Théropodes

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Classe | Sauropsida |
| Super-ordre | Dinosauria |
| Ordre | Saurischia[alpha 1] - [alpha 2] |
Sous-ordre ou Clade
Clades de rang inférieur
- † Daemonosaurus ?
- † Eodromaeus
- † Erythrovenator
- † Tawa ?
- † Velocipes ?
- Neotheropoda (inclut Aves)
Les Théropodes (Theropoda) forment un clade de tétrapodes bipèdes comprenant la quasi-totalité des grands dinosaures carnassiers[3] et l'unique clade actuel des oiseaux. Ce groupe polymorphe de dinosaures souvent carnivores, de tailles très variables (de 40 cm de long et 150 g pour les maniraptors jusqu'à 15 m de long et 11 tonnes pour les Spinosaures), occupe les trois niches écologiques de milieux terrestres, aériens et aquatiques.
Étymologie
Le nom « Theropoda » dérive de deux racines grecques, θήρ / thêr (« bête sauvage ») et πούς, ποδός / poús, podós (« pied »). Nommés de cette façon par Othniel Charles Marsh en 1881, les théropodes, dits « à pieds de bête » au sens étymologique, étaient à l'époque distingués des Ornithopoda (nommés en 1871), « à pieds d'oiseaux », et des Sauropoda (nommés en 1878), « à pieds de reptiles », tous deux créés par Marsh.
Par la suite, ce concept se révéla erroné car l'on sait actuellement que ce sont des théropodes du Jurassique qui ont donné naissance aux oiseaux, et non les Ornithopoda. Une erreur similaire fut également commise par Harry Govier Seeley qui distingua selon la forme du bassin les dinosaures à bassin d'oiseau (Ornithischia) et les dinosaures à bassin de reptiles (Saurischia), alors que les oiseaux appartiennent bien au clade des Saurischia .
Histoire évolutive

%252C_cabo_de_Buena_Esparanza%252C_Sud%C3%A1frica%252C_2018-07-23%252C_DD_87.jpg.webp)
Comme les autres groupes de dinosaures, les théropodes apparaissent au Trias supérieur (Carnien) et acquièrent une distribution géographique globale à partir du Jurassique inférieur[4]. Les oiseaux, qui sont issus de petits Maniraptora du Jurassique moyen, sont les seuls dinosaures (et donc les seuls théropodes) à avoir survécu à l'extinction (ou crise, selon les auteurs) Crétacé/Paléogène (K/Pg) ce qui leur a ainsi permis de prospérer et de se diversifier considérablement durant le Cénozoïque[5]. En incluant les oiseaux, les théropodes forment un des groupes de vertébrés les plus florissants et le clade des dinosaures le plus diversifié morphologiquement[6]. Ayant évolué pendant plus de 230 millions d'années au côté des mammifères qui apparaissent en même temps qu'eux, ils sont avec les mammifères les seuls vertébrés à avoir occupé les trois niches écologiques de milieux terrestres, aériens et aquatiques[7]. Les théropodes non-aviens, qui incluent tous les membres de ce clade mis à part les oiseaux (Avialae), furent un des groupes de carnivores dominant dans les écosystèmes du Mésozoïques du monde entier[6]. Contrairement à une idée reçue, un grand nombre de théropodes non-aviens n'étaient pas carnassiers et plusieurs clades de théropodes coelurosaures comme les Therizinosauria, les Ornithomimosauria et les Oviraptorosauria semblent avoir été exclusivement herbivores[8]. Comme l'intégralité des autres clades des dinosaures, les théropodes non-aviens disparaissent à la fin du Mésozoïque, bien qu'ils aient été particulièrement abondants à l'extrême fin du Crétacé[9].
Description

Les théropodes non-aviens (c'est-à-dire, à l'exception du clade des Avialae — dont les oiseaux) sont aisément reconnaissables par leur posture bipède et leurs membres antérieurs et postérieurs pourvus de griffes acérées. Leur évolution conduit à des membres postérieurs tridactyles, assurant le soutien du corps et la locomotion, tandis que les membres antérieurs, plus courts (perte de leur fonction locomotrice terrestre quadrupède, cette libération ayant permis leur utilisation pour une nouvelle fonction, le vol, chez les théropodes aviens), sont probablement adaptés à la saisie de proies. La grande majorité des théropodes basaux, à savoir les Coelophysoidea, les Ceratosauria, les Megalosauroidea, les Allosauroidea et les Tyrannosauroidea, possède un crâne dont les mâchoires portent des dents pointues typiques d'une alimentation carnivore. La posture bipède et les dents en forme de lames sont deux caractères ancestraux que l'on retrouve chez les archosaures plus primitifs[3]. Bien que la plupart des théropodes basaux fussent des mangeurs de viande, un grand nombre de théropodes non-aviens s'adaptèrent à d'autres régimes alimentaires et devinrent ultérieurement piscivores[10] et surtout herbivores[11],[12],[8]. Plusieurs taxons ont été également considérés comme des omnivores[13] ainsi que des dinosaures susceptibles de filtrer l'eau pour se nourrir[14].
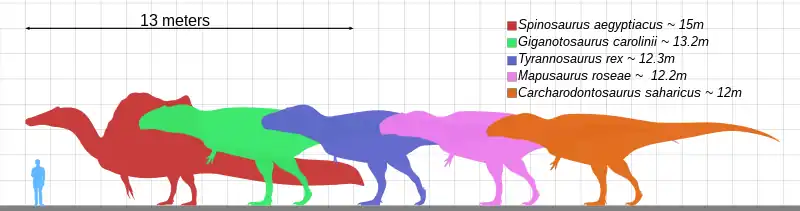
Il existe une grande variabilité de taille parmi les théropodes non-aviens, des genres tels que Spinosaurus ont une longueur de 15 mètres pour un poids de 6.3 à 7.5 tonnes ou encore Giganotosaurus avec une longueur comprise entre 12 et 13.2 mètres pour un poids variant entre 6.8 et 8.2 tonnes [15],[16] alors que d'autres théropodes non-aviens comme Epidexipteryx[17] et Anchiornis[18] ne devaient pas dépasser 40 centimètres de long pour un poids de quelque 150 grammes.
Des théropodes non-aviens récemment découverts tels que Incisivosaurus[19] pourvu de grandes dents au niveau de la mâchoire supérieure, Limusaurus[11] au long bec corné, Microraptor[20] et ses quatre membres ailés et Epidendrosaurus[21] muni de très longs doigts démontrent également une grande variété de morphologies au sein de ce clade.

La forme du crâne des théropodes est particulièrement variée puisque la tête robuste des Tyrannosauridae et des Carcharodontosauridae contraste avec le crâne long et frêle des Spinosauridae, et le crâne court et édenté des Oviraptorosauria. Un grand nombre de théropodes était également muni de crêtes et cornes en tous genres, tels que le Dilophosaurus et d'Oviraptor pourvus d'une crête haute sur toute la tête, de Ceratosaurus et de Spinosaurus ornés d'une petite crête nasale, de Cryolophosaurus muni d'une haute crête en forme de houppe sur l'extrémité postérieure de la tête et de Carnotaurus et Majungasaurus dont le haut du front était garni de petites cornes.

La morphologie des membres antérieurs est également variée. Les Spinosauridae aux bras puissants diffèrent des Ornithomimosauria aux bras longs et grêles, des Tyrannosauridae aux bras atrophiés à deux doigts et des Alvarezsauridae aux membres étrangement réduits à un seul doigt. La forme des griffes des théropodes est elle-même très diverse puisque l'on trouve de larges griffes en forme de faux chez les Spinosauroidea, de longues griffes en forme de lame chez les Therizinosauridae et des griffes courtes et robustes chez les Alvarezsauridae. Enfin, certains théropodes non-aviens étaient déjà munis d'un duvet et même de plumes, en témoigne la découverte des duveteux Dilong, Beipiaosaurus et Sinocalliopteryx et des dinosaures à plumes Sinornithosaurus, Sinosauropteryx, Caudipteryx et Protarchaeopteryx. Le vol plané était même déjà acquis par certains théropodes tels que Microraptor et Cryptovolans qui étaient munis de quatre ailes (une à chaque membre). Néanmoins, et malgré la présence de plumes sur leur corps, ces théropodes ne sont pas des oiseaux (ils en sont par contre des proches parents).
Systématique
Taxinomie
Selon BioLib (24 mai 2016)[22] :
- super-famille Abelisauroidea Bonaparte & Novas, 1985 †
- famille Abelisauridae Bonaparte & Novas, 1985 †
- famille Noasauridae Bonaparte & Powell, 1980 †
- super-famille Allosauroidea †
- famille Allosauridae Marsh, 1878 †
- famille Carcharodontosauridae Stromer, 1931 †
- famille Metriacanthosauridae Paul, 1988 †
- famille Neovenatoridae Benson, Carrano & Brusatte, 2010 †
- super-famille Alvarezsauroidea Bonaparte, 1991 †
- famille Alvarezsauridae Bonaparte, 1991 †
- super-famille Caenagnathoidea Sternberg, 1940
- famille Caenagnathidae Sternberg, 1940 †
- famille Oviraptoridae Barsbold, 1976 †
- super-famille Megalosauroidea Huxley, 1869
- famille Megalosauridae Huxley, 1869 †
- famille Spinosauridae Stromer, 1915 †
- famille Piatnitzkysauridae Carrano, Benson & Sampson, 2012 †
- super-famille Therizinosauroidea Russell, 1997 †
- famille Alxasauridae Russell & Dong, 1995 †
- famille Therizinosauridae Maleev, 1954 †
- super-famille Tyrannosauroidea Osborn, 1906 †
- famille Coeluridae Marsh, 1881 †
- famille Dryptosauridae Marsh, 1890 †
- famille Proceratosauridae Rauhut, Milner & Moore-Fay, 2010 †
- famille Tyrannosauridae Osborn, 1906 †
- famille Archaeopterygidae Huxley, 1871 †
- famille Caudipteridae Ji et al., 1998 †
- famille Ceratosauridae Marsh, 1884 †
- famille Coelophysidae Nopcsa, 1923 †
- famille Compsognathidae Cope, 1871 †
- famille Deinocheiridae Osmólska & Roniewics, 1970 †
- famille Dilophosauridae Charig & Milner, 1990 †
- famille Dromaeosauridae Matthew & Brown, 1922 †
- famille Ornitholestidae Paul, 1988 †
- famille Ornithomimidae Marsh, 1890 †
- famille Scansoriopterygidae Czerkas & Yuan, 2002 †
- famille Troodontidae Gilmore, 1924 †
Phylogénie
Les théropodes non-aviens ont reçu beaucoup d'attention de la part des paléontologues des vertébrés durant ces trente dernières années. Un grand nombre d'analyses cladistiques ont examiné les relations de parenté qui existent entre ces dinosaures si bien que la phylogénie de ce clade est actuellement assez bien connue[4],[6],[23],[24],[25],[26],[27],[28],[29].
Ces différentes études et les dernières découvertes ont été synthétisées dans un article de Hendrickx et al. en 2015 proposant une phylogénie complète de ce groupe[30].
Phylogénie des grands groupes de théropodes incluant les genres basaux, d'après Hendrickx et al. (2015):
- Theropoda
- (†Eoraptor)
-
- (†Herrerasauridae)
-
- †Eodromaeus
-
- †Tawa
-
- †Daemonosaurus
- Neotheropoda
- †Coelophysoidea
-
- †Dilophosauridae
- Averostra
- †Ceratosauria
- Tetanurae
- †Megalosauroidea
- Avetheropoda
- †Allosauroidea
- Coelurosauria
- †Tyrannosauroidea
-
- †Compsognathidae
- Maniraptoriformes
- †Ornithomimosauria
- Maniraptora
- †Alvarezsauroidea
-
- †Therizinosauria
- Pennaraptora
- †Oviraptorosauria
- Paraves
- (→Oiseaux)
- Theropoda
- (†Herrerasauridae)
- Neotheropoda
- †Coelophysidae
-
- †Dilophosauridae
- Averostra
- †Ceratosauria
- †Ceratosauridae
- †Abelisauroidea
- †Abelisauridae
- †Noasauridae
- Tetanurae
- †Megalosauroidea
- †Piatnitzkysauridae
- †Megalosauria
- †Megalosauridae
- †Spinosauridae
- Avetheropoda
- †Allosauroidea
- †Metriacanthosauridae
- †Allosauria
- †Allosauridae
- †Carcharodontosauria
- †Carcharodontosauridae
- †Neovenatoridae
- Coelurosauria
- †Tyrannosauroidea
- †Proceratosauridae
- †Tyrannosauridae
-
- †Compsognathidae
- Maniraptoriformes
- †Tyrannosauroidea
- †Allosauroidea
- †Megalosauroidea
- †Ceratosauria
- Maniraptoriformes
- †Ornithomimosauria
- †Deinocheiridae
- †Ornithomimidae
- Maniraptora
- †Alvarezsauroidea
- †Alvarezsauridae
-
- †Therizinosauria
- †Therizinosauridae
- Pennaraptora
- †Oviraptorosauria
- †Caudipteridae
- †Caenagnathoidea
- †Caenagnathidae
- †Oviraptoridae
- Paraves
- †Dromaeosauridae
-
- †Troodontidae
- Avialae → Oiseaux
- †Oviraptorosauria
- †Therizinosauria
- †Alvarezsauroidea
- †Ornithomimosauria
Illustrations
 Acrocanthosaurus
Acrocanthosaurus
 Australovenator
Australovenator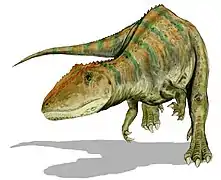 Carcharodontosaurus
Carcharodontosaurus Carnotaurus
Carnotaurus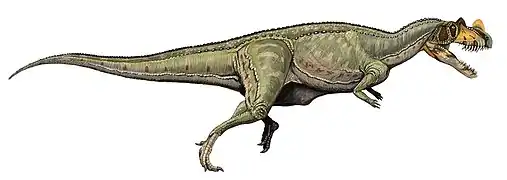 Ceratosaurus
Ceratosaurus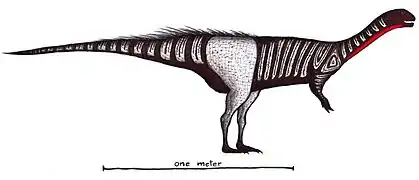 Chilesaurus
Chilesaurus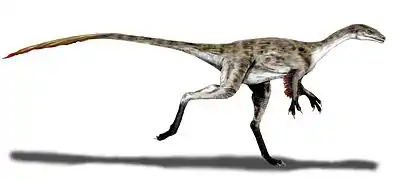 Coelurus
Coelurus Guanlong
Guanlong Concavenator
Concavenator.jpg.webp) Cryolophosaurus
Cryolophosaurus Dilophosaurus
Dilophosaurus Yi
Yi
 Procompsognathus
Procompsognathus Mapusaurus
Mapusaurus
 Torvosaurus
Torvosaurus Nothronychus
Nothronychus Tawa
Tawa.jpg.webp) Yangchuanosaurus
Yangchuanosaurus
Notes et références
Notes
- ↑ Selon une nouvelle classification proposée en 2017, les Théropodes ne sont plus inclus dans l'ordre des Saurischiens mais, avec les Ornithischiens, dans celui des Ornithoscélides[1].
- ↑ Par contre, les études de Zahner et Brinkmann (2019), Poust et al. (2020), Motta et al. (2020), Baiano et al. (2020), confirment les Theropoda inclus dans les Saurischia[2].
Références
- ↑ (en) M.G. Baron, D.B. Norman et P.M. Barrett, « A new hypothesis of dinosaur relationships and early dinosaur evolution », Nature, vol. 543, , p. 501–506 (DOI 10.1038/nature21700).
- ↑ « PBDB », sur paleobiodb.org (consulté le ).
- 1 2 (en) Holtz, T.R. Jr. & Osmólska, H., 2004. Saurischia. Pp. 21-24, in D.B. Weishampel, P. Dodson and H. Osmólska (eds.), The Dinosauria. Second Edition. University of California Press.
- 1 2 (en) Tykoski, R. S. & Rowe, T., 2004. Ceratosauria. Pp. 47–70 in D. B. Weishampel, P. Dodson & H. Osmólska (eds) The Dinosauria, 2nd edn. University of California Press, Berkeley.
- ↑ Zhou, Z.H., 2004. The origin and early evolution of birds: Discoveries, disputes, and perspectives from fossil evidence. Naturwissenschaften 91:455–471.
- 1 2 3 (en) Rauhut, O.W.M., 2003. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontoly 69: 1–213.
- ↑ (en) Paul, G.S., 1988. Predatory Dinosaurs of the World. Simon and Schuster, New York, New York, 464 pp.
- 1 2 (en) Lindsay E. Zanno et Peter J. Makovicky, 2010. "Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution". in Proceedings of the National Academy of Sciences, 2010; DOI: 10.1073/pnas.1011924108.
- ↑ Holtz, T.R. Jr., Chapman, R.E. & Lamanna, M.C., 2004a. Mesozoic Biogeography of Dinosauria. Pp. 627–642 in D.B. Weishampel, Dodson P. & Osmólska H. (eds) The Dinosauria, 2nd edn. University of California Press, Berkeley.
- ↑ (en) Charig, A.J. & Milner, A.C., 1997. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey. Bulletin of the Natural History Museum of London 53: 11-70.
- 1 2 (en) Xu, X., Clark, J.M., Mo, J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C.-K., Han, F.-L. & Guo, Y., 2009. A Jurassic ceratosaur from China helps clarify avian digital homologies. Nature, 459 (18): 940–944.
- ↑ (en) Zanno, L.E., Gillette, D.D., Albright, L.B. & Titus, A.L., 2009. A new North American therizinosaurid and the role of herbivory in 'predatory' dinosaur evolution. Proceedings of the Royal Society B, 276 (1672): 3505-3511.
- ↑ (en) Holtz, T.R. Jr., Brinkman, D.L. & Chandler, C.L., 2000. Denticle morphometrics and a possibly omnivorous feeding habit for the theropod dinosaur Troodon. Gaia 15: 159-166.
- ↑ (en) Mark A. Norell, Peter J. Makovicky et Philip J. Currie, 2001. "The beaks of ostrich dinosaurs". Nature 412: 873-874.
- ↑ Ibrahim, N., Sereno, P. C., Sasso, C. D., Maganuco, S., Fabbri, M., Martill, D. M., Zouhri, S., Myhrvold, N. and Iurino, D. A. 2014. Semiaquatic adaptations in a giant predatory dinosaur. Science 345 (6204): 1613–1616.
- ↑ S. Hartman, « Mass estimates: North vs South redux », Scott Hartman's Skeletal Drawing.com, (consulté le )
- ↑ (en) Zhang, F., Zhou, Z., Xu, X., Wang, X. & Sullivan, C., 2008. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature, 455: 1105-1108.
- ↑ (en) Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. & Guo, Y., 2009. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. Chinese Science Bulletin, 53 (3), 430-435.
- ↑ (en) Xu, X., Cheng, Y.-N. Wang, X.-L. & Chang, C.-H., 2002. An unusual oviraptorosaurian dinosaur from China. Nature, 419: 291-293.
- ↑ (en) Xu, X., Zhou, Z., Wang, X., Kuang, X., Zhang, F. & Du, X., 2003. Four-winged dinosaurs from China. Nature, 421: 335-340.
- ↑ (en) Zhang, F., Zhou, Z., Xu, X. & Wang, X., 2002. A juvenile coelurosaurian theropod from China indicates arboreal habits. Naturwissenschaften, 89 (9): 394-398.
- ↑ BioLib, consulté le 24 mai 2016
- ↑ (en) T. R. Holtz, Jr., R. R. Molnar, et Philip J. Currie, 2004. "Basal Tetanurae". Pp. 71–110 in D. B. Weishampel, P. Dodson & H. Osmólska (éds.) The Dinosauria, 2e éd. University of California Press, Berkeley.
- ↑ R. S. Tykoski, 2005. Anatomy, Ontogeny, and Phylogeny of Coelophysoid Theropods. Unpublished PhD Thesis, The University of Texas at Austin: 1 - 553.
- ↑ (en) Senter, P., 2007. A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology 5 (4): 429-463.
- ↑ (en) N. D. Smith, Peter J. Makovicky, R. Hammer et Philip J. Currie, 2007. "Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution". Zoological Journal of the Linnean Society 151 (2): 377-421
- ↑ (en) S. L. Brusatte et P. Sereno, 2008. "Phylogeny of Allosauroidea (Dinosauria: Theropoda): comparative analysis and resolution". Journal of Systematic Palaeontology, 6: 155-182. Cambridge University Press.
- ↑ (en) M. T. Carrano, et S. D. Sampson, 2008. "The Phylogeny of Ceratosauria (Dinosauria: Theropoda)". Journal of Systematic Palaeontology, 6: 183-236 Cambridge University Press.
- ↑ (en) R. B. J. Benson, M. T. Carrano et S. L. Brusatte, 2009. "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic." Naturwissenschaften, ()
- ↑ (en) C. Hendrickx, S. A. Hartman et O. Mateus, « An Overview of Non- Avian Theropod Discoveries and Classification », PalArch’s Journal of Vertebrate Palaeontology, vol. 12, no 1, , p. 1–73 (lire en ligne).
Voir aussi
Références taxinomiques
- (en) Référence Paleobiology Database : Theropoda Marsh 1881
- (en) Référence Tree of Life Web Project : Theropoda
- (en) Référence BioLib : Theropoda Marsh, 1881 †
- (en) Référence NCBI : Theropoda (taxons inclus)
Articles connexes
- Dinosaure
- Liste de dinosaures
- Merosaurus, un genre informel de dinosaures théropodes du Jurassique inférieur
- Oiseau
Liens externes
- (en) Site de l'université de Berkeley sur les dinosaures.
- (en) Classification détaillée des Theropoda par un spécialiste du genre.
- (it) Blog d'un paléontologue italien spécialiste des théropodes.