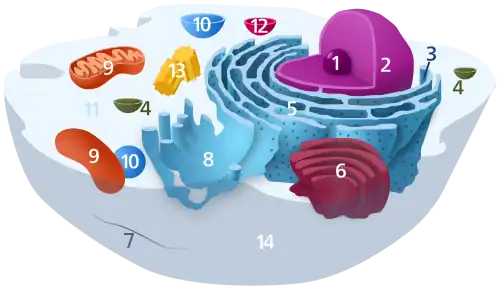
(1) Nucléole
(2) Noyau
(3) Ribosomes
(4) Vésicule
(5) Réticulum endoplasmique rugueux (ou granuleux) (REG)
(6) Appareil de Golgi
(7) Cytosquelette
(8) Réticulum endoplasmique lisse
(9) Mitochondries
(10) Peroxysome
(11) Cytosol
(12) Lysosome
(13) Centrosome (constitué de deux centrioles)
(14) Membrane plasmique
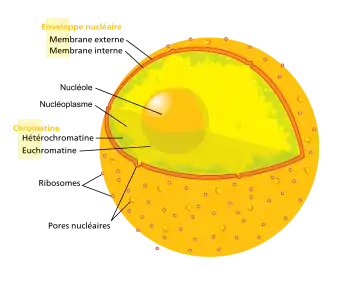
En biologie cellulaire, le nucléole est le plus gros sous-compartiment du noyau des cellules eucaryotes. Aucune membrane ne délimite cet espace particulier du noyau, qui n'est de ce fait jamais considéré comme un organite. Le nombre de nucléoles peut varier au cours du cycle cellulaire (généralement 1 seul en phase G0).
Le nucléole est composé de protéines, d'ADN et d'ARN et se forme autour de régions chromosomiques particulières appelées NOR (Nucleolar Organizing Regions, ou Régions Organisatrices de Nucléoles)
C'est en particulier le lieu où se produit la transcription des ARN ribosomiques[1] (ARNr 5.8S, 18S et 28S), la maturation de ces ARN ribosomiques à partir de précurseurs et la première partie de l'assemblage des deux sous-unités des ribosomes avec l'association de certaines protéines ribosomiques. La fin de l'assemblage des ribosomes se produit dans le cytoplasme sur un ARNm après export des pré-ribosomes au travers du pore nucléaire.
Histoire
Le nucléole a été identifié par microscopie à fond clair au cours des années 1830[2],[3]. On connaissait peu de choses sur la fonction du nucléole jusqu'en 1964 avec une étude[4] des nucléoles par John Gurdon et Donald Brown chez la grenouille africaine Xenopus laevis qui suscita un intérêt croissant pour la fonction et l'étude de la structure détaillée du nucléole. Ils ont constaté que 25 % des œufs de grenouille n'avaient pas de nucléole et que de tels œufs n'étaient pas capables de vie. La moitié des œufs avaient un nucléole et 25 % en avaient deux. Ils ont conclu que le nucléole avait une fonction nécessaire à la vie. En 1966, Max L. Birnstiel et ses collaborateurs ont montré via des expériences d'hybridation d'acide nucléique que l'ADN présent dans le nucléole code l'ARN ribosomique[5],[6].
Observation du nucléole
Il est le plus souvent basophile et donc visible en microscopie optique via des colorants basiques (Pyronine en rouge et bleu de Giemsa), toutefois c'est la microscopie électronique qui a véritablement permis l'observation de ses composants.
En microscopie électronique, on remarque le nucléole dans le noyau par sa forte densité d'électrons (il forme un point sombre dans le noyau). Ceci est dû à la présence de nombreux ARN et au fait que le nucléole soit associé à de l'hétérochromatine (ADN fortement condensé). Il contient les différentes unités de transcription pour les ARN ribosomiques qui y sont transcrits (environ 200 en tout dans le génome humain). On observe de 4 à 7 nucléoles juste après la mitose, puis ils fusionnent entre eux, se présentant généralement sous la forme de 2 nucléoles en début de phase G1 et de 1 nucléole en fin de phase G1 et durant toute la phase G0.
- la zone fibrillaire, centrale, où on retrouve l'ADN possédant les gènes codant l'ARN précurseur 45S ;
- la zone granulaire, périphérique, où sont assemblées les sous-unités des ribosomes (40S et 60S). Celles-ci sont ensuite individuellement exportées dans le cytoplasme ;
- la zone chromosomique, où on retrouve la chromatine condensée adjacente à la chromatine diffuse ;
- la zone amorphe qui est en fait une matrice constituée de protéines et d'ARN.
Le biologiste Oscar Miller fut le premier à isoler par centrifugation le nucléole du noyau de plusieurs cellules, puis décompacter les composants fibrillaires denses qu'il contient. En les observant au microscope électronique en transmission, on a ainsi pu découvrir des structures que Miller [7],[8] a poétiquement appelées "arbres de Noël" en raison de leur forme. Le "tronc" de ces arbres est une molécule d'ADN (ce que l'on peut mettre en évidence avec un test à la DNase), les "branches" sont des ARNr, les "boules" des protéines diverses et les grains raccordant les branches au tronc sont des ARN polymérases.
Dans un noyau, le nucléole est d'autant plus grand que la biosynthèse des ribosomes est importante dans la cellule qui le contient.
Rôle dans la synthèse des ribosomes
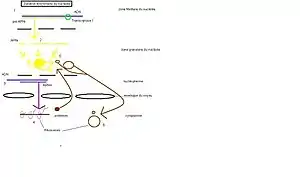
La formation des ribosomes dans le nucléole s'effectue en plusieurs étapes qui se déroulent dans les zones fibrillaires et granulaires.
- Étape 1 : L'ADN codant les ARNr est transcrit en un pré-ARNr 45 S, cette étape se passe dans la zone fibrillaire du nucléole.
- Étape 2 : Le pré-ARNr est ensuite découpé en trois ARNr 5,8 S ; 18 S et 28 S
- Étape 3 : Simultanément dans le nucléoplasme, de l'ADN codant les protéines constitutives des ribosomes (ribonucléoprotéines) est transcrit en ARN messager.
- Étape 4 : L'ARN messager est traduit en protéines dans le cytoplasme.
- Étape 5 : Les protéines traduites entrent dans le noyau et dans le nucléole, elles s'associent avec les ARN ribosomiques pour former des pré-ribosomes.
- Étape 6 : Les pré-ribosomes sont exportés dans le cytoplasme, dans lequel ils terminent leur maturation, avant de s'associer et de former un ribosome mature.
Le nucléole évolue pendant le cycle cellulaire. En effet lors de la mitose le nucléole disparaît.
De plus, le nucléole étant le centre de la synthèse des ribosomes, qui sont des éléments indispensables à la synthèse des protéines, son activité et donc sa taille vont varier en fonction de l'intensité de la synthèse des protéines dans la cellule.
Composition
Les composants biochimiques du nucléole sont :
- Des ARN (≈ 10 %) :
- Majoritairement des ARNr qui sont transcrits dans la zone fibrillaire et maturés (protéines de maturation + ARNsno) dans la zone granulaire. Les pré-ARNr ne sont pas épissés, ils sont clivés par plusieurs ribonucléases.
- De l'ADN (≈ 5 %)
- 5 paires de chromosomes acrocentriques (13, 14, 15, 21, 22) possédant les régions NOR (Nucleolar Organizing Regions) codant l’ARNr 45S.
- Des protéines (≈ 85 %) :
- l'ARN polymérase I, qui transcrit les loci d'ADN ribosomique en un précurseur de 45S (pré-ARNr) (35 S chez Saccharomyces cerevisiae).
- Ainsi que des enzymes diverses et des protéines de liaison dont certaines sont argyrophiles (colorables par l'argent), appelées protéines AgNOR.
Notes et références
- ↑ D. Hernandez-Verdun et E. Louvet, « Le nucléole : structure, fonctions et maladies associées », Med. Sci., vol. 20, p. 37-44, 2004.
- ↑ (en) Thoru Pederson, « The Nucleolus », Cold Spring Harbor Perspectives in Biology, vol. 3, no 3, , a000638 (ISSN 1943-0264, PMID 21106648, DOI 10.1101/cshperspect.a000638, lire en ligne, consulté le )
- ↑ Thoru Pederson, « The Nucleolus », Cold Spring Harbor Perspectives in Biology, vol. 3, no 3, (ISSN 1943-0264, PMID 21106648, PMCID PMC3039934, DOI 10.1101/cshperspect.a000638, lire en ligne, consulté le )
- ↑ Donald D. Brown et J. B. Gurdon, « ABSENCE OF RIBOSOMAL RNA SYNTHESIS IN THE ANUCLEOLATE MUTANT OF XENOPUS LAEVIS », Proceedings of the National Academy of Sciences of the United States of America, vol. 51, no 1, , p. 139–146 (ISSN 0027-8424, PMID 14106673, lire en ligne, consulté le )
- ↑ M. L. Birnstiel, H. Wallace, J. L. Sirlin et M. Fischberg, « Localization of the ribosomal DNA complements in the nucleolar organizer region of Xenopus laevis », National Cancer Institute Monograph, vol. 23, , p. 431–447 (ISSN 0083-1921, PMID 5963987, lire en ligne, consulté le )
- ↑ H. Wallace et M. L. Birnstiel, « Ribosomal cistrons and the nucleolar organizer », Biochimica et Biophysica Acta (BBA) - Nucleic Acids and Protein Synthesis, vol. 114, no 2, , p. 296–310 (DOI 10.1016/0005-2787(66)90311-X, lire en ligne, consulté le )
- ↑ Oscar Miller et B. Beatty, « Visualization of nucleolar genes », Science, vol. 194,p. 955-957, 1969.
- ↑ Steven McKnight, «Oscar Miller (1925–2012) Retrospective», Science, vol. 335, 1457, 2012