في سبات أثناء مرحلة من مراحل الإصابة بالفيروس تسمى كمون الفيروس.[56] لتخليق الفيروس بفعالية، يجب أن تتوفر عوامل نسخ محددة، وأكثرها أهمية هو NF-κB (العامل النووي كابا B)، الذي يُنظَّم بالزيادة حين تصبح الخلايا التائية منشطة.[58] هذا يعني على الأرجح أن الخلايا التي يستهدفها فيروس الإيدز ويدخل إليها ويقتلها في النهاية هي الخلايا التي تحارب الإصابة بنشاط.
أثناء تضاعف الفيروس تُنسخ طليعة فيروس الدنا إلى جزيئات رنا ويخضع بعضها لعمية وصل الرنا لإنتاج جزيئات رنا رسول بالغة، بعد ذلك تُصدّر جزيئات الرنا الرسول هذه من النواة إلى السيتوبلازم أين تتم ترجمتها إلى البروتينات المنظمة tat (الذي يشجع على عمليات تخليق جديدة للفيروس) وRev الذي يرجع عند تخليقه إلى النواة ويرتبط بنسخ رنا فيروسية غير موصولة كاملة الطول ويسمح لها بمغادرة النواة.[59] تعمل بعض جزيئات الرنا كاملة الطول هذه كنسخ جديدة من جينوم الفيروس، بينما تعمل أخرى كجزيئات رنا رسول تُترجَم لإنتاج البروتينات البنيوية Gag وEnv. ترتبط بروتينات Gag بجينوم الرنا الفيروسي لتجّمعه مع بروتينات أخرى لتشكيل فيروسات جديدة.[60]
يبدو أن فيروس الإيدز-1 وفيروس الإيدز-2 يجمَّعان بطريقة مختلفة عند إنشاء فيروسات جديدة.[61][62] يُجمَّع فيروس الإيدز-1 من أي جزيئة رنا مناسبة.[63] بينما يجمع فيروس الإيدز-2 بشكل انتقائي من جزيئة الرنا الرسول التي استُخدمت لإنشاء بروتين Gag الذي يقوم بتجميعها.[64]
التأشُّب
- مقالة مفصلة: إعادة التركيب الجيني
يُحاط جينومي رنا اثنين بقفيصة عند إنتاج فيروس إيدز-1 جديد (انظر بنية وجينوم فيروس الإيدز).يمكن أن يحدث تأشيب (إعادة تركيب) بين الجينومين عند تحفيز النسخ والتضاعف بواسطة الناسخ العكسي.[65][66] يحدث التأشيب حين تُنسخ جينومات الرنا مفردة السلسلة موجبة الاتجاه عكسيا لتشكيل دنا، حيث يمكن للناسخ العكسي إنتاج جزيئة دنا عبر التبديل بين جزيئتي الرنا اللتين ينسخ منهما عدة مرات، ويسمى هذا النوع من التأشيب بتأشيب اختيار النسخ. يمكن أن تظهر حوادث التأشيب في أي نقطة من الجينوم، ومعدل حدوثها يتراوح من 2 إلى 20 حادثة تأشيب خلال كل دورة تضاعف، ويمكن لهذه الأحداث بسرعة خلط وتغيير المعلومة الجينية التي تنتقل من الجينوم الأبوي إلى الذرية.[66]
يُنتِج التأشيب الفيروسي تغيرا جينيا يساهم على الأرجح في تطور مقاومة الفيروس للعلاج بمضادات الفيروسات القهقرية.[67] يمكن أن يساهم التأشيب كذلك -نظريا- في التغلب على الدفاعات المناعية للكائن المضيف. مع ذلك، لكي تحدث التأقلمات والميزات الإيجابية من هذه التغيرات الجينية، يجب أن يكون الجينومان المجمعان في فيروس جديد معدي قد نشآ من سلالات فيروسية سالفة مختلفةٍ في التركيب الجيني. ومن غير المعروف كم مرة يحدث تجميع خلط مثل ذلك تحت الظروف الطبيعية.[68]
اقترح بونهوفر وزملاؤه أن تبديل القالب (السلسلة) بواسطة الناسخ العكسي يعمل كعملية ترميم للتعامل مع الشروخات والتضررات في جينوم الرنا مفرد السلسلة.[69] واقترح هو وتيمان أن التأشيب هو تأقلمٌ لترميم التضرر الحاصل لجينومات الرنا.[65] تبديل السلسلة القالب (تأشيب اختيار النسخ) بواسطة الناسخ العكسي يمكن أن يولِّد نسخا غير متضررة من الدنا الجينومي من نسختي جينوم رنا مفرد السلسلة متضررتين. يمكن أن تفسر وجهة النظر هذه التي تقول بوجود فائدة تأقلمية من التأشيب لدى فيروس الإيدز لماذا يحتوي كل فيروس على جينومين كاملين بدل واحد فقط. فضلا عن ذلك، وجهة النظر القائلة بأن التأشيب هو عملية ترميم تعني أن فائدة الترميم يمكن أن تحدث في كل دورة وأن هذه الفائدة يمكن تحقيقها سواء كان الجينومان مختلفين في التركيب الجيني أو لا. من وجهة نظر أن التأشيب في فيروس الإيدز هو عملية ترميم، توليد نسخا مؤشّبة سيكون نتيجة -وليس سببا- لتطور تبديل السلسلة القالب.[69]
تسبب الإصابة بفيروس الإيدز-1 التهابا مزمنا وإنتاج أنواع الأكسجين التفاعلية.[70] لهذا يمكن أن يكون جينوم فيروس الإيدز عرضة للإجهاد التأكسدي، بما في ذلك شروخات في سلسلة الرنا المفردة. بالنسبة لفيروس الإيدز والفيروسات بصفة عامة، تعتمد الإصابة والعدوى الناجحة على التغلب على استراتيجيات الدفاع التي تشمل في العادة إنتاج أنواع أكسجين تفاعلية ضارة ومتلفة للجينوم. وعليه اقترح ميتشود وزملاؤه أن التأشيب بواسطة الفيروسات هو تأقلمٌ لإصلاح أضرار الجينوم، وأن النسخ المتأشبة هي نواتج ثانوية يمكن أن توفر فائدة منفصلة.[71]
التجميع والتحرير

الخطوة الأخيرة من دورة الفيروس هي تجميع فيروسات الإيدز-1 الجديدة ويحدث ذلك عند الغشاء الخلوي للخلية المضيفة. يذهب عديد البروتين Env (gp160) بعد تخليقه إلى داخل الشبكة الإندوبلازمية ثم ينقل إلى جهاز غولجي أين يتم قصه بواسطة بروتين الفيورين وينتج عنه البروتينان السكريان الخاصان بغلاف فيروس الإيدز gp41 وgp120.[72] بعد ذلك يُنقل هذان الأخيران إلى الغشاء الخلوي الخاص بالخلية المضيفة ويقوم gp41 بإرساء وتثبيت gp120 على الغشاء. يرتبط عديدي البروتين Gag (p55) وGag-Pol (p160) كذلك بالسطح الداخلي للغشاء الخلوي إلى جانب جينوم الرنا الفيروسي عندما يبدأ الفيروس الجديد في التبرعم من الخلية المضيفة. الفيروس المتبرعم يكون غير ناضج بعد لأن عديدات البروتين gag ماتزال يجب أن تقص لتشكيل البروتينات المكونة للمطرس والقفيصة والقفيصة المنواة. وهذا القص يتم بواسطة بروتياز فيروسي تم تجميعه مع الفيروس ويمكن تثبيطه بواسطة الأدوية المضادة للفيروسات الراجعة من قسم مثبط البروتياز. بعد ذلك تتجمع مختلف المكونات البنيوية لإنتاج فيروس إيدز جديد ناضج.[73] ولا يمكن سوى للفيروسات الناضجة إصابة خلايا أخرى.
الانتشار داخل الجسد

تسمى العملية الكلاسيكية عند إصابة فيروس لخلية ما " بالانتشار الحر إلى الخلايا" لتمييزها عن عملية إصابة جديدة معروفة حديثا تسمى "الانتشار من خلية إلى خلية".[74] في عملية الانتشار الحر إلى الخلايا (انظر الصورة) تتبرعم فيروسات الإيدز من الخلايا التائية المصابة وتدخل إلى الدم أو السائل خارج الخلوي وبعدها تصيب خلية تائية أخرى بعد الالتقاء بها صدفة.[74] يمكن لفيروس الإيدز كذلك أن ينتشر مباشرة من خلية لأخرى بواسطة عملية الانتشار من خلية إلى خلية التي وُصِف لها مساران. في الأول: يمكن لخلية تائية نقل الفيروس مباشرة لخلية تائية أخرى مستهدفة عبر مشبك فيروسي.[48][75] والثاني: يمكن لخلية مقدمة للمستضد (APC) مثل الخلية البلعمية أو المتغصنة أن تنقل الفيروس إلى الخلايا التائية إما عبر عملية إصابة فعالة كما هو الحال لدى البلعميات أو عبر التقاط ونقل الفيروسات بشكل مفروق كما هو الحال لدى الخلايا المتغصنة.[76] أيا كان المسار المستعمل، فقد أُبلغ أن الانتشار من خلية إلى خلية أكثر فعالية من الانتشار الحر إلى الخلايا.[77] تساهم عدة عوامل في هذه الفعالية منها: تبرعم الفيروس المستقطب نحو موقع اتصال الخلية بالخلية، التموضع القريب للخلايا من بعضها وهو ما يخفض من مرحلة انتشار الفيروس عبر السائل، وتكدس مستقبلات دخول فيروس الإيدز على الخلايا المستهدفة في منطقة الاتصال.[75] يُعتقد أن الانتشار من خلية إلى خلية مهم بشكل خاص في الأنسجة اللمفاوية أين تكون الخلايا التائية المساعِدة محزَّمة بكثافة ويكون احتمال تآثرها المتكرر كبيرا.[74] دعمت دراسات التصوير أثناء الحياة
- مقالة مفصلة: الأنواع الفرعية لفيروس العوز المناعي البشري

يختلف فيروس الإيدز عن العديد من الفيروسات في كونه يملك تباينا جينيا كبيرا، هذا التنوع هو نتيجة لدورة تضاعفه السريعة التي يتم فيها إنشاء 1010 فيروس كل يوم، بالإضافة إلى معدل طفرات كبير يقدر بحوالي 3 x 10−5 لكل نوكليوتيد لكل دورة تضاعف وكذلك الخصائص التأشيبية للناسخ العكسي.[80][81][82]
تؤدي هذه السمات المميزة إلى نشوء العديد من مغايرات فيروس الإيدز في مريض واحد في مدة يوم واحد فقط.[80] يزداد هذا التنوع حين تصاب خلية واحدة في نفس الوقت بنوعين (مغايرتين) أو أكثر من فيروس الإيدز. عندما تحدث الإصابة المتزامنة، يمكن أن يتكون جينوم فيروسات الذرية من سلاسل رنا من سلالتين مختلفتين. بعدها يصيب هذا الفيروس الهجين خلية جديدة ويخضع لدورة تضاعف، وحين يتم ذلك يولِّد الناسخ العكسي عبر تبديل النسْخ بين السلسلتين جيئة وذهابا تسلسل دنا راجعٍ يكون مؤشبا (توليفة) بين جينومي الفيروسين الأبوين. يبدو هذا التأشيب جليا حين يحدث بين الأنواع الفرعية.[80]
تطوَّرَ فيروس العوز المناعي السعالي (SIV) ذو الصلة القريبة من فيروس الإيدز البشري إلى العديد من السلالات، وصُنف حسب الأنواع الطبيعية المضيفة له. يُعتقد أن سلالات الفيروس السعالي الخاصة بالقرد الإفريقي الأخضر (SIVagm) والمنجبي الأسخم (SIVsmm) لها تاريخ تطوري كبير مع مضيفاتها. تأقلمت هذه المضيفات مع تواجد الفيروس فيها،[83] وهذا التواجد يكون بمستويات كبيرة لكن لا يُحدث سوي استجابة مناعية خفيفة،[84] لا تسبب تطور متلازمة نقص المناعة المكتسبة للسعالي،[85] ولا يخضع لطفرات وتأشُّب كثيف كما هو الحال لدى فيروس الإيدز البشري (HIV).[86]
في المقابل، حين تصيب هذه السلالات الأنواع التي تأقلمت مع فيروس عوز المناعة السعالي (SIV) مثل المكاك الريسوسي أو مكاك طويل الذيل، تطور هذه الحيوانات مرض الإيدز ويولِّد الفيروس تنوعا جينيا مماثلا لما يُشاهَد لدى إصابة البشر بالفيروس.[87] الفيروس السعالي الخاص بالشامبانزي (SIVcpz) هو أقرب الأقارب الجينية لفيروس الإيدز-1 وهو مرتبط بزيادةٍ في الوفيات وأعراض مشابهة لأعراض الإيدز لدى مضيفه الطبيعي.[88] يبدو أن الفيروس السعالي الشبمانزي انتقل حديثا نسبيا إلى جمهرتي الشامبانزي والبشر، لذلك لم يتأقلم هذان المضيفان مع الفيروس بعد.[83] هذا الفيروس فقد كذلك وظيفة الجين nef المتواجدة لدى معظم فيروسات السعالي. في مغايرات فيروس السعالي غير المسببة للمرض، يثبط nef تنشيط الخلايا التائية عبر واسم عنقود التمايز 3 (CD3). وظيفة nef في سلالات فيروس السعالي غير الممرضة هي تخفيض التعبير السيتوكينات المعززة للاتهاب MHC-1 والإشارات التي تؤثر على تحرك وانتقال الخلايا التائية. في فيروس الإيدز-1 والفيروس السعالي الشمبانزي، لا يثبط nef تنشيط الخلايا التائية وفقد تلك الوظيفة. من دون هذه الوظيفة، على الأرجح قد يؤدي نفاذ الخلايا التائية إلى عوز المناعة.[88][89]
تم تحديد ثلاث مجموعات من فيروس الإيدز-1 بناء على اختلافات في منطقة الغلاف (env) هي M وN وO.[90] المجموعة M هي الأكثر انتشارا وتنقسم بدورها إلى ثمان مجموعات فرعية (أو كلادات) بناء على جينوماتها الكاملة التي تختلف جغرافيا.[91] أكثر الأنواع الفرعية انتشارا هو B (المتواجد أساسا في شمال أمريكا وأوروبا)، A وD (المتواجدان أساسا في إفريقيا)، وC ( المتواجد أساسا في إفريقيا وآسيا). تتفرع هذه الأنواع الفرعية من شجرة تطورية تمثل الخط التطوري للمجموعة M من فيروس الإيدز-1. الإصابة المتزامنة بنوعين مختلفين من الأنواع الفرعية تنتج عنها هيئات مؤشبة منتشرة (CRFs). في عام 2000، وهو آخر عام من دراسة أنجِزت لمعرفة الانتشار العالمي للأنواع الفرعية، كانت نتائج الإصابات كالتالي: 47.2% من النوع الفرعي C، 26.7% من النوع الفرعي A/CRF02_AG، 12.3% من النوع الفرعي B، 5.3% من النوع الفرعي D، 3.2% من النوع الفرعي CRF_AE، والبقية 5.3% كانت مكونة من أنواع فرعية وهيئات مؤشبة منتشرة أخرى.[92] تركز معظم البحوث حول فيروس الإيدز-1 على النوع الفرعي B، ومختبرات قليلة تركز على أنواع فرعية أخرى.[93] افتُرض أن تواجد المجموعة الرابعة "P" مبني على فيروس عُزل سنة 2009.[94] ويبدو أن هذه السلالة مشتقة من فيروس السعالي الخاص بالغوريلا (SIVgor) الذي عُزِل من غوريلا السهول الغربية سنة 2006.[94]
أقرب الأقرباء لفيروس الإيدز-2 هو فيروس السعالي الخاص بالمنجبي الأسخم. بما أن فيروس الإيدز-1 مشتق من فيروس السعالي الشبمانزيّ وفيروس الإيدز-2 من فيروس السعالي الخاص بالمنجبي الأسخم، التركيب الجيني لفيروس الإيدز-2 متماثل جزئيا فقط مع فيروس الإيدز-1 ويماثل بشكل أكبر نظيره لدى فيروس السعالي الخاص بالمنجبي الأسخم.[95]
التشخيص
- مقالة مفصلة: تشخيص فيروس نقص المناعة البشرية
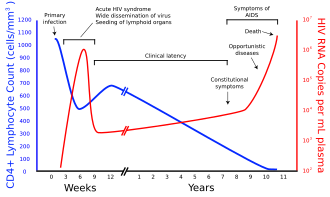
العديد من الأفراد المصابين بفيروس الإيدز لا يعلمون بأنهم مصابون به.[96] على سبيل المثال، أقل من 1% من سكان المدن في إفريقيا النشطين جنسيا تم اختبارهم، وهذه النسبة أقل في المناطق السكانية النائية.[96] زيادة على ذلك، في 2001 حوالي 0.5% فقط من النساء الحوامل اللواتي يرتدن المرافق الصحية الحضرية تم نصحهن، اختبارهن أو حصلن على نتائج فحص.[96] ومجددا هذه النسبة أقل في المرافق الصحية الريفية.[96] بما أن المتبرعين قد لا يعلمون بإصابتهم، فإن الدم المتبرع به ونواتج الدم المستخدمة في الطب والبحوث الطبية تفحص دوريا للكشف عن فيروس الإيدز.[97]
اختبار فيروس الإيدز-1 ينفَّذ في البداية بمقايسة الممتز المناعي المرتبط بالإنزيم (إليزا) (ELISA) للكشف عن الأجسام المضادة للفيروس. والعينات التي لا تُظهر نتيجة تفاعلية من الفحص الأولي بالإليزا تعتبر سلبية لفيروس الإيدز، إلا إذا حدث تعرض جديد لشريك مصاب أو شريك لا تُعلم حالة فيروس الإيدز لديه. العينات التي تكون متفاعلة في فحص إليزا يُعاد اختبارها مجددا مرتين.[98] إن كان أحد الاختبارين متفاعلا، يُبلَّغ عن العينة أنها متكررة التفاعل وتخضع لاختبار تأكيدي بواسطة اختبار تكميلي أكثر دقة (مثل تفاعل البوليميراز المتسلسل (PCR)، لطخة ويسترن، أو بشكل أقل مقايسة التألق المناعي (IFA)). فقط العينات متكررة التفاعل بالإليزا والإيجابية بواسطة مقايسة التألق المناعي أو تفاعل البوليميراز المتسلسل أو لطخة ويسترن هي التي تعتبر إيجابية لفيروس الإيدز وعلامة على الإصابة بالفيروس. قد تُظهر العينات متكررة التفاعل لإليزا أحيانا نتائج فحصٍ بلطخة ويسترن غير محددة، وهو الأمر الذي يحدث بسبب استجابةِ أجسامٍ مضادةٍ غير كاملة لفيروس الإيدز لدى شخص مصاب أو تفاعلات لا نموذجية لدى شخص غير مصاب.[99]
رغم أن مقايسة التألق المناعي تُستخدم لتأكيد الإصابة في هذه الحالات الغامضة، إلا أن هذه المقايسة غير مستخدمة على نطاق واسع. عموما، يجب أن تُجمع عينة أخرى بعد أكثر من شهر ويُعاد اختبار الأفراد الذين نتائج اختبارهم بلطخة ويسترن غير محددة. يمكن استخدام اختبار الحمض النووي رغم توفره بشكل أقل (طريقة مضاعفة الرنا الفيروسي أو طليعة الدنا الفيروسية) للتشخيص في بعض الحالات.[98] زيادة على ذلك، بعض العينات المفحوصة قد لا تمنح نتائج حاسمة بسبب جودة العينة المنخفضة، وفي هذه الحالات تُجمع عينة ثانية وتختبر للكشف عن الإصابة بفيروس الإيدز.
الاختبار الحديث للكشف عن فيروس الإيدز دقيق للغاية عندما تؤخذ فترة النافذة في الحسبان. يكون اختبار تقصٍ واحد صحيحا أكثر من 99% في كل مرة.[100] يقدَّر احتمال حدوث نتيجة إيجابية خاطئة
- مقالة مفصلة: بحوث فيروس الإيدز/الإيدز
تشمل أبحاث فيروس الإيدز/الإيدز جميع البحوث الطبية التي تحاول منع، علاج أو شفاء فيروس الإيدز/إيدز، وكذلك البحوث الأساسية حول طبيعة الفيروس كعامل معدٍ والإيدز كمرض يُسبَّب بواسطة فيروس العوز المناعي البشري.
تشارك العديد من الحكومات ومؤسسات البحث في البحوث حول فيروس الإيدز/الإيدز. وتشمل هذه البحوث التدخلات الصحية السلوكية مثل البحوث في التربية الجنسية وكذلك تطوير العقاقير مثل بحوث مبيدات المكروبات المنتقلة جنسيا، لقاحات فيروس نقص المناعة البشرية والأدوية المضادة للفيروسات الراجعة.[104] تشمل بعض مجالات البحوث الطبية الأخرى مواضيع مثل وقاية قبل التعرض، العلاج الوقائي بعد التعرض للفيروس، الختان وفيروس المناعة البشري وتأثيرات الشيخوخة المتسارعة.
العلاج والانتقال
تاريخ
- مقالة مفصلة: تاريخ مرض الإيدز
الاكتشاف
لوحظ مرض الإيدز سريريا لأول مرة عام 1981 في الولايات المتحدة.[105] كانت الحالات الأولى لمجموعة من متعاطي المخدرات عبر الحقن ورجل مثلي ذو مناعة عليلة من دون سبب معروف أظهر أعراض ذات الرئة التي تسببها المتكيسة، وهي عدوى انتهازية نادرة معروفٌ بأنها تصيب الأفراد الذين جهاز المناعة لديهم ضعيف وعليل.[106] بعد ذلك بوقت قصير طور رجل مثلي آخر سرطان جلد نادر يسمى ساركوما كابوزي.[107][108] ثم تلى ذلك ظهور العديد من الإصابات بذات الرئة والسرطان، وهو ما نبه مراكز السيطرة على الأمراض والوقاية منها الأمريكية فتم تشكيل فريق عمل لمراقبة التفشي.[109] أقدم وصف لحالة مرض بالإيدز كانت في النرويج في بداية عام 1966.[110]
في البداية لم يكن لدى مراكز السيطرة على الأمراض والوقاية منها اسم رسمي للمرض، وغالبا ما كان يشار إليه بالمرض الذي يُحدثه، على سبيل المثال: تضخم العقد اللمفية، وهو المرض الذي سمى به مكتشفو فيروس الإيدز الفيروس أول مرة.[111][112] واستخدموا كذلك ساركوما كابوزي والعداوى الانتهازية، والأخير هو الاسم الذي شُكِّل فريق العمل لمراقبته سنة 1981. في الصحافة العامة، تمت صياغة المصطلح GRID الذي يعني عوز المناعة المرتبط بالمثلي.[113] صاغت السيديسي -أثناء بحثها عن اسم ومراقبتها للمجتمعات المصابة- مصطلح "مرض الهاءات الأربعة" لأن المرض كان يبدو بأنه يصيب المثليين، متعاطي المخدرات، المصابين بالهيموفيليا والهايتيين (جميع هذه الكلمات تبدأ بحرف H في اللغة الإنجليزية).[114][115] لكن بعد معرفة أن الإيدز ليس محصورا في مجتمع المثليين،[116] أُدرِك أن مصطلح GRID كان غير مناسب وتمت صياغة مصطلح الإيدز في اجتماع في يوليو 1982.[117] وبحلول سبتمبر 1982 بدأت السيديسي باستخدامه.[118]
مقالات ذات صلة
المراجع
- http://www.larousse.fr/encyclopedie/medical/virus_de_limmunod%C3%A9ficience_humaine_VIH/16958 — الباب: virus de l'immunodéficience humaine (VIH)
- International Committee on Taxonomy of Viruses (2002). "61.0.6. Lentivirus". معاهد الصحة الوطنية الأمريكية. مؤرشف من الأصل في 30 يناير 200928 فبراير 2006.
- International Committee on Taxonomy of Viruses (2002). "61. Retroviridae". National Institutes of Health. مؤرشف من الأصل في 2 أبريل 201028 فبراير 2006.
- Levy JA (1993). "HIV pathogenesis and long-term survival". AIDS. 7 (11): 1401–10. doi:10.1097/00002030-199311000-00001. PMID 8280406.
- Smith JA, Daniel R (2006). "Following the path of the virus: the exploitation of host DNA repair mechanisms by retroviruses". ACS Chemical Biology. 1 (4): 217–26. doi:10.1021/cb600131q. PMID 17163676.
- Siliciano, R. F.; Greene, W. C. (2011). "HIV Latency". Cold Spring Harbor Perspectives in Medicine. 1 (1): a007096. doi:10.1101/cshperspect.a007096. PMC . PMID 22229121.
- Gilbert PB, McKeague IW, Eisen G, Mullins C, Guéye-NDiaye A, Mboup S, Kanki PJ (February 28, 2003). "Comparison of HIV-1 and HIV-2 infectivity from a prospective cohort study in Senegal". Statistics in Medicine. 22 (4): 573–593. doi:10.1002/sim.1342. PMID 12590415.
- Reeves JD, Doms RW (2002). "Human Immunodeficiency Virus Type 2" ( كتاب إلكتروني PDF ). Journal of General Virology. 83 (Pt 6): 1253–65. doi:10.1099/0022-1317-83-6-1253. PMID 12029140.
- McGovern SL, Caselli E, Grigorieff N, Shoichet BK (2002). "A common mechanism underlying promiscuous inhibitors from virtual and high-throughput screening". Journal of Medicinal Chemistry. 45 (8): 1712–22. doi:10.1021/jm010533y. hdl:11380/977912. PMID 11931626.
- Compared with overview in: Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C. (2007). Lippincott's Illustrated Reviews: Microbiology. Hagerstown, MD: Lippincott Williams & Wilkins. صفحة 3. .
- Various (2008). HIV Sequence Compendium 2008 Introduction ( كتاب إلكتروني PDF ). مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 24 نوفمبر 201731 مارس 2009.
- Chan DC, Fass D, Berger JM, Kim PS (1997). "Core structure of gp41 from the HIV envelope glycoprotein" ( كتاب إلكتروني PDF ). Cell. 89 (2): 263–73. doi:10.1016/S0092-8674(00)80205-6. PMID 9108481. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 14 يناير 2018.
- Klein, Joshua S.; Bjorkman, Pamela J.; Rall, Glenn F. (27 May 2010). "Few and Far Between: How HIV May Be Evading Antibody Avidity". PLOS Pathogens. 6 (5): e1000908. doi:10.1371/journal.ppat.1000908. PMC . PMID 20523901.
- National Institute of Health (June 17, 1998). "Crystal structure of key HIV protein reveals new prevention, treatment targets" (Press release). مؤرشف من الأصل في 19 فبراير 200614 سبتمبر 2006.
- Behrens, Anna-Janina; Vasiljevic, Snezana; Pritchard, Laura K; Harvey, David J; Andev, Rajinder S; Krumm, Stefanie A; Struwe, Weston B; Cupo, Albert; Kumar, Abhinav; Zitzmann, Nicole; Seabright, Gemma E; Kramer, Holger B; Spencer, Daniel I.R; Royle, Louise; Lee, Jeong Hyun; Klasse, Per J; Burton, Dennis R; Wilson, Ian A; Ward, Andrew B; Sanders, Rogier W; Moore, John P; Doores, Katie J; Crispin, Max (2016). "Composition and Antigenic Effects of Individual Glycan Sites of a Trimeric HIV-1 Envelope Glycoprotein". Cell Reports. 14 (11): 2695–706. doi:10.1016/j.celrep.2016.02.058. PMC . PMID 26972002.
- Pritchard, Laura K; Spencer, Daniel I.R; Royle, Louise; Bonomelli, Camille; Seabright, Gemma E; Behrens, Anna-Janina; Kulp, Daniel W; Menis, Sergey; Krumm, Stefanie A; Dunlop, D. Cameron; Crispin, Daniel J; Bowden, Thomas A; Scanlan, Christopher N; Ward, Andrew B; Schief, William R; Doores, Katie J; Crispin, Max (2015). "Glycan clustering stabilizes the mannose patch of HIV-1 and preserves vulnerability to broadly neutralizing antibodies". Nature Communications. 6: 7479. Bibcode:2015NatCo...6.7479P. doi:10.1038/ncomms8479. PMC . PMID 26105115.
- Pritchard, Laura K; Harvey, David J; Bonomelli, Camille; Crispin, Max; Doores, Katie J (2015). "Cell- and Protein-Directed Glycosylation of Native Cleaved HIV-1 Envelope". Journal of Virology. 89 (17): 8932–44. doi:10.1128/JVI.01190-15. PMC . PMID 26085151.
- Crispin, Max; Doores, Katie J (2015). "Targeting host-derived glycans on enveloped viruses for antibody-based vaccine design". Current Opinion in Virology. 11: 63–9. doi:10.1016/j.coviro.2015.02.002. PMC . PMID 25747313.
- Julien, Jean-Philippe; Cupo, Albert; Sok, Devin; Stanfield, Robyn L.; Lyumkis, Dmitry; Deller, Marc C.; Klasse, Per-Johan; Burton, Dennis R.; Sanders, Rogier W. (2013-12-20). "Crystal structure of a soluble cleaved HIV-1 envelope trimer". Science. 342 (6165): 1477–1483. Bibcode:2013Sci...342.1477J. doi:10.1126/science.1245625. ISSN 1095-9203. PMC . PMID 24179159.
- Lyumkis, Dmitry; Julien, Jean-Philippe; de Val, Natalia; Cupo, Albert; Potter, Clinton S.; Klasse, Per-Johan; Burton, Dennis R.; Sanders, Rogier W.; Moore, John P. (2013-12-20). "Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer". Science. 342 (6165): 1484–1490. Bibcode:2013Sci...342.1484L. doi:10.1126/science.1245627. ISSN 1095-9203. PMC . PMID 24179160.
- Sanders, Rogier W.; Derking, Ronald; Cupo, Albert; Julien, Jean-Philippe; Yasmeen, Anila; de Val, Natalia; Kim, Helen J.; Blattner, Claudia; de la Peña, Alba Torrents (2013-09-01). "A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies". PLOS Pathogens. 9 (9): e1003618. doi:10.1371/journal.ppat.1003618. ISSN 1553-7374. PMC . PMID 24068931.
- Pritchard, Laura K.; Vasiljevic, Snezana; Ozorowski, Gabriel; Seabright, Gemma E.; Cupo, Albert; Ringe, Rajesh; Kim, Helen J.; Sanders, Rogier W.; Doores, Katie J. (2015-06-16). "Structural Constraints Determine the Glycosylation of HIV-1 Envelope Trimers". Cell Reports. 11 (10): 1604–1613. doi:10.1016/j.celrep.2015.05.017. ISSN 2211-1247. PMC . PMID 26051934.
- de Taeye, Steven W.; Ozorowski, Gabriel; Torrents de la Peña, Alba; Guttman, Miklos; Julien, Jean-Philippe; van den Kerkhof, Tom L. G. M.; Burger, Judith A.; Pritchard, Laura K.; Pugach, Pavel (2015-12-17). "Immunogenicity of Stabilized HIV-1 Envelope Trimers with Reduced Exposure of Non-neutralizing Epitopes". Cell. 163 (7): 1702–1715. doi:10.1016/j.cell.2015.11.056. ISSN 1097-4172. PMC . PMID 26687358.
- Ouellet DL, Plante I, Landry P, Barat C, Janelle ME, Flamand L, Tremblay MJ, Provost P (April 2008). "Identification of functional microRNAs released through asymmetrical processing of HIV-1 TAR element". Nucleic Acids Research. 36 (7): 2353–65. doi:10.1093/nar/gkn076. PMC . PMID 18299284.
- Klase Z, Winograd R, Davis J, Carpio L, Hildreth R, Heydarian M, Fu S, McCaffrey T, Meiri E, Ayash-Rashkovsky M, Gilad S, Bentwich Z, Kashanchi F (2009). "HIV-1 TAR miRNA protects against apoptosis by altering cellular gene expression". Retrovirology. 6 (1): 18. doi:10.1186/1742-4690-6-18. PMC . PMID 19220914.
- Vasudevan AA, Smits SH, Höppner A, Häussinger D, Koenig BW, Münk C (Nov 2013). "FZJ-2013-05757" "Structural features of antiviral DNA cytidine deaminases". Biological Chemistry (Submitted manuscript). 394 (11): 1357–70. doi:10.1515/hsz-2013-0165. PMID 23787464. مؤرشف من الأصل في 13 مايو 2020.
- Garcia JV, Miller AD (April 1991). "Serine phosphorylation-independent downregulation of cell-surface CD4 by nef". Nature. 350 (6318): 508–11. Bibcode:1991Natur.350..508G. doi:10.1038/350508a0. PMID 2014052. مؤرشف من الأصل في 14 مايو 2020.
- Schwartz O, Maréchal V, Le Gall S, Lemonnier F, Heard JM (March 1996). "Endocytosis of major histocompatibility complex class I molecules is induced by the HIV-1 Nef protein". Nature Medicine. 2 (3): 338–42. doi:10.1038/nm0396-338. PMID 8612235. مؤرشف من الأصل في 14 مايو 2020.
- Stumptner-Cuvelette P, Morchoisne S, Dugast M, Le Gall S, Raposo G, Schwartz O, Benaroch P (October 2001). "HIV-1 Nef impairs MHC class II antigen presentation and surface expression". Proceedings of the National Academy of Sciences of the United States of America. 98 (21): 12144–9. Bibcode:2001PNAS...9812144S. doi:10.1073/pnas.221256498. PMC . PMID 11593029.
- Arrildt, Kathryn Twigg; Joseph, Sarah Beth; Swanstrom, Ronald (March 2012). "The HIV-1 Env Protein: A Coat of Many Colors". Current HIV/AIDS Reports. 9 (Current HIV/AIDS Reports): 53–63. doi:10.1007/s11904-011-0107-3. PMC . PMID 22237899.
- Berger EA, Doms RW, Fenyö EM, Korber BT, Littman DR, Moore JP, Sattentau QJ, Schuitemaker H, Sodroski J, Weiss RA (1998). "A new classification for HIV-1". Nature. 391 (6664): 240. Bibcode:1998Natur.391..240B. doi:10.1038/34571. PMID 9440686. مؤرشف من الأصل في 14 مايو 2020.
- Coakley E, Petropoulos CJ, Whitcomb JM (2005). "Assessing ch vbgemokine co-receptor usage in HIV". Current Opinion in Infectious Diseases. 18 (1): 9–15. doi:10.1097/00001432-200502000-00003. PMID 15647694.
- Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M, Di Marzio P, Marmon S, Sutton RE, Hill CM, Davis CB, Peiper SC, Schall TJ, Littman DR, Landau NR (1996). "Identification of a major co-receptor for primary isolates of HIV-1". Nature. 381 (6584): 661–6. Bibcode:1996Natur.381..661D. doi:10.1038/381661a0. PMID 8649511. مؤرشف من الأصل في 14 مايو 2020.
- Feng Y, Broder CC, Kennedy PE, Berger EA (1996). "HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor". Science. 272 (5263): 872–7. Bibcode:1996Sci...272..872F. doi:10.1126/science.272.5263.872. PMID 8629022.
- Knight SC, Macatonia SE, Patterson S (1990). "HIV I infection of dendritic cells". International Review of Immunology. 6 (2–3): 163–75. doi:10.3109/08830189009056627. PMID 2152500.
- Tang J, Kaslow RA (2003). "The impact of host genetics on HIV infection and disease progression in the era of highly active antiretroviral therapy". AIDS. 17 (Suppl 4): S51–S60. doi:10.1097/00002030-200317004-00006. PMID 15080180.
- Zhu T, Mo H, Wang N, Nam DS, Cao Y, Koup RA, Ho DD (1993). "Genotypic and phenotypic characterization of HIV-1 patients with primary infection". Science. 261 (5125): 1179–81. Bibcode:1993Sci...261.1179Z. doi:10.1126/science.8356453. PMID 8356453.
- van't Wout AB, Kootstra NA, Mulder-Kampinga GA, Albrecht-van Lent N, Scherpbier HJ, Veenstra J, Boer K, Coutinho RA, Miedema F, Schuitemaker H (1994). "Macrophage-tropic variants initiate human immunodeficiency virus type 1 infection after sexual, parenteral, and vertical transmission". Journal of Clinical Investigation. 94 (5): 2060–7. doi:10.1172/JCI117560. PMC . PMID 7962552.
- Zhu T, Wang N, Carr A, Nam DS, Moor-Jankowski R, Cooper DA, Ho DD (1996). "Genetic characterization of human immunodeficiency virus type 1 in blood and genital secretions: evidence for viral compartmentalization and selection during sexual transmission". Journal of Virology. 70 (5): 3098–107. doi:10.1128/JVI.70.5.3098-3107.1996. PMC . PMID 8627789.
- Clevestig P, Maljkovic I, Casper C, Carlenor E, Lindgren S, Navér L, Bohlin AB, Fenyö EM, Leitner T, Ehrnst A (2005). "The X4 phenotype of HIV type 1 evolves from R5 in two children of mothers, carrying X4, and is not linked to transmission". AIDS Research and Human Retroviruses. 21 (5): 371–8. doi:10.1089/aid.2005.21.371. PMID 15929699.
- Moore JP (1997). "Coreceptors: implications for HIV pathogenesis and therapy". Science. 276 (5309): 51–2. doi:10.1126/science.276.5309.51. PMID 9122710.
- Powell MK, Benková K, Selinger P, Dogoši M, Kinkorová Luňáčková I, Koutníková H, Laštíková J, Roubíčková A, Špůrková Z, Laclová L, Eis V, Šach J, Heneberg P (2016). "Opportunistic Infections in HIV-Infected Patients Differ Strongly in Frequencies and Spectra between Patients with Low CD4+ Cell Counts Examined Postmortem and Compensated Patients Examined Antemortem Irrespective of the HAART Era". PLOS ONE. 11 (9): e0162704. Bibcode:2016PLoSO..1162704P. doi:10.1371/journal.pone.0162704. PMC . PMID 27611681.
- Karlsson A, Parsmyr K, Aperia K, Sandström E, Fenyö EM, Albert J (1994). "MT-2 cell tropism of human immunodeficiency virus type 1 isolates as a marker for response to treatment and development of drug resistance". The Journal of Infectious Diseases. 170 (6): 1367–75. doi:10.1093/infdis/170.6.1367. PMID 7995974.
- Koot M, van 't Wout AB, Kootstra NA, de Goede RE, Tersmette M, Schuitemaker H (1996). "Relation between changes in cellular load, evolution of viral phenotype, and the clonal composition of virus populations in the course of human immunodeficiency virus type 1 infection". The Journal of Infectious Diseases. 173 (2): 349–54. doi:. PMID 8568295.
- Cheney K, McKnight A (2010). "HIV-2 Tropism and Disease". Lentiviruses and Macrophages: Molecular and Cellular Interactions. Caister Academic Press. .
- Chan DC, Kim PS (1998). "HIV entry and its inhibition". Cell. 93 (5): 681–4. doi:10.1016/S0092-8674(00)81430-0. PMID 9630213.
- Wyatt R, Sodroski J (1998). "The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens". Science. 280 (5371): 1884–8. Bibcode:1998Sci...280.1884W. doi:10.1126/science.280.5371.1884. PMID 9632381.
- Arthos J, Cicala C, Martinelli E, Macleod K, Van Ryk D, Wei D, Xiao Z, Veenstra TD, Conrad TP, Lempicki RA, McLaughlin S, Pascuccio M, Gopaul R, McNally J, Cruz CC, Censoplano N, Chung E, Reitano KN, Kottilil S, Goode DJ, Fauci AS (2008). "HIV-1 envelope protein binds to and signals through integrin alpha(4)beta(7), the gut mucosal homing receptor for peripheral T cells". Nature Immunology. 9 (3): 301–9. doi:10.1038/ni1566. PMID 18264102. مؤرشف من الأصل في 14 مايو 2020.
- Pope M, Haase AT (2003). "Transmission, acute HIV-1 infection and the quest for strategies to prevent infection". Nature Medicine. 9 (7): 847–52. doi:10.1038/nm0703-847. PMID 12835704. مؤرشف من الأصل في 14 مايو 2020.
- Haedicke J, Brown C, Naghavi MH (Aug 2009). "The brain-specific factor FEZ1 is a determinant of neuronal susceptibility to HIV-1 infection". Proceedings of the National Academy of Sciences. 106 (33): 14040–14045. Bibcode:2009PNAS..10614040H. doi:10.1073/pnas.0900502106. PMC . PMID 19667186.
- Daecke J, Fackler OT, Dittmar MT, Kräusslich HG (2005). "Involvement of clathrin-mediated endocytosis in human immunodeficiency virus type 1 entry". Journal of Virology. 79 (3): 1581–1594. doi:10.1128/jvi.79.3.1581-1594.2005. PMC . PMID 15650184.
- Miyauchi K, Kim Y, Latinovic O, Morozov V, Melikyan GB (2009). "HIV Enters Cells via Endocytosis and Dynamin-Dependent Fusion with Endosomes". Cell. 137 (3): 433–444. doi:10.1016/j.cell.2009.02.046. PMC . PMID 19410541.
- Koch P, Lampe M, Godinez WJ, Müller B, Rohr K, Kräusslich HG, Lehmann MJ (2009). "Visualizing fusion of pseudotyped HIV-1 particles in real time by live cell microscopy". Retrovirology. 6: 84. doi:10.1186/1742-4690-6-84. PMC . PMID 19765276.
- Thorley JA, McKeating JA, Rappoport JZ (2010). "Mechanis ms of viral entry: sneaking in the front door". Protoplasma. 244 (1–4): 15–24. doi:10.1007/s00709-010-0152-6. PMC . PMID 20446005.
- Permanyer M, Ballana E, Esté JA (2010). "Endocytosis of HIV: anything goes". Trends in Microbiology. 18 (12): 543–551. doi:10.1016/j.tim.2010.09.003. PMID 20965729.
- Zheng YH, Lovsin N, Peterlin BM (2005). "Newly identified host factors modulate HIV replication". Immunology Letters. 97 (2): 225–34. doi:10.1016/j.imlet.2004.11.026. PMID 15752562.
- "IV. Viruses> F. Animal Virus Life Cycles > 3. The Life Cycle of HIV". Doc Kaiser's Microbiology Home Page. Community College of Baltimore County. يناير 2008. مؤرشف من الأصل في يوليو 26, 2010.
- Hiscott J, Kwon H, Génin P (2001). "Hostile takeovers: viral appropriation of the NF-kB pathway". Journal of Clinical Investigation. 107 (2): 143–151. doi:10.1172/JCI11918. PMC . PMID 11160127.
- Pollard VW, Malim MH (1998). "The HIV-1 Rev protein". Annual Review of Microbiology. 52: 491–532. doi:10.1146/annurev.micro.52.1.491. PMID 9891806.
- Butsch, M.; Boris-Lawrie, K. (2002). "Destiny of Unspliced Retroviral RNA: Ribosome and/or Virion?". Journal of Virology. 76 (7): 3089–94. doi:10.1128/JVI.76.7.3089-3094.2002. PMC . PMID 11884533.
- Hellmund, Chris; Lever, Andrew M. L. (2016-07-14). "Coordination of Genomic RNA Packaging with Viral Assembly in HIV-1". Viruses. 8 (7): 192. doi:10.3390/v8070192. ISSN 1999-4915. PMC . PMID 27428992.
- Soto-Rifo, R.; Limousin, T.; Rubilar, P. S.; Ricci, E. P.; Décimo, D.; Moncorgé, O.; Trabaud, M. A.; André, P.; Cimarelli, A.; Ohlmann, T. (2011). "Different effects of the TAR structure on HIV-1 and HIV-2 genomic RNA translation". Nucleic Acids Research. 40 (6): 2653–2667. doi:10.1093/nar/gkr1093. PMC . PMID 22121214.
- Saad, Jamil S.; Muriaux, Delphine M. (2015-07-28). Role of Lipids in Virus Assembly (باللغة الإنجليزية). Frontiers Media SA. . مؤرشف من الأصل في 14 مايو 2020.
- Ricci, E. P.; Herbreteau, C. H.; Decimo, D.; Schaupp, A.; Datta, S. A. K.; Rein, A.; Darlix, J. -L.; Ohlmann, T. (2008). "In vitro expression of the HIV-2 genomic RNA is controlled by three distinct internal ribosome entry segments that are regulated by the HIV protease and the Gag polyprotein". RNA. 14 (7): 1443–55. doi:10.1261/rna.813608. PMC . PMID 18495939.
- Hu WS, Temin HM (1990). "Retroviral recombination and reverse transcription". Science. 250 (4985): 1227–33. Bibcode:1990Sci...250.1227H. doi:10.1126/science.1700865. PMID 1700865.
- Charpentier C, Nora T, Tenaillon O, Clavel F, Hance AJ (2006). "Extensive recombination among human immunodeficiency virus type 1 quasispecies makes an important contribution to viral diversity in individual patients". Journal of Virology. 80 (5): 2472–82. doi:10.1128/JVI.80.5.2472-2482.2006. PMC . PMID 16474154.
- Nora T, Charpentier C, Tenaillon O, Hoede C, Clavel F, Hance AJ (2007). "Contribution of recombination to the evolution of human immunodeficiency viruses expressing resistance to antiretroviral treatment". Journal of Virology. 81 (14): 7620–8. doi:10.1128/JVI.00083-07. PMC . PMID 17494080.
- Chen J, Powell D, Hu WS (2006). "High frequency of genetic recombination is a common feature of primate lentivirus replication". Journal of Virology. 80 (19): 9651–8. doi:10.1128/JVI.00936-06. PMC . PMID 16973569.
- Bonhoeffer S, Chappey C, Parkin NT, Whitcomb JM, Petropoulos CJ (2004). "Evidence for positive epistasis in HIV-1". Science. 306 (5701): 1547–50. Bibcode:2004Sci...306.1547B. doi:10.1126/science.1101786. PMID 15567861.
- Israël N, Gougerot-Pocidalo MA (1997). "Oxidative stress in human immunodeficiency virus infection". Cellular and Molecular Life Sciences. 53 (11–12): 864–70. doi:10.1007/s000180050106. PMID 9447238. مؤرشف من الأصل في 14 مايو 2020.
- Michod RE, Bernstein H, Nedelcu AM (May 2008). "Adaptive value of sex in microbial pathogens" ( كتاب إلكتروني PDF ). Infection, Genetics and Evolution. 8 (3): 267–85. doi:10.1016/j.meegid.2008.01.002. PMID 18295550. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 11 مايو 2020.
- Hallenberger S, Bosch V, Angliker H, Shaw E, Klenk HD, Garten W (November 26, 1992). "Inhibition of furin-mediated cleavage activation of HIV-1 glycoprotein gp160". Nature. 360 (6402): 358–61. Bibcode:1992Natur.360..358H. doi:10.1038/360358a0. PMID 1360148. مؤرشف من الأصل في 14 مايو 2020.
- Gelderblom HR (1997). "Fine structure of HIV and SIV" ( كتاب إلكتروني PDF ). In Los Alamos National Laboratory (المحرر). HIV sequence compendium. مختبر لوس ألاموس الوطني. صفحات 31–44.
- Zhang C, Zhou S, Groppelli E, Pellegrino P, Williams I, Borrow P, Chain BM, Jolly C (2015). "Hybrid Spreading Mechanisms and T Cell Activation Shape the Dynamics of HIV-1 Infection". PLOS Computational Biology. 11 (4): e1004179. arXiv:. Bibcode:2015PLSCB..11E4179Z. doi:10.1371/journal.pcbi.1004179. PMC . PMID 25837979.
- Jolly C, Kashefi K, Hollinshead M, Sattentau QJ (2004). "HIV-1 cell to cell transfer across an Env-induced, actin-dependent synapse". Journal of Experimental Medicine. 199 (2): 283–293. doi:10.1084/jem.20030648. PMC . PMID 14734528.
- Sattentau Q (2008). "Avoiding the void: cell-to-cell spread of human viruses". Nature Reviews Microbiology. 6 (11): 815–826. doi:10.1038/nrmicro1972. PMID 18923409. مؤرشف من الأصل في 14 مايو 2020.
- Duncan CJ, Russell RA, Sattentau QJ (2013). "High multiplicity HIV-1 cell-to-cell transmission from macrophages to CD4+ T cells limits antiretroviral efficacy". AIDS. 27 (14): 2201–2206. doi:10.1097/QAD.0b013e3283632ec4. PMC . PMID 24005480.
- Sewald X, Gonzalez DG, Haberman AM, Mothes W (2012). "In vivo imaging of virological synapses". Nature Communications. 3: 1320. Bibcode:2012NatCo...3.1320S. doi:10.1038/ncomms2338. PMC . PMID 23271654.
- Sigal A, Kim JT, Balazs AB, Dekel E, Mayo A, Milo R, Baltimore D (2011). "Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy". Nature. 477 (7362): 95–98. Bibcode:2011Natur.477...95S. doi:10.1038/nature10347. PMID 21849975. مؤرشف من الأصل في 14 مايو 2020.
- Robertson DL, Hahn BH, Sharp PM (1995). "Recombination in AIDS viruses". Journal of Molecular Evolution. 40 (3): 249–59. Bibcode:1995JMolE..40..249R. doi:10.1007/BF00163230. PMID 7723052. مؤرشف من الأصل في 14 مايو 2020.
- Rambaut A, Posada D, Crandall KA, Holmes EC (January 2004). "The causes and consequences of HIV evolution". Nature Reviews Genetics. 5 (52–61): 52–61. doi:10.1038/nrg1246. PMID 14708016. مؤرشف من الأصل في 9 نوفمبر 2019.
- Perelson AS, Ribeiro RM (October 2008). "Estimating drug efficacy and viral dynamic parameters: HIV and HCV". Statistics in Medicine. 27 (23): 4647–57. doi:10.1002/sim.3116. PMID 17960579. مؤرشف من الأصل في 13 مايو 2020.
- Sodora DL, Allan JS, Apetrei C, Brenchley JM, Douek DC, Else JG, Estes JD, Hahn BH, Hirsch VM, Kaur A, Kirchhoff F, Muller-Trutwin M, Pandrea I, Schmitz JE, Silvestri G (2009). "Toward an AIDS vaccine: lessons from natural simian immunodeficiency virus infections of African nonhuman primate hosts". Nature Medicine. 15 (8): 861–865. doi:10.1038/nm.2013. PMC . PMID 19661993.
- Holzammer S, Holznagel E, Kaul A, Kurth R, Norley S (2001). "High virus loads in naturally and experimentally SIVagm-infected African green monkeys". Virology. 283 (2): 324–31. doi:10.1006/viro.2001.0870. PMID 11336557.
- Kurth, R.; Norley, S. (1996). "Why don't the natural hosts of SIV develop simian AIDS?". The Journal of NIH Research. 8: 33–37.
- Baier M, Dittmar MT, Cichutek K, Kurth R (1991). "Development of vivo of genetic variability of simian immunodeficiency virus". Proceedings of the National Academy of Sciences of the United States of America. 88 (18): 8126–30. Bibcode:1991PNAS...88.8126B. doi:10.1073/pnas.88.18.8126. PMC . PMID 1896460.
- Daniel MD, King NW, Letvin NL, Hunt RD, Sehgal PK, Desrosiers RC (1984). "A new type D retrovirus isolated from macaques with an immunodeficiency syndrome". Science. 223 (4636): 602–5. Bibcode:1984Sci...223..602D. doi:10.1126/science.6695172. PMID 6695172.
- Keele BF, Jones JH, Terio KA, Estes JD, Rudicell RS, Wilson ML, Li Y, Learn GH, Beasley TM, Schumacher-Stankey J, Wroblewski E, Mosser A, Raphael J, Kamenya S, Lonsdorf EV, Travis DA, Mlengeya T, Kinsel MJ, Else JG, Silvestri G, Goodall J, Sharp PM, Shaw GM, Pusey AE, Hahn BH (2009). "Increased mortality and AIDS-like immunopathology in wild chimpanzees infected with SIVcpz". Nature. 460 (7254): 515–519. Bibcode:2009Natur.460..515K. doi:10.1038/nature08200. PMC . PMID 19626114.
- Schindler M, Münch J, Kutsch O, Li H, Santiago ML, Bibollet-Ruche F, Müller-Trutwin MC, Novembre FJ, Peeters M, Courgnaud V, Bailes E, Roques P, Sodora DL, Silvestri G, Sharp PM, Hahn BH, Kirchhoff F (2006). "Nef-mediated suppression of T cell activation was lost in a lentiviral lineage that gave rise to HIV-1". Cell. 125 (6): 1055–67. doi:10.1016/j.cell.2006.04.033. PMID 16777597.
- Thomson MM, Pérez-Alvarez L, Nájera R (2002). "Molecular epidemiology of HIV-1 genetic forms and its significance for vaccine development and therapy". The Lancet Infectious Diseases. 2 (8): 461–471. doi:10.1016/S1473-3099(02)00343-2. PMID 12150845.
- Carr JK, Foley BT, Leitner T, Salminen M, Korber B, McCutchan F (1998). "Reference sequences representing the principal genetic diversity of HIV-1 in the pandemic" ( كتاب إلكتروني PDF ). In Los Alamos National Laboratory (المحرر). HIV sequence compendium. لوس ألاموس: مختبر لوس ألاموس الوطني. صفحات 10–19.
- Osmanov S, Pattou C, Walker N, Schwardländer B, Esparza J (2002). "Estimated global distribution and regional spread of HIV-1 genetic subtypes in the year 2000". Journal of Acquired Immune Deficiency Syndromes. 29 (2): 184–190. doi:10.1097/00042560-200202010-00013. PMID 11832690.
- Perrin L, Kaiser L, Yerly S (2003). "Travel and the spread of HIV-1 genetic variants". The Lancet Infectious Diseases. 3 (1): 22–27. doi:10.1016/S1473-3099(03)00484-5. PMID 12505029.
- Plantier JC, Leoz M, Dickerson JE, De Oliveira F, Cordonnier F, Lemée V, Damond F, Robertson DL, Simon F (August 2009). "A new human immunodeficiency virus derived from gorillas". Nature Medicine. 15 (8): 871–2. doi:10.1038/nm.2016. PMID 19648927. مؤرشف من الأصل في 14 مايو 2020. ضع ملخصا.
- Keele BF, Van Heuverswyn F, Li Y, Bailes E, Takehisa J, Santiago ML, Bibollet-Ruche F, Chen Y, Wain LV, Liegeois F, Loul S, Ngole EM, Bienvenue Y, Delaporte E, Brookfield JF, Sharp PM, Shaw GM, Peeters M, Hahn BH (Jul 28, 2006). "Chimpanzee reservoirs of pandemic and nonpandemic HIV-1". Science. 313 (5786): 523–6. Bibcode:2006Sci...313..523K. doi:10.1126/science.1126531. PMC . PMID 16728595.
- Kumaranayake, L.; Watts, C. (2001). "Resource allocation and priority setting of HIV/AIDS interventions: addressing the generalized epidemic in sub-Saharan Africa". Journal of International Development. 13 (4): 451–466. doi:10.1002/jid.797.
- Kleinman S (سبتمبر 2004). "Patient information: Blood donation and transfusion". Uptodate. مؤرشف من الأصل في أبريل 12, 2008.
- Centers for Disease Control and Prevention (2001). "Revised guidelines for HIV counseling, testing, and referral". MMWR Recommendations and Reports. 50 (RR–19): 1–57. PMID 11718472.
- Celum CL, Coombs RW, Lafferty W, Inui TS, Louie PH, Gates CA, McCreedy BJ, Egan R, Grove T, Alexander S (1991). "Indeterminate human immunodeficiency virus type 1 western blots: seroconversion risk, specificity of supplemental tests, and an algorithm for evaluation". The Journal of Infectious Diseases. 164 (4): 656–664. doi:10.1093/infdis/164.4.656. PMID 1894929.
- Chou, Roger; Selph, Shelley; Dana, Tracy; Bougatsos, Christina; Zakher, Bernadette; Blazina, Ian; Korthuis, P. Todd (2012-11-20). "Screening for HIV: systematic review to update the 2005 U.S. Preventive Services Task Force recommendation". Annals of Internal Medicine. 157 (10): 706–718. doi:10.7326/0003-4819-157-10-201211200-00007. ISSN 1539-3704. PMID 23165662.
- Chou R, Huffman LH, Fu R, Smits AK, Korthuis PT (July 2005). "Screening for HIV: a review of the evidence for the U.S. Preventive Services Task Force". Annals of Internal Medicine. 143 (1): 55–73. doi:. PMID 15998755.
- Tolle MA, Schwarzwald HL (July 15, 2010). "Postexposure prophylaxis against human immunodeficiency virus". American Family Physician. 82 (2): 161–6. PMID 20642270.
- "Quick Reference Guide—Laboratory Testing for the Diagnosis of HIV Infection: Updated Recommendations" ( كتاب إلكتروني PDF ). cdc.gov. New York State Department of Health. June 27, 2014. صفحات 1–2. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في March 2, 201713 أبريل 2017.
- "HIV Treatment: FDA-Approved HIV Medicines". AIDSinfo. مؤرشف من الأصل في 23 فبراير 2017.
- Mandell, Gerald L.; Bennett, John E.; Dolin, Raphael, المحررون (2010). "Chapter 169". Mandell, Douglas, and Bennett's principles and practice of infectious diseases (الطبعة 7th). Philadelphia: Churchill Livingstone/Elsevier. .
- Gottlieb MS (2006). "Pneumocystis pneumonia—Los Angeles. 1981". American Journal of Public Health. 96 (6): 980–1, discussion 982–3. doi:10.2105/AJPH.96.6.980. PMC . PMID 16714472. مؤرشف من الأصل في 22 أبريل 2009.
- Friedman-Kien AE (October 1981). "Disseminated Kaposi's sarcoma syndrome in young homosexual men". Journal of the American Academy of Dermatology. 5 (4): 468–71. doi:10.1016/S0190-9622(81)80010-2. PMID 7287964.
- Hymes KB, Cheung T, Greene JB, Prose NS, Marcus A, Ballard H, William DC, Laubenstein LJ (September 1981). "Kaposi's sarcoma in homosexual men — a report of eight cases". The Lancet. 2 (8247): 598–600. doi:10.1016/S0140-6736(81)92740-9. PMID 6116083. مؤرشف من الأصل في 14 مايو 2020.
- Basavapathruni A, Anderson KS (December 2007). "Reverse transcription of the HIV-1 pandemic". The FASEB Journal. 21 (14): 3795–3808. doi:10.1096/fj.07-8697rev. PMID 17639073.
- Lederberg, Joshua, المحرر (2000). Encyclopedia of Microbiology (الطبعة 2nd). Burlington: Elsevier. صفحة 106. . مؤرشف من الأصل في 13 مايو 202009 يونيو 2016.
- Centers for Disease Control (1982). "Persistent, generalized lymphadenopathy among homosexual males". Morbidity and Mortality Weekly Report. 31 (19): 249–251. PMID 6808340. مؤرشف من الأصل في 1 يوليو 2019.
- Barré-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vézinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L (1983). "Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS)". ساينس. 220 (4599): 868–871. Bibcode:1983Sci...220..868B. doi:10.1126/science.6189183. PMID 6189183.
- Altman LK (May 11, 1982). "New homosexual disorder worries health officials". نيويورك تايمز. مؤرشف من الأصل في 13 مايو 202031 أغسطس 2011.
- Gilman, Sander L (1987). Gilman, Sander L. (المحرر). "AIDS and Syphilis: The Iconography of Disease". October. 43: 87–107. doi:10.2307/3397566. JSTOR 3397566.
- "Making Headway Under Hellacious Circumstances" ( كتاب إلكتروني PDF ). الجمعية الأمريكية لتقدم العلوم. July 28, 2006. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 23 سبتمبر 200923 يونيو 2008.
- Centers for Disease Control (1982). "Opportunistic infections and Kaposi's sarcoma among Haitians in the United States". Morbidity and Mortality Weekly Report. 31 (26): 353–354, 360–361. PMID 6811853. مؤرشف من الأصل في 30 مارس 2020.
- Kher U (July 27, 1982). "A Name for the Plague". Time. مؤرشف من الأصل في March 7, 200810 مارس 2008.
- Centers for Disease Control (1982). "Update on acquired immune deficiency syndrome (AIDS)—United States". Morbidity and Mortality Weekly Report. 31 (37): 507–508, 513–514. PMID 6815471.