Sitta europaea

La Sittelle torchepot (Sitta europaea) est une espèce de passereaux de la famille des Sittidae. C'est une sittelle de taille moyenne, mesurant 14 cm de longueur. Comme les autres sittelles, c'est un oiseau trapu à la queue courte et au long bec. Les parties supérieures sont bleu-gris, les parties inférieures de couleurs très variables selon la géographie, et peuvent être blanc pur ou roux jaunâtre, avec ou sans zone pâle sur la joue. L'oiseau a également un trait oculaire noir, et présente un certain dimorphisme sexuel, le mâle ayant des couleurs plus prononcées que la femelle. C'est un oiseau bruyant, au répertoire varié, émettant divers types de chants, en vuih, vuih, vuih, vuih… et, en cas d'excitation ou d'alerte, de longues séries de cris sonores en tuit répétés rapidement. Comme les autres membres de son genre, c'est un oiseau très agile qui escalade les arbres avec facilité, descendant parfois le long des troncs d'arbres tête la première.
Ce passereau se nourrit principalement d'insectes, notamment de chenilles et de coléoptères qu'il trouve sur les troncs et les branches des arbres. En automne et en hiver, son régime est complété de fruits secs et de graines. L'espèce visite volontiers les mangeoires à oiseaux. Cette sittelle fait souvent des réserves de nourriture, en cachant, par exemple, des graines dans l'écorce des arbres. La Sittelle torchepot vit en couples territoriaux en période de reproduction. Le couple fait son nid dans un trou d'arbre, souvent un ancien nid de pic, ou parfois dans une cavité naturelle. Si l'entrée est trop large, la femelle en maçonne l'entrée avec de la boue, pratique qui lui vaut son nom de « torchepot ». La ponte compte six à neuf œufs blancs, tachés de brun. La femelle couve seule durant deux semaines ou davantage, étant alors nourrie par le mâle. Après l'éclosion, les deux parents alimentent les jeunes, presque exclusivement d'insectes, et n'élèvent généralement qu'une couvée par an. En dehors de la saison de reproduction, c'est une espèce très sociable qui se joint volontiers à des volées mixtes d'alimentation. Son principal prédateur naturel est l'Épervier d'Europe (Accipiter nisus).
La Sittelle torchepot peuple toute l'Eurasie tempérée et quelques localités des montagnes marocaines. Ses habitats privilégiés sont les forêts mixtes ou de feuillus avec de grands et vieux arbres, notamment des chênes, mais on l'observe aussi communément dans les parcs et les grands jardins. Elle vit jusqu'à 1 500 mètres d'altitude en Europe et Asie tempérées. Si le Congrès ornithologique international reconnaît quinze sous-espèces, on en distingue parfois jusqu'à plus de vingt. Elles peuvent être classées en trois groupes ; les oiseaux de l'Ouest de l'aire de répartition ont les parties inférieures orange chamoisé avec la gorge blanche, ceux du Nord de l'Europe et jusqu'en Russie ont les parties inférieures blanches et ceux encore plus à l'est sont similaires aux ouest-européens, sans la gorge blanche. La Sittelle de Sibérie (S. arctica), est parfois reconnue comme espèce proche mais distincte, ou parfois considérée comme sous-espèce de S. europaea. La Sittelle torchepot jouit d'une aire de répartition extrêmement vaste et ses effectifs importants sont globalement stables ; l'Union internationale pour la conservation de la nature considère donc cet oiseau comme de « préoccupation mineure ».
Description
Mensurations
La Sittelle torchepot est une sittelle de taille moyenne, le mâle adulte de la sous-espèce type S. e. europaea mesurant 14 cm de long pour une envergure de 22,5-27 cm[1], et un poids de 17-28 g.
Plumage
Les parties supérieures sont bleu-gris, la tête porte un trait oculaire noir. Chez la sous-espèce nominale, la gorge et les parties inférieures sont blanches mais sont, selon le sexe et la sous-espèce, plus ou moins claires et teintées de chamois. Les flancs et le bas-ventre sont rouge orangé, avec des taches blanches sur les sous-caudales. Les différentes sous-espèces a ventre blanc se distinguent souvent par des nuances de teinte des flancs et des parties inférieures[2]. S. e. caesia, la plus répandue des sous-espèces occidentales, a les parties inférieures chamois orangé et une gorge et des joues blanches. Les autres sous-espèces de l'Ouest de l'Europe diffèrent par les tons précis de leurs parties inférieures, quand les formes du Sud-Est présentent un front et un sourcil blancs. S. e. sinensis et S. e. formosana, respectivement de Chine et de Taïwan, ont des parties inférieures entièrement chamoisées, sans la gorge blanche[2].


Plumage des femelles
La femelle est assez semblable au mâle, mais peut néanmoins s'en distinguer par ses parties inférieures un peu plus pâles, un trait oculaire tirant sur le marron et des flancs et un bas-ventre aux couleurs plus lavées[3]. Chez la sous-espèce S. e. asiatica, certains mâles ont les parties inférieures chamoisées comme les femelles et leur sexe est difficile à identifier sur le terrain[2].
Plumage des juvéniles
Le jeune ressemble à la femelle, mais son plumage est plus terne et ses pattes plus pâles[3]. Il est possible de déterminer avec certitude qu'un jeune est femelle lorsqu'il est âgé de 12 jours par ses flancs plus pâles et plus chamoisés, ou, chez certaines sous-espèces à poitrine blanche, par l'aspect plus crémeux de ses parties supérieures[4]. Les juvéniles prêts à l'envol ont une mue de quelques-unes de leur couvertures alaires à l'âge de huit semaines[2].
Mue
Les adultes connaissent une mue post-nuptiale complète qui s'effectue en près de 80 jours, entre fin mai et fin septembre. En Sibérie, cette mue est plus rapide et a lieu de juin à mi-septembre.
Autres caractéristiques
Le bec est robuste, gris foncé avec la base de la mandibule inférieure plus claire. L'iris est marron foncé, et les pattes et les doigts sont marron clair ou grisâtres[3]. En vol, l'apparence de l'oiseau est caractéristique, avec une tête pointue, des ailes arrondies et une queue courte et carrée.
Espèces similaires
Dans la plus grande partie de son aire de répartition, la Sittelle torchepot est la seule sittelle présente. Dans le Sud-Est de l'Europe et le Sud-Ouest asiatique, la Sittelle des rochers (S. tephronota) et la Sittelle de Neumayer (S. neumayer) peuplent les milieux rocailleux, sont plus grandes et plus pâles que l'espèce eurasiatique et n'ont pas de points blancs sur la queue. Dans la même zone, la Sittelle de Krüper (S. krueperi) est plus petite, avec une calotte sombre et une grande tache rousse sur la poitrine. Dans le Sud-Ouest de la Chine, la Sittelle des Naga (S. nagaensis) est très semblable à la torchepot, mais a les parties supérieures plus foncées, a moins de blanc sur la face et a des parties inférieures plus grisâtres[2]. La Sittelle de Sibérie (S. arctica) était autrefois considérée comme sous-espèce de la Sittelle torchepot mais en diffère assez nettement, étant plus grande, pâle, avec un trait oculaire plus court et moins épais, un bec plus long et au culmen droit, et plus de blanc dans la queue que n'importe quelle autre sous-espèce[5].
- Autres sittelles dont l'aire de répartition chevauche celle de la Sittelle torchepot
.jpg.webp) Sittelle des rochers (S. tephronota).
Sittelle des rochers (S. tephronota). Sittelle de Neumayer (S. neumayer).
Sittelle de Neumayer (S. neumayer).%252C_Achladeri_Forest%252C_Lesvos%252C_Greece%252C_16.04.2015_(2).jpg.webp) Sittelle de Krüper (S. krueperi).
Sittelle de Krüper (S. krueperi)..jpg.webp) Sittelle des Naga (S. nagaensis).
Sittelle des Naga (S. nagaensis). Sittelle de Sibérie (S. arctica).
Sittelle de Sibérie (S. arctica).
Comportement
Locomotion
En vol, l'apparence de l'oiseau est caractéristique, avec une tête pointue, des ailes arrondies et une queue courte et carrée. Le vol est rapide et généralement de courte durée, et la sittelle referme ses ailes entre deux battements[1].
Voix
La Sittelle torchepot est un oiseau aux cris sonores, qu'on entend à longueur d'année[6] ; elle est souvent d'abord repérée à sa voix. Son répertoire est varié, et comprend un simple zit aigu ou ziit plus étiré lorsque l'oiseau est en quête de nourriture, « un tuit énergique, très sonore, ou tchuit vaguement montant, souvent répété en groupes brefs avec de courtes pauses » si l'oiseau est excité ou en guise de cri d'alarme[7]. Le chant est variable, mais toujours sonore et constitué d'une lente série de sifflements clairs, légèrement modulés[8],[7]. Ils peuvent être montants en « vuih, vuih, vuih, vuih…, ou descendants viiu, viiu, viiu, viiu…, ou en trille rapide, vivivivivivivi… à sonorité claire »[7],[6]. Quelques sonagrammes de cris et de chants de la Sittelle de Sibérie (S. arctica) sont publiés en 1996[9], et la voix est décrite comme « différant nettement » de celle de la torchepot, mais sans précision supplémentaire[10],[11].
Alimentation
.jpg.webp)
La Sittelle torchepot se nourrit surtout d'insectes, notamment de chenilles et de coléoptères. En automne et en hiver, elle complète son régime alimentaire de fruits à coque et de graines, et notamment de noisettes et de faînes. Les parents nourrissent principalement leurs jeunes d'insectes et de peu de graines. L'oiseau trouve sa nourriture le long des troncs d'arbre et des branches, mais les petits rameaux sont aussi prospectés et la nourriture peut être trouvée au sol, notamment en dehors de la saison de reproduction. Comme toutes les sittelles, elle peut descendre des troncs la tête la première, ou y grimper. Certaines proies sont attrapées en vol, et la sittelle peut arracher des bouts d'écorce pour attraper des insectes, bien qu'elle ne puisse pas excaver dans le bois sain comme le ferait un pic. Les couples peuvent se mêler à des volées mixtes d'alimentation, si l'une passe près de leur territoire[3]. La Sittelle torchepot vient aux mangeoires et se montre alors agressive, repoussant les autres espèces d'oiseaux. Elle peut aussi visiter les tables et consommer de la graisse, du beurre, du fromage ou du pain[12],[13]. On l'a même observée dans un abattoir emporter des abats[13]. Elle coince les grosses noix et les gros insectes dans l'écorce pour les écraser à l'aide de son solide bec[14]. Une observation publiée en 2018 rapporte l'utilisation par la Sittelle torchepot d'un outil, en l'occurrence un petit morceau de bois, pour soulever les écorces à la recherche de nourriture dans un parc au Royaume-Uni. Ce type de comportement n'avait alors été observé que chez certaines espèces de sittelles d'Amérique du Nord[15].
La Sittelle torchepot fait des réserves de nourriture tout au long de l'année mais surtout en automne. Elle coince des graines dans l'écorce des arbres, parfois dans des murs ou au sol, et les cache à l'aide de lichen, de mousse ou de morceaux d'écorces. Les populations de Sibérie cachent des graines de Pin nain de Sibérie (Pinus pumila), parfois assez pour durer toute une année[3]. La Sittelle torchepot peut aussi cacher du pain, ainsi que des chenilles ou d'autres larves après les avoir martelées du bec pour les immobiliser[16]. Ces réserves sont une stratégie de long terme, et sont utilisées par temps froid, quand la nourriture se fait rare, jusqu'à trois mois après la cache. Les oiseaux utilisant des réserves ont une meilleure survie que les autres[17]. On a observé que les oiseaux évitaient d'utiliser leurs cachettes dans des conditions relativement bénignes, préférant les réserver pour les périodes les plus rudes[17]. Certaines de ses réserves n'ayant pas été consommées, il arrive que des tournesols ou des noisetiers se mettent à pousser sur des murs fendus ou sur l'écorce d'un arbre[18]. Les productions de fruits par les frênes sont très variables d'une année sur l'autre, et là où cet arbre représente une importante source de nourriture, le taux de survie des adultes n'est pas affecté lors des mauvaises saisons, alors que le nombre de juvéniles chute en automne, ceux-ci mourant de faim ou d'épuisement à force de se déplacer[19]. Des tendances similaires ont été observées là où le noisetier est l'espèce prédominante[20].
Reproduction

La Sittelle torchepot est monogame, et le couple occupe généralement son territoire à l'année[3]. Celui-ci couvre deux à dix hectares en Europe, mais jusqu'à 30,2 ha en moyenne dans les forêts de conifères de Sibérie, habitat sub-optimal[21]. Le mâle chante pour défendre son territoire et pour attirer une partenaire. Les deux membres du couple exécutent une parade nuptiale, comptant un vol léger et tremblotant, et le mâle fait également des vols en cercle avec la queue étalée et la tête relevée, et nourrit la femelle[1]. Malgré l'appariement à vie, des recherches génétiques effectuées en Allemagne ont montré qu'au moins 10 % des jeunes sont issus d'autres mâles, généralement de territoires adjacents[22].

Le nid est placé dans le trou d'un arbre, généralement un ancien nid de pic, mais parfois d'origine naturelle. Le nid est situé entre deux et vingt mètres du sol. Parfois, la femelle élargit l'entrée d'un trou existant dans le bois pourri. Si au contraire l'entrée est trop large, elle est maçonnée avec de la boue, de l'argile ou parfois de la bouse pour la réduire[3], pratique qui vaut à l'oiseau son nom de « torchepot ». Les nids aux entrées plus étroites sont moins victimes des prédateurs et ont donc un plus grand succès reproducteur[23]. Localement, une entrée étroite peut aussi éviter que des Étourneaux sansonnets (Sturnus vulgaris) s'approprient le nid[3],[12]. L'intérieur du nid est grand, avec une épaisse couche d'écorce de pins et divers copeaux de bois, parfois d'autres débris secs de végétaux en son fond, dans lesquels les adultes enfoncent les œufs ou les petits oisillons quand ils quittent le nid afin de limiter leur prédation. La femelle réalise l'essentiel de la construction, et maçonne souvent l'intérieur aussi. Cette tâche lui prend jusqu'à quatre semaines, et le nid est souvent réutilisé d'une année sur l'autre[3].
La femelle pond les œufs entre avril et juillet. La ponte compte généralement six à neuf œufs, bien qu'il puisse y en avoir jusqu'à treize. Ils mesurent en moyenne 19,5 × 14,4 mm pour S. e. caesia[2] et pèsent 2,3 g dont 6 % de coquille[24]. Ils sont blancs et tachetés de brun. La couvaison dure entre 13 et 18 jours, et est réalisée par la femelle seule, nourrie par le mâle. Les oisillons sont nidicoles et sont nourris par les deux parents. Ils sont prêts à l'envol à l'âge de 20-26 jours mais continuent à être alimentés par les adultes jusqu'à leur indépendance pendant huit à quatorze jours de plus. Il y a généralement une seule ponte par an, exceptionnellement deux[2],[24]. La Sittelle torchepot est plus réticente à utiliser les nichoirs artificiels que les autres oiseaux nichant dans des cavités[12]. Quand elle le fait, le nombre d'œufs et de jeunes à l'envol est plus élevé dans les grands nichoirs, alors qu'il ne sont pas liés à la taille de la cavité dans les emplacements de nidification naturels[25]. La sédentarité de cette espèce implique que les jeunes ne peuvent établir leur propre territoire qu'en trouvant un territoire inoccupé ou en remplaçant un adulte mort. En Europe, les jeunes se déplacent toujours vers un nouveau territoire, alors qu'en Sibérie, où les territoires sont beaucoup plus vastes, la plupart vivent dans le territoire d'adultes. Le taux de survie annuel (la proportion d'individus passant l'année) est de 42-47 % en Suède, 51-59 % en Belgique et 67 % en Sibérie[26]. L'espérance de vie de cette sittelle est de deux ans mais le record pour un oiseau sauvage est de 12 ans et 11 mois, au Royaume-Uni[27], et un individu suisse a vécu 10 ans et 6 mois[28].
Prédateurs et parasites

Dans la majeure partie de son aire de répartition européenne, le principal prédateur de la Sittelle torchepot est l'Épervier d'Europe (Accipiter nisus)[29],[30]. L'Autour des palombes (A. gentilis)[31], le Faucon hobereau (Falco subbuteo)[32], la Chouette hulotte (Strix aluco)[33], la Chevêchette d'Europe (Glaucidium passerinum), la Belette (Mustela nivalis)[34],[35] et la Martre (Martes martes)[18] sont aussi connus pour être prédateurs de l'espèce. Une étude suédoise a montré que 6,2 % des nids de sittelles de la zone d'étude étaient victimes de prédation. Les prédateurs n'ont pas été identifiés, mais sur la même zone, le principal prédateur des nids de mésanges était le Pic épeiche (Dendrocopos major)[36].
L'Étourneau sansonnet (Sturnus vulgaris) peut s'approprier le nid de la Sittelle torchepot, surtout lorsque le nid est haut placé, réduisant son succès reproducteur[36]. La Perruche à collier (Psittacula krameri), espèce introduite en Europe, peut aussi concurrencer cette sittelle pour l'accès au nid, mais les perruches ont plutôt tendance à habiter les bois fragmentés des villes quand les sittelles privilégient les grands peuplements de vieux chênes, ce qui limite la compétition. Une étude menée en Belgique en 2010 a conclu que ces oiseaux ne posaient pas de problèmes trop graves aux sittelles et que leur abattage n'était pas nécessaire[37].
L'acarien Ptilonyssus sittae est connu pour parasiter les cavités nasales de la Sittelle torchepot[38],[39]. Parmi les vers intestinaux, on compte les nématodes Capillaria parusi, Aonchotheca longifilla[40] et Aonchotheca caudinflata[41]. L'espèce est aussi l'hôte d'acanthocéphales comme Mediorhynchus micracanthus et de cestodes comme Anonchotaenia globata, Orthoskrjabinia bobica, Capiuterilepis naja, Hymenolepis clerci et Passerilepis passeris[41]. Parmi les trématodes trouvés chez S. europaea, on recense Platynosomum ventricosum, Echinochasmus beleocephalus, Laterotrema vexans, Mosesia caprimulgi, Urogonimus macrostomum, Leucochloridium phragmitophila, Plagiorchis elegans, Plagiorchis maculosus et Prosthogonimus ovatus[41]. De petites études en Slovaquie et en Espagne n'ont pas décelé de parasites sanguins, mais des prospections plus grandes réalisées en Espagne ont montré que cette sittelle pouvait être infectée par Plasmodium[42],[43],[44]. Haemoproteus sittae, protozoaire sanguin initialement décrit chez la Sittelle à poitrine blanche (Sitta carolinensis)[45], parasite aussi la Sittelle torchepot[46].
Répartition et habitat
Répartition
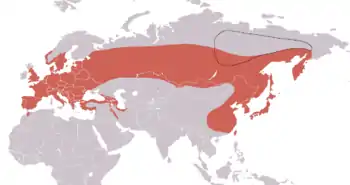
Les zones de reproduction de la Sittelle torchepot s'étendent dans l'hémisphère nord, à travers toute l'Eurasie tempérée, depuis la Grande-Bretagne (mais pas l'Irlande) jusqu'au Japon[2]. Elle vit entre les isothermes de juillet de 16 et 20 °C[1], sans dépasser au nord le 64e parallèle dans l'Ouest de la Russie et le 69e parallèle en Sibérie, là où la Sittelle de Sibérie (S. arctica) se rencontre aussi. En Russie également, elle se rencontre dans le Sud jusqu'aux alentours des 54e-55e parallèles, et également dans le Nord du Kazakhstan et de la Mongolie. En Europe, elle niche jusqu'aux côtes de la Méditerranée dans le Sud, mais pas sur les îles mésogéennes à l'exception de la Sicile. Elle peuple également quelques localités du Rif marocain et certaines zones côtières ou montagneuses du Proche-Orient. Dans l'Est, elle vit dans une majeure partie de la Chine orientale, sur Taïwan et en Corée[2]. Elle est erratique au Liban[47], sur les îles Anglo-Normandes et S. e. europaea a été signalée plusieurs fois en Finlande qui abrite normalement la sous-espèce S. e. asiatica[1].
Hivernage
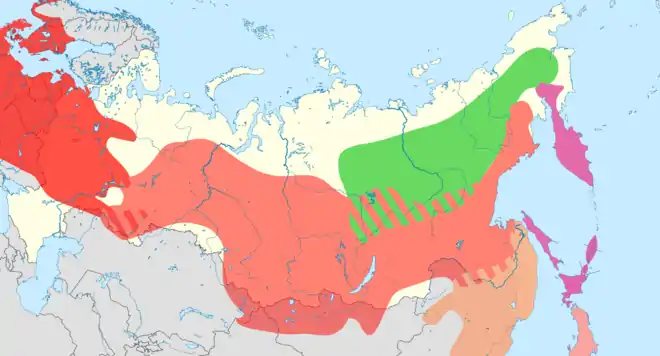
La plupart des populations sont sédentaires, à l'exception de la dispersion des jeunes après la saison de reproduction, et l'espèce se montre réticente à traverser ne serait-ce que de petites étendues d'eau. Les nicheurs du nord et de l'est sont dépendants de l'approvisionnement en graines par le Pin de Sibérie (Pinus sibirica), et lors des mauvaises saisons, la sous-espèce S. e. asiatica peut se déplacer vers l'ouest, dans le Nord de la Suède et en Finlande en automne, y restant parfois pour nicher. S. e. amurensis, du Sud-Est de la Russie et de Chine, descend souvent dans la péninsule coréenne en hiver[2].
|
|
|
Habitat

La Sittelle torchepot privilégie les vieilles forêts, avec de grands et vieux arbres, qui lui fournissent à la fois une nourriture abondante et des cavités pour nicher. En Europe, on la trouve plutôt dans les forêts mixtes ou de feuillus, et notamment celles contenant du chêne (Quercus). Cette espèce peut également vivre dans les parcs, les vieux vergers et autres endroits boisés tant que ceux-ci offrent une parcelle d'un hectare au moins d'habitat adapté. En montagne, la Sittelle torchepot peut peupler les forêts d'épicéas et de pins, et les pins sont également l'habitat privilégié sur Taïwan. Dans la majeure partie de la Russie, les conifères sont utilisés pour nicher, mais les densités de population sont relativement basses. Au Maroc, l'espèce niche dans les chênes, les Cèdres de l'Atlas (Cedrus atlantica) et les sapins (Abies). Parmi les habitats moins courants, on compte les genévriers nains (Juniper) en Mongolie et des milieux rocailleux dans quelques zones du Sud de la Sibérie[3]. La Sittelle torchepot est avant tout un oiseau de plaine dans le Nord de son aire de répartition, mais se trouve jusqu'à la limite des arbres en Suisse, à 1 200 m ou plus haut encore, et se reproduit même parfois à 1 800-2 100 m en Autriche. Elle niche à des altitudes équivalentes dans les montagnes de Turquie, du Moyen-Orient et d'Asie centrale. Cette espèce est, en revanche, principalement montagnarde dans le Sud du Japon et à Taïwan où elle vit respectivement entre 760 et 2 100 m et entre 800 et 3 300 m). Dans le Sud de la Chine, elle se trouve essentiellement à de basses altitudes, les montagnes étant occupées par la Sittelle des Naga (S. nagaensis)[3].
Systématique
Taxinomie

La Sittelle torchepot est décrite en 1758 par le naturaliste suédois Carl von Linné dans son ouvrage Systema Naturae et sous son nom scientifique actuel, Sitta europaea[49]. Cette description concerne la sous-espèce type, S. europaea europaea, au ventre blanc, qui se trouve dans le Sud de la Scandinavie où vit Linné, et dans le Nord-Est de l'Europe jusque dans l'Ouest de la Russie.
Le nom du genre dérive du grec ancien σίττη (sittè), trouvé dans l'Histoire des animaux d'Aristote, et peut-être dérivé du chant de la Sittelle torchepot ou plus probablement de la Sittelle de Neumayer (S. neumayer). Par la suite, de très nombreux taxons aujourd'hui considérés comme sous-espèces ou comme synonymes de S. europaea sont décrits, à commencer par S. europaea caesia, la sous-espèce aux parties inférieures chamoisées et à la gorge blanche peuplant l'essentiel de l'Europe de l'Ouest, décrite dès 1810 par l'ornithologue allemand Johann Wolf[48]. Dans le découpage en sous-genres du genre Sitta, peu utilisé, la Sittelle torchepot est placée dans Sitta (Sitta) Linnaeus, 1758[50]. La Sittelle torchepot forme une super-espèce avec la Sittelle de Sibérie (S. arctica), la Sittelle des Naga (S. nagaensis), la Sittelle du Cachemire (S. cashmirensis), la Sittelle indienne (S. castanea), la Sittelle de Blyth (S. cinnamoventris) et la Sittelle d'Indochine (S. neglecta), et les formes qui la composent se remplacent géographiquement les unes les autres à travers l'Eurasie[2]. La Sittelle de Sibérie (S. arctica) était jusqu'en 2006 généralement considérée comme une sous-espèce de la torchepot mais elle vit pourtant partiellement en sympatrie avec Sitta europaea sans s'hybrider (ou très peu) pour autant[11],[51].
Plus généralement tous les taxons formant le « groupe europaea » ne sont pas toujours reconnus comme espèces à part entière. Le groupe de sous-espèces « caesia », peuplant une grande partie de l'Europe jusqu'au Moyen-Orient, est parfois considéré comme distinct du groupe « europaea » (de Scandinavie et de Russie) avec une zone d'hybridation importante de la Baltique à la mer Noire[52].
Sous-espèces
De nombreuses formes ont été décrites chez la Sittelle torchepot, et leur validité en tant que sous-espèce est fluctuante, avec parfois plus d'une vingtaine reconnues. Elles sont regroupées en trois groupes : S. e. caesia, aux parties inférieures chamoisées mais à la gorge blanche, d'Europe, d'Afrique du Nord et du Moyen-Orient, S. e. europaea aux parties inférieures entièrement blanches, de Scandinavie, de Russie, du Japon et du Nord de la Chine et S. e. sinensis aux parties inférieures entièrement chamoisées, du Sud et de l'Est de la Chine et de Taïwan. Ces groupes auraient pu être isolés les uns des autres jusqu'à récemment, mais des oiseaux d'apparences intermédiaires se rencontrent aux zones de chevauchement. Selon le Congrès ornithologique international (version 6.4, 2016)[53] et Alan P. Peterson[54] il existe vingt-et-une sous-espèces :
- Sitta europaea caesia Wolf, 1810, de l'Ouest et centre de l'Europe ;
- Sitta europaea europaea Linnaeus, 1758, du Nord et de l'Est de l'Europe ;
- Sitta europaea hispaniensis Witherby, 1913, de la péninsule Ibérique et du Maroc ;
- Sitta europaea cisalpina Sachtleben, 1919, du versant Sud des Alpes suisses, dans toute l'Italie (dont la Sicile) et jusqu'en Croatie à l'est ;
- Sitta europaea levantina Hartert, 1905, du Sud de la Turquie, Nord de la Syrie et Nord du Liban ;
- Sitta europaea persica Witherby, 1903, du Sud-Est de la Turquie, Nord de l'Irak et Ouest de l'Iran ;
- Sitta europaea caucasica Reichenow, 1901, du Nord-Est de la Turquie jusqu'au Sud-Ouest de la Russie ;
- Sitta europaea rubiginosa Tschusi & Zarudny, 1905, du Nord de l'Iran et Azerbaïdjan ;
- Sitta europaea asiatica Gould, 1835, dans le centre et l'ouest du Sud de la Sibérie, du nord du Kazakhstan à l'Ouest de la Mongolie ;
- Sitta europaea baicalensis Taczanowski, 1882, de l'est de la Sibérie au centre de la Mongolie ;
- Sitta europaea albifrons Taczanowski, 1882, du Nord-Est de la Sibérie et Nord des îles Kouriles ;
- Sitta europaea sakhalinensis Buturlin, 1916, de Sakhaline ;
- Sitta europaea takatsukasai Momiyama, 1931, du centre et du Sud des îles Kouriles ;
- Sitta europaea clara Stejneger, 1887, du Sud des îles Kouriles et à Hokkaidō (Japon) ;
- Sitta europaea hondoensis Buturlin, 1916, de Honshū au Nord de Kyūshū (Japon) ;
- Sitta europaea amurensis Swinhoe, 1871, du Sud-Est de la Sibérie, Corée, centre et Sud du Japon et du Hebei jusqu'au Nord-Est de la Mandchourie ;
- Sitta europaea roseilia Bonaparte, 1850, du Sud de Kyushu ;
- Sitta europaea bedfordi Ogilvie-Grant, 1909, de Jeju-do ;
- Sitta europaea seorsa Portenko, 1955, du Nord-Ouest de la Chine ;
- Sitta europaea sinensis J. Verreaux, 1871, du centre-Nord et Est de la Chine ;
- Sitta europaea formosana Buturlin, 1911, de Taïwan
 Sitta europaea asiatica, avec le sourcil et la base du front blancs. Le brun-roux du croupion est peu étendu.
Sitta europaea asiatica, avec le sourcil et la base du front blancs. Le brun-roux du croupion est peu étendu. Sitta europaea europaea, individu à ventre blanc, comme en Scandinavie et dans l'Ouest de la Russie.
Sitta europaea europaea, individu à ventre blanc, comme en Scandinavie et dans l'Ouest de la Russie. Sitta europaea caesia, peuplant l'Europe continentale.
Sitta europaea caesia, peuplant l'Europe continentale. Sitta europaea sinensis, sous-espèce aux parties inférieures de couleurs prononcées.
Sitta europaea sinensis, sous-espèce aux parties inférieures de couleurs prononcées.
Hybrides observés
Différentes sous-espèces de la Sittelle torchepot peuvent s'hybrider quand leurs aires de répartition entrent en contact, voire avec d'autres espèces proches. Par ordre alphabétique, on a notamment observé les hybrides suivants[52] :
- Sitta (europaea) caesia × Sitta europaea europaea, quand Sitta (europaea) caesia est considérée en tant qu'espèce à part entière, est un hybride fréquent dans la nature (de la mer Baltique à la mer Noire) ;
- Sitta cashmirensis × Sitta (europaea) sinensis, hybride observé en captivité pour deux espèces aux répartitions disjointes ;
- Sitta castanea × Sitta (europaea) sinensis, hybride observé en captivité pour deux espèces aux répartitions disjointes ;
- Sitta europaea europaea × Sitta (europaea) sinensis, quand Sitta (europaea) sinensis est considérée en tant qu'espèce à part entière, hybride fréquent dans la nature (Nord-Est de la Chine) et correspondant peut-être à la sous-espèce Sitta europaea amurensis ;
- Sitta nagaensis × Sitta (europaea) sinensis, hybride supposé, mais probablement fréquent (Sud de la Chine), et correspondant peut-être à la sous-espèce Sitta nagaensis montium.
Phylogénie
| Extrait de la phylogénie des sittelles selon Päckert et al. (2020)[55] : | |
|
En 2014, Éric Pasquet et al. publient une phylogénie fondée sur l'ADN nucléaire et mitochondrial de 21 espèces de sittelles[56]. Le groupe « europaea » est rapproché des deux sittelles des milieux rocheux, la Sittelle de Neumayer (S. neumayer) et la Sittelle des rochers (S. tephronota), et ces deux clades divergent l'un de l'autre il y a treize millions d'années. Au sein du groupe « europaea », la Sittelle de l'Himalaya (S. himalayensis) — et par conséquent la Sittelle du Victoria (S. victoriae), bien qu'elle ne soit pas incluse dans l'étude — apparaît comme basale, et la Sittelle torchepot est rapprochée de la Sittelle des Naga (S. nagaensis) et de la Sittelle du Cachemire (S. cashmirensis). La Sittelle indienne (S. castanea), la Sittelle de Blyth (S. cinnamoventris), la Sittelle d'Indochine (S. neglecta) et la Sittelle de Sibérie (S. arctica) ne sont pas incluses dans l'étude. Toutes les espèces du groupe « europaea » maçonnent l'entrée de leur nid[56]. En 2020 paraît une nouvelle phylogénie couvrant le genre de façon plus exhaustive : elle inclut notamment les quatre espèces mentionnées précédemment. Elle utilise 3 gènes mitochondriaux et deux nucléaires. Les trois espèces du sud du continent asiatique (Sittelles indienne, de Blyth et d'Indochine) sont rapprochées de la Sittelle du Cachemire, mais étonnamment la Sittelle de Sibérie est située sur une branche propre, assez distante de la Sittelle torchepot dont elle était pourtant longtemps considérée comme une sous-espèce[55].
L'étude de Päckert et al. (2020) comprend également un échantillonnage assez exhaustif des sous-espèces de la Sittelle torchepot. Elle met en évidence trois grands groupes de sous-espèces, pas parfaitement recouvrants avec les groupes traditionnellement distingués (voir Liste des sous-espèces de la Sittelle torchepot) sur la base de la coloration des parties inférieures. Un premier groupe concerne la « Sittelle d'Europe », qui inclut toutes les sous-espèces européennes qu'elles soient à ventre chamoisé ou ventre blanc, ainsi que les sous-espèces du Proche-Orient (Turquie, Iran, Irak…)[55]. Il est rapproché du groupe de la « Sittelle d'Asie », incluant les sous-espèces peuplant le Nord du continent asiatique, du Kazakhstan jusqu'en Corée et au Japon, toutes à ventre blanc. Enfin, un troisième groupe de sous-espèces, la « Sittelle orientale », inclut les sous-espèces asiatiques vivant plus au sud, dans le Nord et l'Est de la Chine et à Taïwan[55], qui ont les parties inférieures entièrement chamoisées, joues comprises.
La Sittelle torchepot et l'homme
Menaces et protection
La Sittelle torchepot est largement répandue, son aire de répartition connue étant proche de 23 300 000 km2 et couvrant une grande partie des habitats disponibles et adaptés[57],[58]. On estime qu'il existe en Europe de 22,5 à 57 millions d'individus, soit une population totale comptant entre 45,9 et 228 millions d'individus, les effectifs étant globalement stables. La Chine, Taïwan, la Corée, le Japon et la Russie comptent au moins chacun entre 10 000 et 100 000 couples reproducteurs[57]. Pour ces raisons, l'espèce est considérée comme de « préoccupation mineure » par l'Union internationale pour la conservation de la nature[47]. C'est une espèce commune sur l'essentiel de son aire de répartition, bien que les densités soient plus faibles le plus au nord et dans les forêts de conifères. Les effectifs de Sibérie fluctuent selon les disponibilités de pommes de pin[2]. Lors des dernières décennies, la Sittelle torchepot a colonisé l'Écosse et les Pays-Bas, et a augmenté sa distribution au pays de Galles, dans le Nord de l'Angleterre, en Norvège et dans le Haut Atlas. S. e. asiatica se reproduit parfois en Finlande et dans le Nord de la Suède au gré d'irruptions. Sa dépendance aux gros arbres peut causer des déclins locaux en cas de destruction ou de fragmentation de vieux boisements[1]. Une étude de 2009 a essayé de prédire l'impact que pourront avoir les changements climatiques sur la répartition de plusieurs espèces de sittelles en Asie, en modélisant deux scénarios ; la Sittelle torchepot pourrait voir sa distribution diminuer de 64,7 à 64,8 %[59].
Dans la culture
Dans l'Edda poétique, recueil de poèmes du XIIIe siècle sur la mythologie nordique, un poème narre l'histoire d'un jeune aventurier, Sigurth, qui allié au nain Regin, tue le dragon Fafnir. Il absorbe un peu de sang du dragon et se met alors à comprendre le langage des sittelles autour de lui. Elles le préviennent de la traîtrise du nain qui veut le tuer pour s'emparer seul du trésor du dragon. Sigurth tue alors Regin puis, guidé par les sittelles, se rend au siège du royaume avec le trésor, où il épousera une belle princesse[60].
Le naturaliste français Georges-Louis Leclerc de Buffon rapporte et commente dans son Histoire naturelle un commentaire de Pierre Belon à propos des relations entre les partenaires sittelles : « « Les paysans ont observé, dit Belon, que le mâle bat sa femelle quand il la trouve qu'elle s'est départie de lui, dont ils ont fait un proverbe pour un qui se conduit sagement en ménage, qu'il ressemble au torche-pot » mais quoi qu'il en soit de la sagesse des maris, je ne crois point que, dans ce cas particulier, celui-ci ait la moindre intention de battre sa femme; je croirais bien plutôt que cette femelle, qui se fait désirer si long-temps avant la ponte, est la première à se retirer après l'éducation de la famille, et que lorsque le mâle la rencontre après une absence un peu longue, il l'accueille par des caresses d'autant plus vives, même un peu brusques, et que des gens qui n'y regardent pas de si près, auront prises pour de mauvais traitements »[61].
Voir aussi
Références taxinomiques
- (en) Référence Congrès ornithologique international : (consulté le )
- (en) Référence Zoonomen Nomenclature Resource (Alan P. Peterson) : Sitta europaea dans Sittidae (consulté le )
- (fr+en) Référence Avibase : Sitta europaea (+ répartition) (consulté le )
- (fr+en) Référence ITIS : Sitta europaea Linnaeus, 1758 (consulté le )
Liens externes
- Ressources relatives au vivant :
- Avibase
- Global Biodiversity Information Facility
- Oiseaux.net
- TAXREF (INPN)
- (en) Animal Diversity Web
- (cs + en) BioLib
- (en) BirdFacts
- (en) BirdLife International
- (sv) Dyntaxa
- (en) EPPO Global Database
- (en) EU-nomen
- (en) European Nature Information System
- (en) Fauna Europaea
- (mul + en) iNaturalist
- (en) Interim Register of Marine and Nonmarine Genera
- (en) Internet Bird Collection
- (cs) Nálezová databáze ochrany přírody
- (en) NBN Atlas
- (nl) Nederlands Soortenregister
- (en) Système d'information taxonomique intégré
- (en) Union internationale pour la conservation de la nature
- (en) Xeno-canto
- Notice dans un dictionnaire ou une encyclopédie généraliste :
- (fr) Référence Oiseaux.net : Sitta europaea Linnaeus, 1758 (+ répartition) (consulté le )
- (en) « Sitta europaea Linnaeus, 1758 », sur xeno-canto.org (consulté le )
- (en) Référence Animal Diversity Web : Sitta europaea (consulté le )
- (fr) Référence CITES : taxon Sitta europaea (sur le site du ministère français de l'Écologie) (consulté le )
- (en) Référence Fauna Europaea : Sitta europaea Linnaeus, 1758 (consulté le )
- (en) Référence NCBI : Sitta europaea (taxons inclus) (consulté le )
- (en) Référence Catalogue of Life : Sitta europaea Linnaeus, 1758 (consulté le )
- (en) Référence UICN : espèce Sitta europaea (consulté le )
Bibliographie
- Simon Harrap, Chickadees, Tits, Nuthatches & Treecreepers, (œuvre littéraire), Princeton University Press, Princeton,
- Erik Matthysen, The Nuthatches, (œuvre littéraire), T. & A. D. Poyser, Londres,
- (en) David Snow et Christopher M. Perrins, The Birds of the Western Palearctic (BWP) : Concise edition, Oxford, Oxford University Press, (ISBN 0-19-854099-X)
- (en) Josep del Hoyo, Andrew Elliott, Jordi Sargatal et David A. Christie, « Eurasian Nuthatch », sur Handbook of the Birds of the World Alive, Lynx Edicions (consulté le )
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Eurasian nuthatch » (voir la liste des auteurs).
- 1 2 3 4 5 6 Snow et Perrins (1998), p. 1402-1404, chapitre « Eurasian Nuthatch »
- 1 2 3 4 5 6 7 8 9 10 11 12 Harrap (1995), p. 109-114, chapitre « Eurasian Nuthatch »
- 1 2 3 4 5 6 7 8 9 10 11 Handbook of the Birds of the World Alive
- ↑ Matthysen (1998), p. 31
- ↑ Matthysen (1998), p. 27
- 1 2 Frédéric Jiguet, Jan Pedersen et Lars Svensson, « Chants d'Oiseaux », Larousse, 2012, (ISBN 978-2-03-587204-3), p. 201.
- 1 2 3 (fr) Lars Svensson (trad. du suédois par Guilhem Lesaffre et Benoît Paepegaey, ill. Killian Mullarney et Dan Zetterström), Le guide ornitho : Le guide le plus complet des oiseaux d'Europe, d'Afrique du Nord et du Moyen-Orient : 900 espèces, Paris, Delachaux et Niestlé, coll. « Les Guides du Naturaliste », , 446 p. (ISBN 978-2-603-01695-4), p. 348-349
- ↑ Oiseaux.net
- ↑ (ru) Vladimir Vladimirovich Leonovitch, G.V. Demina et Olga Dmitriyevna Veprintseva, « Обыкновенный поползень (Sitta europaea L.) и «малые» поползни (S. villosa Verreaux, S. krueperi Pelzeln): вопросы систематики и филогении [Sitta europaea L. & «small» nuthatches S. villosa Verreaux and S. krueperi Pelzeln: problems of taxonomy and phylogeny] », бюллетень московского общества испытателей природы. отдел биологический [Bulletin de la Société des naturalistes de Moscou - division Biologie], vol. 101, no 1, , p. 37–49
- ↑ (ru) Leonidas Aleksandrovitch Portenko, Фауна Анадырского края, Léningrad, , 211 p.
- 1 2 3 (en) Yaroslav Red'kin et Maria V. Konovalova, « Systematic notes on Asian birds. 63. The eastern Asiatic races of Sitta europaea Linnaeus, 1758 », Zoologische Mededelingen, vol. 80, no 15, , p. 241-261 (ISSN 0024-0672, lire en ligne)
- 1 2 3 (en) Bodil Enokssson, « Nuthatch », dans David Wingham Gibbons, James B. Reid et Robert A. Chapman, The New Atlas of Breeding Birds in Britain and Ireland: 1988–1991, Londres, T. & A.D. Poyser, (ISBN 0-85661-075-5), p. 376
- 1 2 Matthysen (1998), p. 51
- ↑ Harrap (1995), p. 16
- ↑ (en) Christian Rutz et Simon Deans, « Nuthatch uses tool in London park », Ethology, vol. 124, no 2, , p. 135–138 (DOI 10.1111/eth.12714)
- ↑ (en) T.J. Richards, « Concealment and recovery of food by birds, with some relevant observations on squirrels », British Birds, vol. 51, no 12, , p. 497–508
- 1 2 (en) Jan-Åke Nilsson et Hans Källander Owe Persson, « A prudent hoarder: effects of long-term hoarding in the European nuthatch, Sitta europaea », Behavioral Ecology, OUP, vol. 4, no 4, , p. 369-373 (ISSN 1045-2249 et 1465-7279, OCLC 41963900, DOI 10.1093/BEHECO/4.4.369)
- 1 2 Bernhard Grzimek (dir.), Le Monde animal en 13 volumes : Encyclopédie de la vie des bêtes, t. IX : Oiseaux 3, Zurich, Éditions Stauffacher S.A., , 1re éd., 594 p., chap. XIII (« Mésanges, Sittelles et Grimpereaux »), p. 298-303
- ↑ (en) Erik Matthysen, « Nuthatch Sitta europaea demography, beech mast, and territoriality », Ornis Scandinavica, Wiley-Blackwell, vol. 20, no 4, , p. 278-282 (ISSN 0030-5693, DOI 10.2307/3676492, JSTOR 3676492)
- ↑ (en) Bodil Enoksson, « Autumn territories and population regulation in the Nuthatch Sitta europaea: an experimental study », Journal of Animal Ecology, Wiley-Blackwell, vol. 59, no 3, , p. 1047 (ISSN 0021-8790 et 1365-2656, OCLC 42799265, DOI 10.2307/5030, JSTOR 5030)
- ↑ (en) Vladimir V. Pravosudov, « Social organization of the Nuthatch Sitta europaea asiatica », Ornis Scandinavica, Wiley-Blackwell, vol. 24, no 4, , p. 290-296 (ISSN 0030-5693, DOI 10.2307/3676790, JSTOR 3676790)
- ↑ (en) Gernot Segelbacher, Daniela Kabisch, Michael Stauss et Jürgen Tomiuk, « Extra-pair young despite strong pair bonds in the European Nuthatch (Sitta europaea) », Journal für Ornithologie, Berlin, Springer Science+Business Media et inconnu, vol. 146, no 2, , p. 99-102 (ISSN 0021-8375 et 1439-0361, OCLC 754654105 et 1588080, DOI 10.1007/S10336-004-0062-5)
- ↑ (en) Tomasz Wesołowski et Patryk Rowiński, « Breeding behaviour of Nuthatch Sitta europaea in relation to natural hole attributes in a primeval forest », Bird Study, Taylor & Francis, vol. 51, no 2, , p. 143-155 (ISSN 0006-3657, 1944-6705 et 0523-6894, DOI 10.1080/00063650409461346)
- 1 2 (en) « Nuthatch Sitta europaea [Linnaeus, 1758] », sur Bird Facts, British Trust for Ornithology (consulté le )
- ↑ (en) Vladimir V. Pravosudov, « Clutch size and fledging rate in the Eurasian Nuthatch breeding in natural cavities is unrelated to nest cavity size (Sin relación el tamaño de la cavidad y el tamaño de la camaday la tasa de pichones que dejan el nido de Sitta europea) », Journal of Field Ornithology, vol. 66, no 2, , p. 231–235 (JSTOR 4514008)
- ↑ Matthysen (1998), p. 149, chapitre 9 « Population Dynamics »
- ↑ (en) « European Longevity Records », EURING (consulté le )
- ↑ (en) « Eurasian Nuthatch Sitta europaea », sur Birds of Switzerland, Swiss Ornithological Institute (consulté le )
- ↑ Matthysen (1998), p. 148
- ↑ (en) Bogumila Jedrzejewska et Wlodzimierz Jedrzejewski, Predation in Vertebrate Communities : The Bialowieza Primeval Forest as a Case Study, Berlin, Springer, , 450 p. (ISBN 978-3-540-64138-4, lire en ligne)
- ↑ Jedrzejewska et Jedrzejewski (1998), p. 252
- ↑ Jedrzejewska et Jedrzejewski (1998), p. 282
- ↑ Jedrzejewska et Jedrzejewski (1998), p. 286
- ↑ Jedrzejewska et Jedrzejewski (1998), p. 294
- ↑ Jedrzejewska et Jedrzejewski (1998), p. 200
- 1 2 (en) Sven G. Nilsson, « The evolution of nest-site selection among hole-nesting birds: the importance of nest predation and competition », Ornis Scandinavica, vol. 15, no 3, , p. 167–175 (JSTOR 3675958)
- ↑ (en) Diederik Strubbe, Erik Matthysen et Catherine H. Graham, « Assessing the potential impact of invasive ring-necked parakeets Psittacula krameri on native nuthatches Sitta europeae in Belgium », Journal of Applied Ecology, vol. 47, no 3, , p. 549–557 (DOI 10.1111/j.1365-2664.2010.01808.x)
- ↑ (fr) Z. Fieder et Iulia Mironescu, « Contribution à la connaissance des rhinonyssides parasites nasicoles de la sittelle Sitta europaea caesia Wolf », Revue Roumaine de Biologie (Série de Zoologie), vol. 15, , p. 17–22 (lire en ligne)
- ↑ (en) A. Fain, W. Sixl et Ch. Moritsch, « The nasal mites of the family Rhinonyssidae with description of a new species (Acarina) », Mitteilungen der Abteilung für Zoologie am Landesmuseum Joanneum, vol. 3, no H1, , p. 129-137 (lire en ligne)
- ↑ (en) Carter T. Atkinson, Nancy J. Thomas et D. Bruce Hunter, Parasitic Diseases of Wild Birds, Londres, Wiley-Blackwell, , 595 p. (ISBN 978-0-8138-2081-1 et 0-8138-2081-2), p. 483
- 1 2 3 (en) « Host-parasite database - Sitta europaea », Londres, Natural History Museum (consulté le )
- ↑ (en) Kateřina Hauptmanová, Václav Benedikt et Ivan Literák, « Blood parasites in passerine birds in Slovakian east Carpathians », Acta Protozoologica, vol. 45, , p. 105–109 (lire en ligne)
- ↑ (en) Santiago Merino, Jaime Potti et Juan A. Fargallo, « Blood parasites of passerine birds from central Spain », Journal of Wildlife Diseases, Wildlife Disease Association (d), vol. 33, no 3, , p. 638-641 (ISSN 0090-3558 et 1943-3700, OCLC 1587351, PMID 9249714, DOI 10.7589/0090-3558-33.3.638, lire en ligne)
- ↑ (en) Josué Martinez-de la Puente, Javier Martinez, Juan Rivero-de Aguilar, Jessica Herrero et Santiago Merino, « On the specificity of avian blood parasites: revealing specific and generalist relationships between haemosporidians and biting midges », Molecular Ecology, vol. 20, , p. 3275-3287 (lire en ligne)
- ↑ (en) Gordon F. Bennett, « New species of haemoproteids from the avian families Paridae and Sittidae », Revue canadienne de zoologie, vol. 67, no 11, , p. 2685-2688 (DOI 10.1139/z89-379)
- ↑ (en) Gediminas Valkiunas, Avian Malaria Parasites and other Haemosporidia, CRC Press, , 946 p. (lire en ligne), p. 502-503
- 1 2 Union internationale pour la conservation de la nature
- 1 2 (de) Bernhard Meyer et Johann Wolf, Taschenbuch der deutschen Vögelkunde, oder, Kurze Beschreibung aller Vögel Deutschlands, vol. 1, (lire en ligne), p. 128
- ↑ (la) Carl von Linnaeus, Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, t. I, Holmiae (Laurentii Salvii), , 10e éd., 824 p. (lire en ligne), p. 115 :
« SITTA. Roſtrum ſubcultrato-conicum, rectum, porrectum: integerrimum, mandibula ſuperiore obtuſiuſcula. Lingua lacero-emarginata.
europaea. S. rectricibus nigris: lateralibus quatuor infra apicem albis. » - ↑ Matthysen (1998), p. 269-270, Appendix I « Scientific and Common Names of Nuthatches
- ↑ (en) Robert M. Zink, Sergei V. Drovetski et Sievert Rohwer, « Selective neutrality of mitochondrial ND2 sequences, phylogeography and species limits in Sitta europaea », Molecular Phylogenetics and Evolution, vol. 40, no 3, , p. 679-686 (PMID 16716603, DOI 10.1016/j.ympev.2005.11.002, lire en ligne)
- 1 2 (en) Eugene M. McCarthy, Handbook of Avian Hybrids of the World, Oxford, Oxford University Press, , 608 p. (ISBN 978-0-19-518323-8, lire en ligne), « Nuthatches - Family Sittidae », p. 246-247
- ↑ Congrès ornithologique international
- ↑ Alan P. Peterson
- 1 2 3 4 (en) Martin Päckert, Marcella Bader-Blukott, Berit Künzelmann, Sun Yue-Hua, Yu-Cheng Hsu, Christian Kehlmaier, Frederik Albrecht, Juan Carlos Illera et Jochen Martens, « A revised phylogeny of nuthatches (Aves, Passeriformes, Sitta) reveals insight in intra- and interspecific diversification patterns in the Palearctic », Vertebrate Zoology, vol. 70, no 2, , p. 241-262 (ISSN 1864-5755 et 2625-8498, DOI 10.26049/VZ70-2-2020-10)
- 1 2 (en) Éric Pasquet, Frederick Keith Barker, Jochen Martens, Annie Tillier, Corinne Cruaud et Alice Cibois, « Evolution within the nuthatches (Sittidae: Aves, Passeriformes): molecular phylogeny, biogeography, and ecological perspectives », Journal of Ornithology, Springer Science+Business Media, vol. 155, no 3, , p. 755-765 (ISSN 2193-7192 et 2193-7206, DOI 10.1007/S10336-014-1063-7)
- 1 2 (en) « Wood Nuthatch - BirdLife Species Factsheet », BirdLife International (consulté le )
- ↑ (en) Shaily Menon, Zafar-Ul Islam, Jorge Soberón et A. Townsend Peterson, « Preliminary analysis of the ecology and geography of the Asian nuthatches (Aves: Sittidae) », Wilson Journal of Ornithology, vol. 120, , p. 692–699
- ↑ (en) Shaily Menon, M. Zafar-ul Islam et A. Townsend Peterson, « Projected climate change effects on nuthatch distribution and diversity across Asia », The Raffles Bulletin of Zoology, vol. 57, no 2, , p. 569-575
- ↑ (fr) « Mythes et légendes », Oiseaux de collection, Paris, Eaglemoss International, no 10 « La sittelle », , p. 13
- ↑ (fr) Georges-Louis Leclerc de Buffon, Histoire Naturelle, vol. 5, Paris, (lire en ligne), p. 460-475