تطور الثدي ، المعروف أيضاً باسم mammogenesis، هو عبارة عن عملية معقدة بيولوجية تحدث خلال حياة الأنثى. حيث تمر عبر عدة مراحل، بما في ذلك تطور ما قبل الولادة، وسن البلوغ، والحمل. عند سن اليأس، يتوقف نمو الثدي ويحدث ضمور للثديين. هذه العملية تؤدي إلي ظهور عضو بارز على الصدر يعرف باسم الثدي في الرئيسيات، والذي له دور في إظهار بعض السمات الجنسية كما أنه يعمل كغدة لإفراز اللبن والهرمونات. يدخل في هذه العملية مجموعة متنوعة من الهرمونات، وعوامل النمو، وأهمها: هرمون الاستروجين، والبروجسترون، والبرولاكتين، وهرمون النمو.
| الثدي | |
|---|---|
| الاسم اللاتيني mamma[1] |
|
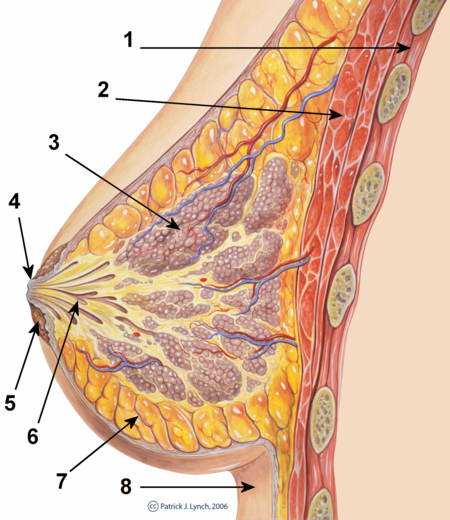
 شكل الثدي البشري
| |
| تفاصيل | |
[ ] |
|
الكيمياء الحيوية
الهرمونات
العوامل الرئيسية لتطور الثدي هي من الهرمونات الستيرويدية؛ الإستروجين والبروجسترون، وهرمون النمو (GH)، عامل النمو شبيه الانسولين-1 (IGF-1)، والبرولاكتين.[2] وتحفز هذه الهرمونات نشاط عوامل النمو، مثل EGF، IGF-1 ، EGF ، IGF-1 ، و عوامل نمو الأرومة الليفية (FGF)، والتي بدورها لها أدوار محددة في نمو الثدي ونضجه.[2]
عند سن البلوغ، يبدأ إفراز الهرمون المحفز للغدد التناسلية (GnRH) بطريقة نابضة، من تحت المهاد.[3][4] هذا الهرمون، بدوره، يحفز إفراز الهرمونات الجنسية؛ الهرمون المنبه للجريب (FSH)، والهرمون الملوتن (LH)، من الغدة النخامية.[3][4] تنتقل هذه الهرمونات إلى المبيضين خلال مجرى الدم وتسبب إفراز هرمون الاستروجين والبروجستيرون، ويتم إطلاقهم في الجسم بكميات متغيرة مع كل دورة شهرية.[3][4] هرمون النمو (GH)، الذي يفرز من الغدة النخامية، وعامل النمو شبيه الانسولين-1 (IGF-1)، والذي يتم إنتاجه في الجسم استجابةً للـGH ، هي هرمونات تساعد علي النمو.[5] خلال مرحلة ما قبل الولادة، والرضاعة، والطفولة، تكون مستويات GH و IGF-1 منخفضة، ولكنها تزداد تدريجيًا وتصل إلى ذروتها عند البلوغ،[6] حيث تحدث زيادة 1.5 إلى 3 أضعاف في إفراز هرمون النمو النابض و 3 أضعاف أو أكثر في مستويات IGF-1 في هذا الوقت.[7] في أواخر مرحلة المراهقة ومراحل البلوغ المبكر، تنخفض مستويات GH و IGF-1 بشكل ملحوظ،[8] وتستمر في الانخفاض طوال بقية الحياة.[6] وقد وجد أن كلاً من هرمون الاستروجين وهرمون النمو (GH) ضروريان لنمو الثدي عند البلوغ- ففي غياب أي منهما، لن يحدث أي تطور.[9][10] علاوة على ذلك، وجد أن معظم دور هرمون النمو في نمو الثدي يكون من خلال تحفيز إفراز IGF-1، والذي بدوره يقوم بإنقاذ نمو الثدي في غياب هرمون النمو.[10] يحدث تحفيز IGF-1 وإفرازه من قبل همون النمو في جميع أنواع الأنسجة في الجسم تقريبًا، ولكن بشكل خاص في الكبد، الذي يمثل مصدر 80٪ تقريبًا من IGF-1 الموجود بالجسم.[6][11][12] على الرغم من أن IGF-1 مسؤول عن معظم دور هرمون النمو في التوسّع في نمو الثدي، فقد وجد أن هرمون النمو نفسه يلعب دورًا مباشرًا إضافيًا أيضًا، لأنه يزيد من تعبير مستقبلات هرمون الاستروجين (ER) في الثدي، في حين أن IGF-1، على النقيض من ذلك، وجد أنه لا يفعل ذلك.[9][10][13][14][15]
تطور الثديين خلال مرحلة ما قبل الولادة تكون مستقل وغير معتمدة علي الجنس البيولوجي والهرمونات الجنسية.[16] خلال مراحل التطور الجنيني، يتم توليد براعم الثدي، التي تتكون فيها شبكات من الأنابيب، التي تتكون من الأديم الظاهر.[17] ستصبح هذه الأنابيب البدائية في النهاية عند النضج هي القنوات اللثوية أو القنوات اللبنية، التي تربط نسيج الثدي (حِزَم "اللبن")، بعناقيد تشبه العنب في (الحويصلات الثديية)، وتحمل اللبن إلى الحلمتين.[18] حتى سن البلوغ، تبقى شبكات الأنابيب في براعم الثدي بدائية وخاملة،[2] حيث لا تظهر أي فروق بين ثدي الذكر والأنثي.[16] خلال فترة البلوغ في الإناث، فإن الإستروجين، بالتزامن مع GH / IGF-1، ومن خلال تفعيل مستقبلات هرمون الاستروجين ألفا (ERα) على وجه التحديد،[19][20] يسبب نمو وتحويل الأنابيب الصغيرة إلى نظام الأقنية الناضجة للثديين.[16][17][21] تحت تأثير الإستروجين، تنبت القنوات وتستطيل، ويتكون لها براعم نهاية طرفية (TEBs)، تخترق طبقة الدهون وتتفرع.[16][17][21] يستمر هذا حتى تتشكل شبكة تشبه الشجرة من القنوات المتفرعة، والتي تنغمس وتملأ طبقة الدهون بأكملها في الثدي.[2][16][17][21] بالإضافة إلى دوره في التوسط في تطوير القنوات، يتسبب الاستروجين في نمو النسيج اللحظي و الأنسجة الدهنية لتتراكم،[16][17] وتزيد في الحجم.[22]
يؤثر البروجسترون، بالتزامن مع GH / IGF-1 بالمثل كالاستروجين، على نمو الثديين خلال فترة البلوغ وبعد ذلك أيضًا.[16][17][21] إلى حد أقل من الإستروجين، يساهم البروجسترون في تطوير القنوات في هذا الوقت، وكما يتضح من نتائج التجارب علي الفئران التي عولجت بمضادات مستقبلات البروجسترون (ميفيبريستون)، حيث حدث تأخر (وإن كانت طبيعية في نهاية المطاف، بسبب عمل الاستروجين من تلقاء نفسه) في نمو القنوات خلال سن البلوغ، وتم التأكدأن البروجسترون يحث على نمو القنوات من تلقاء نفسهه في الغدة الثديية بشكل رئيسي عن طريق حث وإفراز (amphiregulin)، وهو عامل النمو نفسه الذي يدفع الأستروجين في المقام الأول إلى التوسط في عمله على تطوير القنوات.[16][16][17][21][23][24] ومع ذلك، فإن الحويصلات البدائية فقط تتطور استجابة لمستويات ما قبل الحمل من البروجسترون والإستروجين، وتستمر في التطور حتى يحدث الحمل، إذا حدث ذلك.[17] بالإضافة إلى GH / IGF-1، فإن هرمون الاستروجين ضروري للبروجسترون للتأثير على الثديين،[16][21] حيث أن الإستروجين يعمل علي الثديين عن طريق حث التعبير علي مستقبل البروجسترون (PR) في النسيج الظهاري للثدي.[24][24]
خلال الحمل، يحدث نمو ونضوج الثدي بشكل واضح ونضوج من أجل الاستعداد للرضاعة.[16][25][26] يزداد مستويات هرمون الاستروجين والبروجيسترون بشكل كبير، [16] حتي يصل في أواخر الحمل إلي مستويات تكون أعلى مئات الأضعاف من مستويات الدورة الشهرية المعتادة.[27] يسبب الاستروجين والبروجستيرون إفراز مستويات عالية من البرولاكتين من الغدة النخامية الأمامية،[28][29] والتي تصل إلى مستويات أعلى 20 مرة من مستويات الدورة الشهرية العادية.[27] كما يرتفع مستوى IGF-1 و IGF-2 بشكل كبير أثناء الحمل، بسبب إفراز هرمون النمو (PGH) من المشيمة).[30] أثناء الحمل يحدث مزيد من تطور ونمو القنوات اللبنية، عن طريق الاستروجين، مرة أخرى بالتزامن مع GH / IGF-1.[17][18] بالإضافة إلى ذلك، فإن الأستروجين، والبروجسترون (مرة أخرى على وجه التحديد من خلالPRB) ،[24] والبرولاكتين، بالاقتران مع GH / IGF-1، وكذلك عامل النمو الشبيه بالإنسولين 2 (IGF- 2)،[31][32] يعملون معاً، في استكمال تطوير ونمو القنوات والحويصلات في الغدد الثديية أثناء الحمل.[17][18][33][34][35] بالإضافة إلى دوره البرولاكتين في نمو القنوات والحويصلات، لفإنه أيضاً يعمل على زيادة حجم الحلمة أثناء الحمل.[36] وبحلول نهاية الشهر الرابع من الحمل، يكون الثديان على أتم الاستعداد للرضاعة.[26][37][38]
أيضاً يلعب كل من الأنسولين، والجلوكوكورتيكويد مثل الكورتيزول (ومعها الهرمون الموجه لقشر الكظر (ACTH))، و هرمونات الغدة الدرقية (و معها الهرمون المنبه للدرقية (TSH) و الهرمون المطلق لموجهة الدرقية (TRH))،أدوارًا مبسّطة ولكن غير مفهومة بشكل جيد / ضعيفة في نمو الثدي أثناء فترة البلوغ والحمل، ولكنها مطلوبة للتطوير الوظيفي الكامل.[39][40][41][42] كما وجد أن اللبتين هو عامل هام في نمو الغدة الثديية، حيث وجد أنه يعمل علي تعزيز انتشار الخلايا الظهارية الثديية.[3][43]
على النقيض من الهرمونات الجنسية المرتبطة بالإناث؛ الإستروجين والبروجسترون، وجد أن الهرمونات الجنسية المرتبطة بالذكور؛ الأندروجينات، مثل التستوستيرون و ديهدروتستوستيرون (DHT)، تعمل علي معاكسة عمل الاستروجين في الثديين.[33][42][44][45] حيث أنها تعمل من خلال الحد من التعبير عن مستقبلات هرمون الاستروجين في أنسجة الثدي.[44][45][46] في غياب النشاط الذكوري، كما هو الحال في النساء ذوات متلازمة عدم حساسية الأندروجين الكاملة [CAIS]، فإن المستويات المتواضعة من الإستروجين (50 بيكوغرام / مل) قادرة على التوسُّع في نمو الثدي بشكل كبير، حيث يظهر هؤلاء المرضي بأحجام كبيرة للثدي.[33] المزيج من المستويات العليا من الأندروجينات (حوالي 10 أضعاف أكثر) والمستويات الأقل بكثير من الاستروجين (حوالي 10 أضعاف أقل)،[47] بسبب المبيضين في الإناث التي تنتج كميات كبيرة من هرمون الاستروجين ولكن كميات منخفضة من الأندروجينات و الخصية في الذكور التي تنتج كميات كبيرة من الأندروجينات ولكن كميات منخفضة من هرمون الاستروجين،[48] هو السبب الذي يجعل الذكور بشكل عام ليس لها ثديًا بارزًا أو متطورًا نسبيًا بالنسبة للإناث.[42][49]
كالسيتريول، الشكل الهرموني الفعال لـ فيتامين د، الذي يعمل من خلال مستقبلات فيتامين د (VDR)، وجد أنه منظم سلبي لتطور الغدة الثديية في الفئران، خلال سن البلوغ.[35][50][51] بالإضافة إلى ذلك، تم إثبات أن عملية إزالة مستقلات فيتامين د قد أدت إلى زيادة استجابة أنسجة الغدة الثديية لدى الفئران إلى الإستروجين والبروجيستيرون، والذي كان يمثل زيادة في نمو الخلايا استجابة لهذه الهرمونات.[50][52][53]
يمكن الإشارة إلى آلية محتملة للآثار التنظيمية السلبية لـ مستقبلات فيتامين د على نمو الثدي عن طريق دراسة مكملات فيتامين D3 في النساء، حيث وجد أن فيتامين D3 يمنع ظهور (cyclooxygenase 2 (COX-2 في الثدي، وبالتالي، يقلل ويزيد، على التوالي، مستويات البروستاغلاندين E2 (PGE2) و(TGF-β2)، وهو عامل تثبيط معروف في نمو الثدي.[54][55][56] تبعاً لذلك، فإن إفراز COX-2 في نسيج الغدة الثديية ينتج تضخم الغدة اللبنية بالإضافة إلى نمو الغدة الثديية المبكرة في الفئران الإناث، مما يعكس النمط الظاهري للفئران الذين تم إزالة مستقبلات فيتامين د لديهم، ويظهر تأثير تنشيطي قوي لـ COX-2، والذي يتم تقليله بواسطة تنشيط مستقبلات فيتامين د في الغدد الثديية.[55][56] أيضا وفقا لذلك، وجد أن نشاط COX-2 في الثديين يرتبط ارتباطًا إيجابيًا بحجم الثدي لدى النساء.[57]
عوامل النمو
ينتج الإستروجين والبروجسترون والبرولاكتين وكذلك GH / IGF-1 آثارهما على نمو الثدي عن طريق تعديل الإشارات الناتجة في نسيج الثدي من مجموعة متنوعة من عوامل النمو،[21][40][58][59][60] بما في ذلك IGF-1، IGF-2،[61] EGF، FGF، HGF،[62] TNF-α، TNF-β، TGF-α،[63] TGF-β،[34][64][65] RANKL،[34] وLIF.[34] هذه العوامل تنظم النمو الخلوي، وتكاثرها، وتمايزها عبر تفعيل بعض الإشارات بين الخلايا والتي تتحكم في وظيفتها، مثل Erk، Akt، JNK، Jak .[11][66][67][68]
استنادًا إلى الأبحاث التي أجريت على مستقبل عامل نمو البشرة (EGFR) في الفئران، فإن EGFR، وهو الهدف الجزيئي للـEGF، و TGF-α، [2] وجد أنها تكون ضرورية لنمو وتطور الغدد الثديية.[69] الاستروجين والبروجسترون يتوسطان تطوير القنوات بشكل رئيسي من خلال تحريض تعبير مادة كيميائية تسمي (amphiregulin)، وبالتالي تنشيط EGFR.[23][61][66][70][70][71] وفقًا لذلك، فإن علاج الفئران بالـamphiregulin أو بروابط EGFR أخرى مثل TGF-α أو heregulin يحفز تطوير القنوات في الغدة الثديية للفئران، هذه النتائج تحدث حتى في حالة عدم وجود هرمون الاستروجين والبروجسترون.[65][72] لأن كلاً من IGF-1R و EGFR ضروريان بشكل مستقل لتطور الغدة اللبنية، كما وجد أن الجمع بين IGF-1 و EGF، من خلال مستقبلات كل منهما، يحفز نمو الخلايا البشرية الظهارية الثديية، وبالتالي فإن عوامل النمو يبدو أنها تعمل معًا في التوسط في نمو الثدي.[73][74][75]
تم العثور على مستويات مرتفعة من HGF، وإلى حد أقل، IGF-1 (بنسبة 5.4 أضعاف و 1.8 ضعف، على التوالي)، في الأنسجة اللحمية الثديية، في حالات كبر الثدي، وهي حالة نادرة جدًا جدًا.[76][77]
الرضاعة
عند الولادة، ينخفض الاستروجين والبروجيسترون بسرعة إلى مستويات منخفضة للغاية، حيث أن مستويات البروجسترون تكون غير واضحة.[16] على العكس، تبقى مستويات البرولاكتين مرتفعة.[16][25] بينما يعمل الاستروجين والبروجيسترون على تثبيط عملية تصنيع وإفراز اللبن نتيجة لإفراز البرولاكتين عن طريق كبت التعبير عن مستقبلات البرولاكتين (PRLR) في نسيج الثدي، فإن غيابها المفاجئ ي}دي إلى بدء إنتاج الحليب والرضاعة عن طريق البرولاكتين.[16][25] قد يزيد التعبير عن PRLR في نسيج الثدي بمقدار 20 ضعفاً عندما تنخفض مستويات هرمون الاستروجين والبروجسترون عند الولادة.[16] مع بداية الرضاعة من الرضيع، فإن البرولاكتين والأوكسيتوسين يتم إفرازهما ويستكملان عملية إنتاج الحليب وإفرازه.[16][17][25] يثبط البرولاكتين إفراز LH و FSH، مما يؤدي بدوره إلى استمرار انخفاض مستويات هرمون الاستروجين والبروجسترون، ويحدث مؤقتًا انقطاع الطمث (غياب الدورة الشهرية).[25] في غياب الإرضاع المنتظم، والذي يحافظ على تركيزات البرولاكتين عالية، تنخفض مستويات البرولاكتين بسرعة، وتستأنف الدورة الشهرية، وبالتالي ستعود مستويات الاستروجين والبروجسترون الطبيعية.[25]
حجم الثدي ومخاطر الاصابة بالسرطان
بعض العوامل في مورفولوجيا الثدي، بما في ذلك كثافته، تكون متورطة بوضوح في سرطان الثدي. في حين أن حجم الثدي قابل للتوريث بشكل كبير، فإن العلاقة بين حجم الثدي والسرطان غير مؤكدة. لم يتم تحديد المتغيرات الجينية التي تؤثر على حجم الثدي بعد.[78][79][80][81]
من خلال دراسات ارتباطية على مستوى الجينوم، تم ربط مجموعة متنوعة من الجينات المسئولة عن تعدد الأشكال البيولوجية بحجم الثدي.[78] بعض هذه الجينات تشمل rs7816345 بالقرب من ZNF703 (zinc finger protein 703)؛ rs4849887 و rs17625845 flanking INHBB (inhibin βB؛ rs12173570 بالقرب من ESR1 (ERα) ؛ rs7089814 في ZNF365 ((zinc finger protein 365) ؛ rs12371778 بالقرب من PTHLH (هرمون شبيه بالهرمونات الدرقية) ؛ rs62314947 بالقرب من AREG (amphiregulin)؛ [78] وكذلك rs10086016 عند 8p11.23 و rs5995871 في متلازمة الحذف 22q13 (يحتوي على جين MKL1، الذي وجد أنه يعدل النشاط النسخي لل ERα).[82] ويرتبط العديد من هذه الأشكال مع خطر الإصابة بسرطان الثدي، مما يكشف عن ارتباط إيجابي محتمل بين حجم الثدي وخطر الإصابة بسرطان الثدي.[78][82] ومع ذلك، على العكس، تظهر بعض الأشكال المتعددة ارتباطًا سلبيًا بين حجم الثدي وخطر الإصابة بسرطان الثدي.[82] في أي حال، خلص التحليل التلوي إلى أن حجم الثدي وخطر الإصابة بسرطان الثدي لهما صلة مهمة في الواقع.[83]
ترتبط مستويات IGF-1 بشكل إيجابي مع حجم الثدي لدى النساء.[84] بالإضافة إلى ذلك، يرتبط غياب التكرار الشائع 19 في جين IGF1 ارتباطًا إيجابيًا بكثافة الثدي عند النساء، وكذلك مع مستويات IGF-1 العالية أثناء استخدام وسائل منع الحمل عن طريق الفم مع التقليل من الانخفاض الطبيعي المرتبط بالسن في توزع IGF-1 في هؤلاء النساء.[84] هناك تباين كبير في انتشار الأليل IGF1-19 المتكرر بين المجموعات العرقية، وقد وجد أن غيابه يكون أعلى بين النساء الأفريقيات الأمريكيات.[84][85]
ارتبط تعبير COX-2 بشكل إيجابي مع حجم الثدي و الالتهاب في نسيج الثدي، وكذلك مع خطر سرطان الثدي وتشخيصه.[33][57][86][87][88]
- مرحلة تانر لنمو الثدي

طالع أيضاً
مراجع
- "mammal – Definitions from Dictionary.com". Dictionary.reference.com. مؤرشف من الأصل في 08 مارس 201631 أكتوبر 2011.
- Hynes NE, Watson CJ (2010). "Mammary Gland Growth Factors: Roles In Normal Development And In Cancer". Cold Spring Harb Perspect Biol. 2 (8): a003186. doi:10.1101/cshperspect.a003186. PMC . PMID 20554705. [1]
- Ismail Jatoi; Manfred Kaufmann (11 February 2010). Management of Breast Diseases. Springer Science & Business Media. صفحات 12, 27. . مؤرشف من الأصل في 14 أبريل 2020.
- Ronnie Ann Rosenthal; Michael E. Zenilman; Mark R. Katlic (29 June 2013). Principles and Practice of Geriatric Surgery. Springer Science & Business Media. صفحات 325–. . مؤرشف من الأصل في 08 يوليو 2016.
- Shane Bullock; Majella Hayes (20 September 2012). Principles of Pathophysiology. Pearson Higher Education AU. صفحات 349–. . مؤرشف من الأصل في 29 نوفمبر 2019.
- Chong YM, Subramanian A, Sharma AK, Mokbel K (2007). "The potential clinical applications of insulin-like growth factor-1 ligand in human breast cancer". Anticancer Res. 27 (3B): 1617–24. PMID 17595785.
- Shim KS (2015). "Pubertal growth and epiphyseal fusion". Ann Pediatr Endocrinol Metab. 20 (1): 8–12. doi:10.6065/apem.2015.20.1.8. PMC . PMID 25883921.
- Jaak Jürimäe; Andrew P. Hills; T. Jürimäe (1 January 2010). Cytokines, Growth Mediators, and Physical Activity in Children During Puberty. Karger Medical and Scientific Publishers. صفحات 5–. . مؤرشف من الأصل في 14 أبريل 2020.
- Ruan W, Kleinberg DL (1999). "Insulin-like growth factor I is essential for terminal end bud formation and ductal morphogenesis during mammary development". Endocrinology. 140 (11): 5075–81. doi:10.1210/endo.140.11.7095. PMID 10537134. مؤرشف من الأصل في 10 نوفمبر 2019.
- Kleinberg DL, Feldman M, Ruan W (2000). "IGF-I: an essential factor in terminal end bud formation and ductal morphogenesis". J Mammary Gland Biol Neoplasia. 5 (1): 7–17. PMID 10791764.
- Pauline M. Camacho (26 September 2012). Evidence-Based Endocrinology. Lippincott Williams & Wilkins. صفحات 20, 98. . مؤرشف من الأصل في 14 أبريل 2020.
- Kleinberg DL, Ruan W (2008). "IGF-I, GH, and sex steroid effects in normal mammary gland development". J Mammary Gland Biol Neoplasia. 13 (4): 353–60. doi:10.1007/s10911-008-9103-7. PMID 19034633. مؤرشف من الأصل في 13 مارس 2020.
- Feldman M, Ruan W, Tappin I, Wieczorek R, Kleinberg DL (1999). "The effect of GH on estrogen receptor expression in the rat mammary gland". J. Endocrinol. 163 (3): 515–22. doi:10.1677/joe.0.1630515. PMID 10588825.
- Felice, Dana L.; El-Shennawy, Lamiaa; Zhao, Shuangping; Lantvit, Daniel L.; Shen, Qi; Unterman, Terry G.; Swanson, Steven M.; Frasor, Jonna (2013). "Growth Hormone Potentiates 17β-Estradiol-Dependent Breast Cancer Cell Proliferation Independently of IGF-I Receptor Signaling". Endocrinology. 154 (9): 3219–3227. doi:10.1210/en.2012-2208. ISSN 0013-7227. PMC . PMID 23782942.
- Brisken; Malley (2 December 2010). "Hormone Action in the Mammary Gland". Cold Spring Harbor Perspectives in Biology. Cold Spring Harb Perspect Biol. 2 (12): a003178. doi:10.1101/cshperspect.a003178. PMC . PMID 20739412.
- Leonard R. Johnson (2003). Essential Medical Physiology. Academic Press. صفحة 770. . مؤرشف من الأصل في 29 نوفمبر 2019.
- Anthony W. Norman; Helen L. Henry (30 July 2014). Hormones. Academic Press. صفحة 311. . مؤرشف من الأصل في 14 أبريل 2020.
- Susan Blackburn (14 April 2014). Maternal, Fetal, & Neonatal Physiology. Elsevier Health Sciences. صفحات 146–. . مؤرشف من الأصل في 14 أبريل 2020.
- Jerome Frank Strauss; Robert L. Barbieri (13 September 2013). Yen and Jaffe's Reproductive Endocrinology. Elsevier Health Sciences. صفحات 236–. . مؤرشف من الأصل في 14 أبريل 2020.
- Scaling AL, Prossnitz ER, Hathaway HJ (2014). "GPER mediates estrogen-induced signaling and proliferation in human breast epithelial cells and normal and malignant breast". Horm Cancer. 5 (3): 146–60. doi:10.1007/s12672-014-0174-1. PMC . PMID 24718936.
- Jane Coad; Melvyn Dunstall (2011). Anatomy and Physiology for Midwives, with Pageburst online access,3: Anatomy and Physiology for Midwives. Elsevier Health Sciences. صفحة 413. . مؤرشف من الأصل في 14 أبريل 2020.
- Elmar P. Sakala (2000). Obstetrics and Gynecology. Lippincott Williams & Wilkins. صفحات 376–. . مؤرشف من الأصل في 14 أبريل 2020.
- Aupperlee MD, Leipprandt JR, Bennett JM, Schwartz RC, Haslam SZ (2013). "Amphiregulin mediates progesterone-induced mammary ductal development during puberty". Breast Cancer Res. 15 (3): R44. doi:10.1186/bcr3431. PMC . PMID 23705924.
- Sandra Z. Haslam; Janet R. Osuch (1 January 2006). Hormones and Breast Cancer in Post-Menopausal Women. IOS Press. صفحات 42, 69. . مؤرشف من الأصل في 14 أبريل 2020.
- Susan Scott Ricci; Terri Kyle (2009). Maternity and Pediatric Nursing. Lippincott Williams & Wilkins. صفحات 435–. . مؤرشف من الأصل في 28 نوفمبر 2019.
- James W. Wood. Dynamics of Human Reproduction: Biology, Biometry, Demography. Transaction Publishers. صفحات 333–. . مؤرشف من الأصل في 08 أبريل 2020.
- Horst-Dieter Dellmann (9 March 2013). Comparative Endocrinology of Prolactin. Springer Science & Business Media. صفحات 181–. . مؤرشف من الأصل في 14 أبريل 2020.
- Stefan Silbernagl; Agamemnon Despopoulos (1 January 2011). Color Atlas of Physiology. Thieme. صفحات 305–. . مؤرشف من الأصل في 14 أبريل 2020.
- Barbara Fadem (2007). High-yield Comprehensive USMLE Step 1 Review. Lippincott Williams & Wilkins. صفحات 445–. . مؤرشف من الأصل في 14 أبريل 2020.
- L. Joseph Su; Tung-chin Chiang (14 June 2015). Environmental Epigenetics. Springer London. صفحات 93–. . مؤرشف من الأصل في 14 أبريل 2020.
- Brisken, Cathrin; Ayyannan, Ayyakkannu; Nguyen, Cuc; Heineman, Anna; Reinhardt, Ferenc; Jan, Tian; Dey, S.K.; Dotto, G.Paolo; Weinberg, Robert A. (2002). "IGF-2 Is a Mediator of Prolactin-Induced Morphogenesis in the Breast". Developmental Cell. 3 (6): 877–887. doi:10.1016/S1534-5807(02)00365-9. ISSN 1534-5807.
- Kleinberg DL, Barcellos-Hoff MH (2011). "The pivotal role of insulin-like growth factor I in normal mammary development". Endocrinol. Metab. Clin. North Am. 40 (3): 461–71, vii. doi:10.1016/j.ecl.2011.06.001. PMID 21889714.
- Jerome F. Strauss, III; Robert L. Barbieri (13 September 2013). Yen and Jaffe's Reproductive Endocrinology. Elsevier Health Sciences. صفحات 236–. . مؤرشف من الأصل في 14 أبريل 2020.
- Kirby I. Bland; Edward M. Copeland III (9 September 2009). The Breast: Comprehensive Management of Benign and Malignant Diseases. Elsevier Health Sciences. صفحات 44–45. . مؤرشف من الأصل في 14 أبريل 2020.
- Wanda M. Haschek; Colin G. Rousseaux; Matthew A. Wallig (1 May 2013). Haschek and Rousseaux's Handbook of Toxicologic Pathology. Elsevier Science. صفحات 2675–. . مؤرشف من الأصل في 14 أبريل 2020.
- Karen Wambach; University of Kansas School of Nursing Karen Wambach; Jan Riordan (26 November 2014). Breastfeeding and Human Lactation. Jones & Bartlett Publishers. صفحات 85–. . مؤرشف من الأصل في 14 أبريل 2020.
- Gutzman, Jennifer H; Miller, Kristin K; Schuler, Linda A (2004). "Endogenous human prolactin and not exogenous human prolactin induces estrogen receptor α and prolactin receptor expression and increases estrogen responsiveness in breast cancer cells". The Journal of Steroid Biochemistry and Molecular Biology. 88 (1): 69–77. doi:10.1016/j.jsbmb.2003.10.008. ISSN 0960-0760. PMID 15026085.
- Nelson D. Horseman (6 December 2012). Prolactin. Springer Science & Business Media. صفحات 227–. . مؤرشف من الأصل في 14 أبريل 2020.
- Philip J. Di Saia; William T. Creasman (2012). Clinical Gynecologic Oncology. Elsevier Health Sciences. صفحات 372–. . مؤرشف من الأصل في 14 أبريل 2020.
- Tommaso Falcone; William W. Hurd (2007). Clinical Reproductive Medicine and Surgery. Elsevier Health Sciences. صفحة 253. . مؤرشف من الأصل في 14 أبريل 2020.
- Leon Speroff; Philip D. Darney (November 2010). A Clinical Guide for Contraception. Lippincott Williams & Wilkins. صفحات 21–. . مؤرشف من الأصل في 14 نوفمبر 2016.
- Christopher B. Wilson; Victor Nizet; Yvonne Maldonado; Jack S. Remington; Jerome O. Klein (24 February 2015). Remington and Klein's Infectious Diseases of the Fetus and Newborn Infant. Elsevier Health Sciences. صفحات 190–. . مؤرشف من الأصل في 14 أبريل 2020.
- Mechanisms of Leptin in Mammary Tumorigenesis. ProQuest. 2007. صفحات 3–. . مؤرشف من الأصل في 14 أبريل 2020.
- Jernström H, Olsson H (1997). "Breast size in relation to endogenous hormone levels, body constitution, and oral contraceptive use in healthy nulligravid women aged 19-25 years". Am. J. Epidemiol. 145 (7): 571–80. doi:10.1093/oxfordjournals.aje.a009153. PMID 9098173. مؤرشف من الأصل في 14 أبريل 2020.
- Zhou J, Ng S, Adesanya-Famuiya O, Anderson K, Bondy CA (2000). "Testosterone inhibits estrogen-induced mammary epithelial proliferation and suppresses estrogen receptor expression". FASEB J. 14 (12): 1725–30. doi:10.1096/fj.99-0863com. PMID 10973921. مؤرشف من الأصل في 14 أبريل 2020.
- Eigeliene N, Elo T, Linhala M, Hurme S, Erkkola R, Härkönen P (2012). "Androgens inhibit the stimulatory action of 17β-estradiol on normal human breast tissue in explant cultures". J. Clin. Endocrinol. Metab. 97 (7): E1116–27. doi:10.1210/jc.2011-3228. PMID 22535971.
- Michael Eysenck (17 April 2015). AQA Psychology: AS and A-level Year 1. Psychology Press. صفحات 237–. . مؤرشف من الأصل في 14 أبريل 2020.
- Cecie Starr; Ralph Taggart; Christine Evers (1 January 2012). Biology: The Unity and Diversity of Life. Cengage Learning. صفحات 629–. . مؤرشف من الأصل في 14 أبريل 2020.
- Lemaine V, Cayci C, Simmons PS, Petty P (2013). "Gynecomastia in adolescent males". Semin Plast Surg. 27 (1): 56–61. doi:10.1055/s-0033-1347166. PMC . PMID 24872741.
- Lopes N, Paredes J, Costa JL, Ylstra B, Schmitt F (2012). "Vitamin D and the mammary gland: a review on its role in normal development and breast cancer". Breast Cancer Res. 14 (3): 211. doi:10.1186/bcr3178. PMC . PMID 22676419.
- Welsh J (2007). "Targets of vitamin D receptor signaling in the mammary gland". J. Bone Miner. Res. 22 Suppl 2: V86–90. doi:10.1359/jbmr.07s204. PMID 18290729.
- Narvaez CJ, Zinser G, Welsh J (2001). "Functions of 1alpha,25-dihydroxyvitamin D(3) in mammary gland: from normal development to breast cancer". Steroids. 66 (3–5): 301–8. doi:10.1016/s0039-128x(00)00202-6. PMID 11179738.
- Welsh J (2011). "Vitamin D metabolism in mammary gland and breast cancer". Mol. Cell. Endocrinol. 347 (1–2): 55–60. doi:10.1016/j.mce.2011.05.020. PMID 21669251.
- Qin W, Smith C, Jensen M, Holick MF, Sauter ER (2013). "Vitamin D favorably alters the cancer promoting prostaglandin cascade". Anticancer Res. 33 (9): 3861–6. PMID 24023320.
- Chang SH, Ai Y, Breyer RM, Lane TF, Hla T (2005). "The prostaglandin E2 receptor EP2 is required for cyclooxygenase 2-mediated mammary hyperplasia". Cancer Res. 65 (11): 4496–9. doi:10.1158/0008-5472.CAN-05-0129. PMID 15930264.
- Al-Salihi MA, Ulmer SC, Doan T, Nelson CD, Crotty T, Prescott SM, Stafforini DM, Topham MK (2007). "Cyclooxygenase-2 transactivates the epidermal growth factor receptor through specific E-prostanoid receptors and tumor necrosis factor-alpha converting enzyme". Cell. Signal. 19 (9): 1956–63. doi:10.1016/j.cellsig.2007.05.003. PMC . PMID 17572069.
- Markkula A, Simonsson M, Rosendahl AH, Gaber A, Ingvar C, Rose C, Jernström H (2014). "Impact of COX2 genotype, ER status and body constitution on risk of early events in different treatment groups of breast cancer patients". Int. J. Cancer. 135 (8): 1898–910. doi:10.1002/ijc.28831. PMC . PMID 24599585.
- Hynes, N. E.; Watson, C. J. (2010). "Mammary Gland Growth Factors: Roles in Normal Development and in Cancer". Cold Spring Harbor Perspectives in Biology. 2 (8): a003186–a003186. doi:10.1101/cshperspect.a003186. ISSN 1943-0264. PMC . PMID 20554705.
- Jay R. Harris; Marc E. Lippman; C. Kent Osborne; Monica Morrow (28 March 2012). Diseases of the Breast. Lippincott Williams & Wilkins. صفحات 94–. . مؤرشف من الأصل في 14 أبريل 2020.
- Lamote I, Meyer E, Massart-Leën AM, Burvenich C (2004). "Sex steroids and growth factors in the regulation of mammary gland proliferation, differentiation, and involution". Steroids. 69 (3): 145–59. doi:10.1016/j.steroids.2003.12.008. PMID 15072917.
- LaMarca HL, Rosen JM (2007). "Estrogen regulation of mammary gland development and breast cancer: amphiregulin takes center stage". Breast Cancer Res. 9 (4): 304. doi:10.1186/bcr1740. PMC . PMID 17659070.
- El-Attar HA, Sheta MI (2011). "Hepatocyte growth factor profile with breast cancer". Indian J Pathol Microbiol. 54 (3): 509–13. doi:10.4103/0377-4929.85083. PMID 21934211.
- Bates SE, Valverius EM, Ennis BW, Bronzert DA, Sheridan JP, Stampfer MR, Mendelsohn J, Lippman ME, Dickson RB (1990). "Expression of the transforming growth factor-alpha/epidermal growth factor receptor pathway in normal human breast epithelial cells". Endocrinology. 126 (1): 596–607. doi:10.1210/endo-126-1-596. PMID 2294006.
- Serra R, Crowley MR (2005). "Mouse models of transforming growth factor beta impact in breast development and cancer". Endocr. Relat. Cancer. 12 (4): 749–60. doi:10.1677/erc.1.00936. PMID 16322320.
- Kenney NJ, Bowman A, Korach KS, Barrett JC, Salomon DS (2003). "Effect of exogenous epidermal-like growth factors on mammary gland development and differentiation in the estrogen receptor-alpha knockout (ERKO) mouse". Breast Cancer Res. Treat. 79 (2): 161–73. doi:10.1023/a:1023938510508. PMID 12825851.
- Kariagina A, Xie J, Leipprandt JR, Haslam SZ (2010). "Amphiregulin mediates estrogen, progesterone, and EGFR signaling in the normal rat mammary gland and in hormone-dependent rat mammary cancers". Horm Cancer. 1 (5): 229–44. doi:10.1007/s12672-010-0048-0. PMC . PMID 21258428.
- Hennighausen L, Robinson GW, Wagner KU, Liu X (1997). "Developing a mammary gland is a stat affair". J Mammary Gland Biol Neoplasia. 2 (4): 365–72. PMID 10935024.
- Rawlings JS, Rosler KM, Harrison DA (2004). "The JAK/STAT signaling pathway". J. Cell Sci. 117 (Pt 8): 1281–3. doi:10.1242/jcs.00963. PMID 15020666.
- Sebastian J, Richards RG, Walker MP, Wiesen JF, Werb Z, Derynck R, Hom YK, Cunha GR, DiAugustine RP (1998). "Activation and function of the epidermal growth factor receptor and erbB-2 during mammary gland morphogenesis". Cell Growth Differ. 9 (9): 777–85. PMID 9751121.
- McBryan J, Howlin J, Napoletano S, Martin F (2008). "Amphiregulin: role in mammary gland development and breast cancer". J Mammary Gland Biol Neoplasia. 13 (2): 159–69. doi:10.1007/s10911-008-9075-7. PMID 18398673.
- Sternlicht MD, Sunnarborg SW (2008). "The ADAM17-amphiregulin-EGFR axis in mammary development and cancer". J Mammary Gland Biol Neoplasia. 13 (2): 181–94. doi:10.1007/s10911-008-9084-6. PMC . PMID 18470483.
- Kenney NJ, Smith GH, Rosenberg K, Cutler ML, Dickson RB (1996). "Induction of ductal morphogenesis and lobular hyperplasia by amphiregulin in the mouse mammary gland". Cell Growth Differ. 7 (12): 1769–81. PMID 8959346.
- Strange KS, Wilkinson D, Emerman JT (2002). "Mitogenic properties of insulin-like growth factors I and II, insulin-like growth factor binding protein-3 and epidermal growth factor on human breast epithelial cells in primary culture". Breast Cancer Res. Treat. 75 (3): 203–12. doi:10.1023/a:1019915101457. PMID 12353809.
- Ahmad T, Farnie G, Bundred NJ, Anderson NG (2004). "The mitogenic action of insulin-like growth factor I in normal human mammary epithelial cells requires the epidermal growth factor receptor tyrosine kinase". J. Biol. Chem. 279 (3): 1713–9. doi:10.1074/jbc.M306156200. PMID 14593113.
- Rodland KD, Bollinger N, Ippolito D, Opresko LK, Coffey RJ, Zangar R, Wiley HS (2008). "Multiple mechanisms are responsible for transactivation of the epidermal growth factor receptor in mammary epithelial cells". J. Biol. Chem. 283 (46): 31477–87. doi:10.1074/jbc.M800456200. PMC . PMID 18782770.
- Zhong, Aimei; Wang, Guohua; Yang, Jie; Xu, Qijun; Yuan, Quan; Yang, Yanqing; Xia, Yun; Guo, Ke; Horch, Raymund E.; Sun, Jiaming (2014). "Stromal-epithelial cell interactions and alteration of branching morphogenesis in macromastic mammary glands". Journal of Cellular and Molecular Medicine. 18 (7): 1257–1266. doi:10.1111/jcmm.12275. ISSN 1582-1838. PMC . PMID 24720804.
- Menashe I, Maeder D, Garcia-Closas M, Figueroa JD, Bhattacharjee S, Rotunno M, Kraft P, Hunter DJ, Chanock SJ, Rosenberg PS, Chatterjee N (2010). "Pathway analysis of breast cancer genome-wide association study highlights three pathways and one canonical signaling cascade". Cancer Res. 70 (11): 4453–9. doi:10.1158/0008-5472.CAN-09-4502. PMC . PMID 20460509.
- Eriksson N, Benton GM, Do CB, Kiefer AK, Mountain JL, Hinds DA, Francke U, Tung JY (2012). "Genetic variants associated with breast size also influence breast cancer risk". BMC Med. Genet. 13: 53. doi:10.1186/1471-2350-13-53. PMC . PMID 22747683.
- International position paper on women's health and menopause : a comprehensive approach. DIANE Publishing. صفحات 78–. . مؤرشف من الأصل في 08 أبريل 2020.
- J. Larry Jameson; Leslie J. De Groot (25 February 2015). Endocrinology: Adult and Pediatric. Elsevier Health Sciences. صفحات 238–. . مؤرشف من الأصل في 14 أبريل 2020.
- Quaynor, Samuel D.; Stradtman, Earl W.; Kim, Hyung-Goo; Shen, Yiping; Chorich, Lynn P.; Schreihofer, Derek A.; Layman, Lawrence C. (2013). "Delayed Puberty and Estrogen Resistance in a Woman with Estrogen Receptor α Variant". New England Journal of Medicine. 369 (2): 164–171. doi:10.1056/NEJMoa1303611. ISSN 0028-4793. PMC . PMID 23841731.
- Li J, Foo JN, Schoof N, Varghese JS, Fernandez-Navarro P, Gierach GL, Quek ST, Hartman M, Nord S, Kristensen VN, Pollán M, Figueroa JD, Thompson DJ, Li Y, Khor CC, Humphreys K, Liu J, Czene K, Hall P (2013). "Large-scale genotyping identifies a new locus at 22q13.2 associated with female breast size". J. Med. Genet. 50 (10): 666–73. doi:10.1136/jmedgenet-2013-101708. PMC . PMID 23825393.
- Jansen LA, Backstein RM, Brown MH (2014). "Breast size and breast cancer: a systematic review". J Plast Reconstr Aesthet Surg. 67 (12): 1615–23. doi:10.1016/j.bjps.2014.10.001. PMID 25456291.
- Jernström H, Sandberg T, Bågeman E, Borg A, Olsson H (2005). "Insulin-like growth factor-1 (IGF1) genotype predicts breast volume after pregnancy and hormonal contraception and is associated with circulating IGF-1 levels: implications for risk of early-onset breast cancer in young women from hereditary breast cancer families". Br. J. Cancer. 92 (5): 857–66. doi:10.1038/sj.bjc.6602389. PMC . PMID 15756256.
- Lundin KB, Henningson M, Hietala M, Ingvar C, Rose C, Jernström H (2011). "Androgen receptor genotypes predict response to endocrine treatment in breast cancer patients". Br. J. Cancer. 105 (11): 1676–83. doi:10.1038/bjc.2011.441. PMC . PMID 22033271.
- Martin RM, Lin CJ, Nishi MY, et al. (July 2003). "Familial hyperestrogenism in both sexes: clinical, hormonal, and molecular studies of two siblings". The Journal of Clinical Endocrinology and Metabolism. 88 (7): 3027–34. doi:10.1210/jc.2002-021780. PMID 12843139. مؤرشف من الأصل في 14 أبريل 2020.
- Stratakis CA, Vottero A, Brodie A, et al. (April 1998). "The aromatase excess syndrome is associated with feminization of both sexes and autosomal dominant transmission of aberrant P450 aromatase gene transcription". The Journal of Clinical Endocrinology and Metabolism. 83 (4): 1348–57. doi:10.1210/jc.83.4.1348. PMID 9543166. مؤرشف من الأصل في 14 أبريل 2020.
- Gregory Makowski (22 April 2011). Advances in Clinical Chemistry. Academic Press. صفحة 158. . مؤرشف من الأصل في 14 أبريل 202024 مايو 2012.
مزيد من القراءة
- Hovey, Russell C.; Aimo, Lucila (2010). "Diverse and Active Roles for Adipocytes During Mammary Gland Growth and Function". Journal of Mammary Gland Biology and Neoplasia. 15 (3): 279–290. doi:10.1007/s10911-010-9187-8. ISSN 1083-3021.
- Sun, Susie X.; Bostanci, Zeynep; Kass, Rena B.; Mancino, Anne T.; Rosenbloom, Arlan L.; Klimberg, V. Suzanne; Bland, Kirby I. (2018). "Breast Physiology": 37–56.e6. doi:10.1016/B978-0-323-35955-9.00003-9.