الاستماتة أو الموت المبرمج للخلية الحية (Apoptosis، من الإغريقية وتعني "السقوط") هي العملية التي بواسطتها تقتل الخلية الحية نفسها في الكائنات متعددة الخلايا.[2][3][4] تعتبر هذه العملية واحدة من عدة طرق ممكنة تؤدي للموت الخلوي والتي تحدث بتوازن وتزامن مع عملية التكاثر الخلاوي للمحافظة على حياة الكائنات متعددة الخلايا. تتم هذه العملية عن طريق مجموعة من الاحداث البيوكيماوية التي تؤدي لتغيرات في مظهر وبنية الخلايا (مورفولوجيا)، الأمر الذي ينتهي بموت الخلية. وتشمل هذه التغييرات انكماش الخلية:
- تفتيت النووية
- تكاثف الشبكات الكروماتينية
- تفتيت الحمض النووي
- اضمحلال الحمض النووي الرايبوزي (الرنا الرسول).
| استماتة (بيولوجيا) | |
|---|---|
![علاج سرطان البروستات بالاستماتة.[1] رابط لمشاهدة الفيديو](https://upload.wikimedia.org/wikipedia/commons/thumb/2/23/Apoptosis_DU145_cells_mosaic.jpg/450px-Apoptosis_DU145_cells_mosaic.jpg) | |
| معلومات عامة | |
| من أنواع | موت الخلية المبرمج |
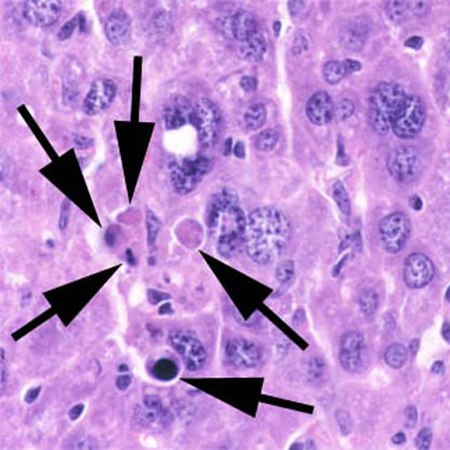
يموت وسطياً ما بين 50 إلى 70 مليار خلية باليوم بسبب الموت الخلوي المبرمج عند الإنسان البالغ. بينما يكون هذا المعدل ما بين 20 مليار إلى 30 مليار خلية يومياً عند الأطفال التي تتراوح أعمارهم ما بين 8 و 14 عام.
وخلافا على ذلك النخر , وهو شكل من موت الخلايا نتيجة الإصابة بالالتهاب الخلوى، و الاستماتة هي عملية منظمة تضفى المزايا أثناء دورة حياة الكائنات الحية.على سبيل المثال، الفصل بين اصابع جنين بشرى في البلدان النامية الآن الخلايا بين الارقام تخضع للاستماتة و خلافا على ذلك النخر، الاستماتة تنتج شظايا خلية تسمى اجسام تشظى حمض د. ن. ا. ووتيرة الزيغ الكروموزومى، و تستطيع هذه الخلايا ابتلاع و ابعاد الخلايا قبل وصولهم لمحتويات الخلية و انتشارهم على الخلايا المحيطة، و أحيانا قد تتسبب في تلف الخلايا المجاورة.
الاستماتة لا يمكن ان تتوقف بمجرد ان بدأت، لانها على درجة عالية من التنظيم.الاستماتة يمكن ان تبدأ عبر أحد المسارات. مسار جوهري يمكن للخلية ان تقتل نفسها، لانه يستشعرهذا المسار بقتل الخلايا لنفسها، و ذلك بسبب بعض الاشارات من الخلايا الاخرى. بعض المسارات التي تحمل موت الخلايا ممكن ان تتنشط من خلال تكسير الانزيمات للبروتين.
البحث عن الطفرات التي تصيب سلسلة الاستماتة زاد بشكل كبيرة منذ اوائل التسعينات، إضافة إلى اهمية ظاهرة بيولوجية قد تورطت مجموعة واسعة من الامراض. الفطريات المفرطة التي تسبب الضمور المفرط، تكون سبب في تكاثر الخلايا غير المنضبط، مثل السرطان. بعض العوامل مثل مستقبلات كاسباس تعمل على تعزيز الفطريات، بينما تمنع بعض أفراد من عائلة bcl-2 من البروتينات حدوث الفطريات.
اكتشافات أصل الكلمة
كان العالم الألماني كارل فوجت أول من وصف مبدأ موت الخلايا المبرمج في 1842. وفي عام 1885، ألقى التشريح فالتر فليمنغ وصف أكثر دقة من عملية موت الخلايا المبرمج. ومع ذلك، لم يكن حتى عام 1965 أن الموضوع هو الأموات. بينما دراسة الانسجة يتم باستخدام المجهر الإلكتروني، وكان جون فوكستون روس كير في جامعة كوينزلاند قادرا على التمييز بين موت الخلايا المبرمج من موت الخلايا الصدمة. في أعقاب نشر ورقة تصف هذه الظاهرة، دعي كير للانضمام اليستر R كوري، وكذلك أندرو وايلي، الذي كان طالب دراسات عليا كوري، وفي جامعة أبردين. في عام 1972، نشر مقال الثلاثي المنوية في المجلة البريطانية للسرطان. وكان كير يستخدم في البداية مصطلح برمجة نخر الخلية، ولكن في هذه المادة، عملية موت الخلايا الطبيعية وتسمى الخلايا. تلقى كير بول إرليخ وجائزة لودفيج في 14 مارس 2000، عن وصفه لموت الخلايا المبرمج. وقال إنه يشارك الجائزة مع بوسطن الأحياء H.
لسنوات عديدة، مصطلح "موت الخلايا المبرمج" لم يكن معروف بشكل كبير، و تحولت الخلايا الميتة من الغموض إلى حقل رئيسي للبحوث وكانوا أمرين: تحديد مكونات سيطرة موت الخلايا وآليات المستجيب، وربط شذوذ في موت الخلايا إلى الأمراض التي تصيب البشر، في سرطان معين.
حصل على جائزة نوبل في الطب عام 2002 لسيدني برينر، هورفيتز وجون إي. وقد تم التعرف على الجينات في الدراسات في الديدان الخيطية C. ايليجانس وهذه الجينات نفسها تعمل في البشر للموت الخلايا المبرمج.
في اليونانية، موت الخلايا المبرمج يترجم إلى "تسقط" من أوراق من شجرة. كورماك، أستاذ اللغة اليونانية، فإن مصطلح للاستخدام الطبي كما كان معنى الطبي بالنسبة لليونانيين أكثر من قبل ألفي سنة. يستخدم أبقراط هذا المصطلح ليعني "التي تقع قبالة العظام". مددت جالينوس معناها إلى "إسقاط الجلبة". وكان كورماك لا شك فيه على بينة من هذا الاستخدام عندما اقترح الاسم.
في كير، ايلي وكوري الورقة الأصلية، وهناك حاشية بشأن النطق:
"نحن في غاية الامتنان للأستاذ جيمس كورماك من قسم اليونانية في جامعة أبردين، لاقتراح هذا المصطلح. إن كلمة" موت الخلايا المبرمج " يستخدم في اليونانية لوصف" انزال "أو" تسقط "من بتلات من الزهور، أو أوراق الشجر. لإظهار الاشتقاق بوضوح، نقترح أن الضغط يجب أن يكون على المقطع ما قبل الأخير، في النصف الثاني من كلمة يجري ضوحا مثل "إطراق" (مع "ع" صامت)، والتي تأتي من نفس الجذر "لتسقط"، ويستخدم بالفعل لوصف تدلى الجفن العلوي ".
آليات التفعيل
يتم تنظيم الشروع في الاستماتة أو موت الخلايا المبرمج بشكل محكم من خلال آليات التفعيل، لأنه حينما تبدأ عملية الاستماتة، فإنها ستؤدي لا محالة إلى موت الخلية. وهناك آليتان للتفعيل هما الأكثر وضوحاً وفهماً: المسار الداخلي (أو مسار الميتوكندريا)، والمسار الخارجي. يتم تفعيل المسار الداخلي عن طريق إشارات بين-خلوية تتولد عندما تكون الخلايا في حالة إجهاد، وتعتمد على إطلاق البروتينات من المساحة بين الغشائية للميتوكندريا. أما المسار الخارجي فيتم تفعيله من روابط خارج الخلية ملتصقة بمستقبلات الموت على سطح الخلية، مما يؤدي إلى تشكل منظومة إشارات تحفيز الموت DISC .
تطلق الخلية إشارات بين-خلوية للاستماتة كرد فعل للتعرض إلى إجهاد، مما قد يؤدي إلى انتحار الخلية. إن ارتباط مستقبلات النواة بالغلوكوكورتيكويدات، الحرارة، إشعاع، الحرمان من التغذية، العدوى الفيروسية، نقص الأكسجة، أو زيادة تركيز الكالسيوم، على سبيل المثال من خلال تضرر الغشاء، قد تؤدي إلى تحفيز إشارات الاستماتة داخل الخلية من قبل خلية تالفة. ويساعد عدد من المركبات الخلوية، مثل متعدد البلمرة الريبوزي ADP، إلى المساعدة في تنظيم عملية الاستماتة.
قبل العملية الفعلية لموت الخلية والتي تتم من خلال الانزيمات، يجب أن تؤدي اشارات الاستماتة إلى دفع البروتينات المنظمة إلى بدء مسار الاستماتة. تتيح هذه الخطوة للإشارات بأن تسبب موت الخلية، أو توقف العملية، إذا لم تعد هناك حاجة لموت الخلية. ترتبط بهذا الأمر العديد من البروتينات، لكن تم تحديد منهجين رئيسيين لتنظيمها: استهداف الميتوكندريا، أو تحويل الإشارة بشكل مباشر من خلال البروتينات المحولة إلى آليات الاستماتة. وتم تحديد المسار الخارجي لبداية العملية في العديد من دراسات المواد السامة، بكونه زيادة في تركيز الكالسيوم داخل الخلية بسبب نشاط مادة دوائية أو مخدرة، مما قد يؤدي أيضاً إلى عملية موت الخلية من خلال ملزمة الكالسيوم البروتيني (كالبين).
المسار الداخلي
إن الميتوكندريا أساسية من أجل الحياة متعددة الخلايا. فبدونها، تتوقف الخلية عن التنفس الهوائي وتموت بسرعة. وتشكل هذه الحقيقة أساساً للعديد من مسارات الاستماتة. تؤثر البروتينات المرتبطة بهذه العملية على الميتوكندريا بطرق متعددة. قد تؤدي إلى تضخم الميتوكندريا من خلال تكوين مسامات في غشائها، أو قد تزيد من نفاذية غشاء المتوكندريا والتسبب بتسرب مؤثرات الاستماتة إلى الخارج. وهي مرتبطة بقوة بالمسار الداخلي، وتظهر الأورام أكثر خلال المسار الداخلي من المسار الخارجي بسبب الحساسية. وهناك كم متزايد من الأدلة التي تشير إلى أن أكسيد النيتريك له القدرة على تحفيز الاستماتة من خلال المساعدة على تبديد مكونات غشاء الميتوكندريا وجعله بالتالي أكثر نفاذية. ووجد أن أكسيد النيتريك مرتبط ببدء أو منع الاستماتة من خلال عمله المحتمل كجزيء إشارة في مسارات لاحقة لتحفيز الاستماتة.
يتم إفراز بروتينات الميتوكندريا المعروفة بـ SMACs إلى العصارة الخلوية للخلية بعد زيادة نفاذية أغشية الميتوكندريا، ترتبط تلك البروتينات بالبروتينات التي تمنع موت الخلية مما يؤدي إلى تثبيطها، مما يسمح بالتالي لعملية الاستماتة بالحدوث. تعمل البروتينات المثبطة لموت الخلية على منع نشاط مجموعة من الانزيمات البروتينية السيستينية تدعى الكاسباس، التي تقوم بعملية تحلل الخلية. ولذلك يمكن اعتبار أن انزيمات التحلل يتم تنظيمها بشكل غير مباشر من خلال نفاذية الميتوكندريا.
يتم إفراز السيتوكروم (سي) أيضاً من الميتوكندريا بسبب تشكل القناة (قناة الاستماتة المحفزة عن طريق الميتوكندريا) في الغشاء الخارجي للميتوكندريا، وله وظيفة تنظيمية حيث يسبق التغير المورفولوجي المرتبط بالاستماتة. وحين يتم افراز السيتوكروم (سي) يرتبط بعامل تنشيط الانزيم البروتيني للاستماتة (Apaf) وجزيئات ATP، التي ترتبط بدورها بمحفز الكاسباس-9 لتكوين مركب بروتيني يعرف باسم apoptosome، مشكلا الكاسباس-9 الذي ينشط الكاسباس-3. ويتم تنظيم القناة من خلال العديد من البروتينات.
المسار الخارجي
تم اقتراح نظريتين حول البدء المباشر لآليات الاستماتة في الثدييات: TNF أي نموذج عامل النخر الورمي، وFAS وهو نموذج يتم بواسطة جزيئات FAS. ويتعلق النموذجان بمستقبلات TNF المرتبطة بإشارات خارجية.
مسار TNF:
إن ألفا-TNF هو سايتوكين يتم إنتاجه من قبل الخلايا البلعمية، وهو الوسيط الخارجي الرئيسي للاستماتة. معظم الخلايا في الجسم البشري لديها مستقبلين للألفا-TNF : TNF-1 و TNF-2. وجد أن ارتباط الألفا-TNF بالمستقبل الأول يؤدي إلى إطلاق مسار تنشيط الكاسباس من خلال البروتينات الغشائية الوسيطة المرتبطة بنطاق الإماتة (TRADD) و(FADD). يؤدي الارتباط بهذا المستقبل بشكل غير مباشر إلى تفعيل عوامل النسخ المتعلقة ببقاء الخلية وردود الفعل الالتهابية. لكن إطلاق الاشارات من خلال هذا المستقبل قد يؤدي إلى تفعيل الاستماتة على نحو مستقل عن الكاسباس. وتظهر الصلة بين ألفا-TNF والاستماتة سبب كون الإنتاج غير الطبيعي من الالفا-TNF يلعب دورا في العديد من الامراض البشرية، خاصة أمراض المناعة الذاتية.
مسار FAS :
يرتبط مستقبل فاس (أول إشارة للاستماتة) ويعرف أيضا بـ Apo-1 أو CD95، ببروتين FasL، وهو بروتين عابر للأغشية من عائلة TNF. يؤدي التفاعل بين الاثنين إلى تكوين منظومة إشارات تحفيز الموت، المحتوية على كاسباس-8 وكاسباس-10 والفاد (FADD). وهذه المكونات تطلق بشكل مباشر كاسباسات أخرى تؤدي إلى تنفيذ عملية الاستماتة في بعض انواع الخلايا. وفي بعض انواع الخلايا الاخرى تؤدي هذه المكونات إلى اطلاق حلقة من العوامل المحفزة للاستماتة من الميتوكندريا وزيادة تنشيط كاسباس-8.
مكونات مشتركة:
بعد تفعيل المسارات السابقة في خلايا الثدييات يتم خلق توازن بين المكونات المحفزة للاستماتة وتلك المثبطة لها. وهذا التوازن ينجم عن كمية مركبات محفزة للاستماتة في الغشاء الخارجي للميتوكندريا. وهذه المركبات لازمة لجعل الغشاء نفاذا لاطلاق محفزات الكاسباس مثل السيتوكروم (سي) وSMAC.
الكاسباس:
تلعب الكاسباس دورا اساسيا في نقل اشارات الاستماتة، والكاسباس هي بروتينات انزيمية سيستينية محددة الاسبارتات. وهناك نوعان من الكاسباس: الكاسباس البادئ (2-8-9-10-11-12) والكاسباس المستجيب (3-6-7).
وهناك أيضاً مسارات استماتة مستقلة عن الكاسباس تتم عن طريق عوامل منشطة للاستماتة AIF.
نموذج الاستماتة في البرمائيات
يشكل الضفدع البرمائي Xenopus laevis نموذجاً مثالياً لدراسة آليات الاستماتة. في الواقع، يحفز اليود والثيروكسين أيضاً عمليات الاستماتة المدهشة في خلايا خياشيم وذيل وزعانف اليرقة خلال عملية تحول البرمائي، وتعمل على تحفيز تطور الجهاز العصبي ليتحول الشرغوف النباتي المائي إلى ضفدع أرضي آكل للحم.
قتل الخلية
العديد من المسارات والإشارات تؤدي إلى موت الخلايا المبرمج (الاستماتة)، ولكن هذه تتقارب على آلية واحدة تسبب فعلا موت الخلية. بعد أن تتلقى الخلية تحفيزا، فإن العضيات الخلوية تخضع للتدهور عن طريق نشاط البروتين caspases . بالإضافة إلى تدمير العضيات الخلوية، إن الحمض النوي الرايبوزي الرسول يتدهور بسرعة عن طريق آلية التي لم تتميز تماما بعد.يتم تشغيل اضمحلال الحمض النوي الرايبوزي الرسول في وقت مبكر جدا في الاستماتة. وتظهر الخلايا التي تمر في موت الخلايا المبرمج (الاستماتة) بأشكال متعددة: 1- انكماش الخلايا والتقريب بسبب انهيار الهيكل الخلوي البروتيني عن طريق ال caspases
2- يظهر السيتوبلازم بشكل كثيف، والعضيات تظهر معبأة بإحكام.
3- الكروماتين يخضع للتكثيف إلى بقع مدمجة ضد الغلاف النووي (المعروف أيضا باسم المغلف حول النواة) في عملية تعرف باسم التغلظ، وهي السمة المميزة لموت الخلايا المبرمج.
4- المغلف النووي يصبح متقطع والحمض النووي داخله موزعة في عملية يشار إلى تمزق النواة. النواة تكسر إلى العديد من الاجسام الكروماتينية أو وحدات أنوية صغيرة وذلك بسبب تدهور الحمض النووي.
5- يظهر غشاء الخلية على شكل براعم غير منتظمة والمعروفة باسم الفقاعات.
6- الخلية تنهار إلى حويصلات متعددة تسمى اجسام الاستماتة، والتي يحدث لها فيما بعد البلعمة. موت الخلايا المبرمج يتقدم بسرعة ويتم إزالة مننجاتها بسرعة، مما يجعل من الصعب اكتشافها أو تصورها. خلال تمزق النواة، وتفعيل داخل النواة يترك اجزاء من الحمض النووي قصيرة، ومتباعدة بصورة منتظمة في الحجم. هذه تعطي سمة 'laddered' والتي تظهر على صحن بتري الجل بعد تعريضها لتيار كهربائي. اختبارات الحمض النووي laddering يفرق موت الخلايا المبرمج من نقص تروية أو موت الخلايا السامة.
التخلص من الخلايا الميتة
وقد أطلق على إزالة الخلايا الميتة من الخلايا البلعمية المجاورة ب دفن الخلايا الميتة موت الخلايا التي تخضع للمراحل النهائية من الاستماتة عرض الجزيئات بلعمية، مثل فسفاتيديل، على سطح الخلية الخاصة بهم. تم العثور على فسفاتيديل عادة على سطح الخلية الداخلي لغشاء البلازما، ولكن يتم إعادة توزيعه خلال موت الخلايا المبرمج على سطح الخلية من بروتين يعرف باسم scramblase. هذه الجزيئات تمنح الخلية علامة لعملية البلعمة من خلال الخلايا التي تمتلك المستقبلات المناسبة، مثل الخلاية المسؤولة عن البلعمة. إزالة الخلايا الميتة من قبل البالعات يحدث بشكل منظم دون استثارة واستجابة التهابية.
Pathway knock-outs
تم إجراء العديد من طرق الرافضة في مسارات موت الخلايا المبرمج لاختبار وظيفة كل من البروتينات. العديد من caspases، بالإضافة إلى APAF-1 وFADD، وقد تحور لتحديد النمط الظاهري الجديد. من أجل إنشاء عامل نخر الورم (TNF)، تمت إزالة exon تحتوي على النيوكليوتيدات 3704-5364 من الجين. هذا exon يقوم بترميز جزء من TNF الناضجة، وكذلك التسلسل الرئيسي، وهو منطقة محفوظة للغاية اللازمة لمعالجة الخلايا. TNF - / - الفئران تتطور بشكل طبيعي وليس لديهم تشوهات هيكلية أو المورفولوجية الجسيمة. ومع ذلك، عند التحصين مع SRBC ( خلايا الدم الحمراء للأغنام)، وأظهرت هذه الفئران وجود نقص في نضوج استجابة الأجسام المضادة. كانت قادرة على توليد مستويات طبيعية من الغلوبولين المناعي، ولكن لا يمكن أن تتطور لمستويات محددة. Apaf-1 هو البروتين الذي يتحول على caspase 9 من الانقسام لبدء سلسلة caspase الذي يؤدي إلى موت الخلايا المبرمج. منذ - / - طفرة في الجين APAF-1 هي الجنينية القاتلة، كان يستخدم استراتيجية فخ الجينات من أجل توليد APAF-1 - / -الفأر. يستخدم هذا الاختبار لتعطيل وظيفة الجين عن طريق إنشاء الانصهار الجين داخل الجين الاخر. عند تقديم اعتراض الجينات APAF-1 في الخلايا، تحدث العديد من التغيرات المورفولوجية، مثل السنسنة المشقوقة، واستمرار شبكات بين الأصابع، وفتح الدماغ. وبالإضافة إلى ذلك، بعد اليوم الجنينية 12.5، أظهر دماغ الأجنة العديد من التغييرات الهيكلية. محمية APAF-1 الخلايا من المحفزات الخلايا مثل الإشعاع. وBAX-1 المعارض الفأر وتشكيل الدماغ الأمامي العادي وانخفضت إلى موت الخلايا المبرمج في بعض السكان العصبية وفي الحبل الشوكي، مما يؤدي إلى زيادة في الخلايا العصبية الحركية.
البروتينات caspase هي جزء لا يتجزأ من مسار موت الخلايا المبرمج، لذلك يترتب على knock-outs المصنوعة وقد تحتوي على نتائج ضارة ومتنوعة.A caspase 9 knock-out يؤدي إلى تشوه شديد في الدماغ. ، caspase 8 knock-out يؤدي إلى فشل القلب والفتك. ومع ذلك، مع استخدام التكنولوجيا، تم إنشاء caspase 8 knock-out يؤدي إلى زيادة في خلايا T الطرفية، استجابة خلايا T الضعيفة، وخلل في إغلاق الأنبوب العصبي. تم العثور على هذه الفئران لتكون مقاومة لموت الخلايا المبرمج بوساطة CD95، TNFR، وما إلى ذلك ولكن ليست مقاومة لموت الخلايا المبرمج التي تسببها الأشعة فوق البنفسجية، وأدوية العلاج الكيميائي، وغيرها من المحفزات. وأخيرا، caspase 3 knock-out تميزت على الخلايا خارج الرحم في ملامح الدماغ والاستماتة الغير طبيعية مثل غشاء ال blebbing أو تجزئة النووي. وهناك ميزة رائعة من هذه KO الفئران هو أن لديهم النمط الظاهري محدودة جدا: Casp3، 9، وقد أظهرت الفئران APAF-1 KO تشوهات الأنسجة العصبية وFADD وCASP 8 KO تطور القلب ، ولكن في كلا النوعين من KO غيرها من الأجهزة المتقدمة عادة، وكانت لا تزال بعض أنواع الخلايا الحساسة من الاستماتة مما يشير إلى أن مسارات ما قبل الاستماتة غير معروفة.
طرق للتمييز بين الاستماتة والنخر
من أجل أداء تحليل الاستماتة مقابل النخر، يمكن للمرء القيام به بتحليل الشكل لكل منه بواسطة المجهر، وقياس التدفق الخلوي، و المجهر الإلكتروني النافذ. هناك أيضا العديد من التقنيات الحيوية لتحليل علامات سطح الخلية (تعرض فسفاتيديل مقابل نفاذية الخلية من خلال قياس التدفق الخلوي)، علامات خلوية مثل تجزئة الحمض النووي (قياس التدفق الخلوي)، وتفعيل caspase، محاولة انشقاق، وتحرير السيتوكروم ج. من المهم أن تعرف كيف الابتدائية والثانوية من خلايا النخرالتي يمكن تمييزها من خلال تحليل supernatant.
HMGB1 والإفراج عن cytokeratin 18. ومع ذلك، فقد لم تم تحديد أي سطح واضح أو علامات حيوية في موت الخلايا النخرية بعد، وتتوفر فقط علامات سلبية. وتشمل هذه عدم وجود علامات للاستماتة (تفعيل caspase، وتحرير السيتوكروم ج، وتفتيت الحمض النووي وطاقة حركية مشتقة من علامات موت الخلايا (تعريض فسفاتيديل ونفاذية غشاء الخلية. مجموعة مختارة من التقنيات التي يمكن استخدامها لتمييز خلايا الاستماتة من خلايا النخريمكن العثور عليها في هذه المراجع.
المشاركة في المرض
المسارات الناقصة (غير المكتملة)
أنواع كثيرة مختلفة من مسارات الاستماتة تحتوي على العديد من المكونات الكيميائية الحيوية المختلفة، وكثير منهم لم بتم فهمه حتى الان. كمسار هو أكثر أو أقل متسلسل في الطبيعة، وإزالة أو تعديل عنصر واحد يؤدي إلى التأثير في بلد آخر. في الكائن الحي، وهذا يمكن أن يكون له آثار وخيمة، وغالبا في شكل مرض أو اضطراب. و من شأن مناقشة كل الأمراض الناجمة عن تعديل مختلف لمسارات الاستمانة لبكون غير عملي، ولكن هذا المفهوم بغمر كل واحد هو نفسه: لقد تم تعطل السير العادي للمسار في مثل هذه الطريقة لتنال من قدرة الخلية على الخضوع ل الخلايا العادية. هذه النتائج في الخلية التي تعيش ماضيها "استخدام من قبل التاريخ"، وغير قادرة على تكرار وتمر على أي آلية خاطئة لنسله، مما يزيد من احتمال الخلية تصبح سرطانية أو المريضة.
و مؤخرا تم وصف مثال لهذا المفهوم في العمل يمكن أن ينظر في تطوير سرطان الرئة يسمى NCI-H460. المانع العاشر مرتبطة من البروتين الاستماتة (XIAP) هي overexpressed في الخلايا من خط الخلية H460 . XIAPs يربط النموذج من كاسباس 9، و يمنع انشطة الفطريات لمنشط السيتوكروم ج، وبالتالي overexpression يؤدي إلى انخفاض في كمية منبهات proapoptotic. ونتيجة لذلك، ويخل بالتوازن من المؤثرات مكافحة للفطريات وproapoptotic في صالح الأول، وتستمر الخلايا التالفة لتكرار رغم أنها موجهة للموت.
ديسريغولاتيون من البروتين p53
يتراكم البروتين p53 عند تلف الحمض النووي نتيجة لسلسلة من العوامل البيوكيميائية. ويشمل جزء من هذا المسار ألفا انترفيرون بيتا وانترفيرون، مما يحفز نسخ من الجين p53، مما يؤدي إلى زيادة مستوى البروتين p53 وتعزيز سرطان الخلايا المبرمج. البروتين p53 يمنع الخلايا من التكاثر عن طريق إيقاف دورة الخلية في G1، أو الطور البيني، لإتاحة الوقت خلية للإصلاح، ومع ذلك فإنه سوف يحفز الخلايا إذا كان الضرر واسع النطاق وتفشل جهود الإصلاح. فإن أي اختلال في تنظيم الجينات البروتين p53 أو فيروسات تؤدي إلى موت الخلايا المبرمج ضعف واحتمال تكوين الأورام.
الكبت (المنع)
تثبيط الخلايا يمكن أن يؤدي إلى عدد من أنواع السرطان، وأمراض المناعة الذاتية، والأمراض الالتهابية، والعدوى الفيروسية. وكان يعتقد في البداية أن تراكم الخلايا المرتبط يرجع إلى زيادة في انتشار الخليوي، ولكن من المعروف الآن أنه يرجع أيضا إلى انخفاض في موت الخلايا. الأكثر شيوعا من هذه الأمراض هو السرطان، ومرض الانتشار الخلوي المفرط، والتي غالبا ما تتميز overexpression من أفراد الأسرة IAP. ونتيجة لذلك، فإن الخلايا الخبيثة تجرب استجابة غير طبيعية لتحريض موت الخلايا المبرمج: الجينات التي تنظم دورة (مثل البروتين p53، رأس أو ج-MYC) وتحور أو المعطل في الخلايا المريضة، وجينات أخرى (مثل بي سي إل-2) تعديل أيضا التعبير عنها في الأورام.
الخلية هيلا
موت الخلايا المبرمج في هيلا هو تحول خلايا البروتينات التي تنتجها الخلية؛ هذه البروتينات المثبطة تستهدف شبكية البروتينات قمع الورم. هذه بروتينات قمع ورم تنظم دورة الخلية، ولكن بإبطال عندما منضم إلى البروتين المثبط. فيروس الورم الحليمي البشري E6 و E7 هي بروتينات المثبطة التي أعرب عنها فيروس الورم الحليمي البشري، وفيروس الورم الحليمي البشري هي المسؤولة عن تشكيل ورم عنق الرحم التي تشتق خلايا هيلا. فيروس الورم الحليمي البشري E6 يسبب البروتين p53، الذي ينظم دورة الخلية، لتصبح غير نشطة. فيروس الورم الحليمي البشري E7 ترتبط بالبروتين قمع الشبكية الورم ويحد من قدرتها على السيطرة على انقسام الخلايا. هذه اثنين من البروتينات المثبطة هي جزئيا مسؤولة عن الخلود خلايا هيلا "عن طريق تثبيط الخلايا أن تحدث. CDV (سل الكلاب الفيروسات) قادر على إحداث موت الخلايا المبرمج على الرغم من وجود هذه البروتينات المثبطة. هذا هي خاصية حال الورم مهمة من CDV: هذا الفيروس قادر على قتل خلايا سرطان الغدد الليمفاوية الكلاب. Oncoproteins E6 و E7 لا تزال تترك البروتين p53 خاملة، ولكنها ليست قادرة على تجنب تفعيل caspases الناجم عن ضغوط عدوى فيروسية. قدمت هذه الخصائص حال الورم وجود صلة واعد بين CDV وموت الخلايا المبرمج سرطان الغدد الليمفاوية، والتي يمكن أن تؤدي إلى تطوير طرق العلاج البديلة لكل من سرطان الغدد الليمفاوية الكلاب وسرطان الغدد الليمفاوية غير هودجكين البشرية. ويعتقد أن العيوب في دورة الخلية لتكون مسؤولة عن مقاومة للعلاج الكيميائي أو الإشعاع عن طريق الخلايا السرطانية معينة، لذلك الفيروس الذي يمكن أن يحفز الخلايا على الرغم من وجود عيوب في دورة الخلية هو مفيد لعلاج السرطان.
العلاجات
الطريقة الرئيسية لعلاج الموت من الأمراض المرتبطة يشير على زيادة أو تقليل الحساسية من موت الخلايا المبرمج في الخلايا المريضة، وهذا يتوقف على ما إذا كان المرض سببه إما تثبيط أو الخلايا الزائدة. على سبيل المثال، وتهدف علاجات لاستعادة الخلايا لعلاج الأمراض مع موت الخلايا ناقصة، وزيادة عتبة الفطريات لعلاج الأمراض تتعامل مع موت الخلايا المفرط. لتحفيز الخلايا، يمكن للمرء زيادة عدد بروابط مستقبلات الموت (مثل TNF أو TRAIL)، استعداء مكافحة الفطريات بي سي إل 2 ممرا أو إدخال محاكيات بالألغام لمنع المانع (خطط العمل الفردية). إضافة عوامل مثل هيرسيبتين، اريسا، أو جليفيك تعمل على وقف الخلايا من ركوب الدراجات ويؤدي تنشيط الخلايا من خلال منع النمو والبقاء على قيد الحياة مما يشير إلى مزيد من المنبع. وأخيرا، مضيفا المجمعات البروتين p53-MDM2 يزيح البروتين p53 ويقوم بتشغيل الممر البروتين p53، مما أدى إلى اعتقال دورة الخلية وموت الخلايا المبرمج. العديد من الأساليب المختلفة التي يمكن أن تستخدم إما لتحفيز أو تثبيط الخلايا في أماكن مختلفة على طول الطريق مما يشير إلى الموت.
موت الخلايا المبرمج هو متعددة الخطوات، متعدد مسار برنامج الخلية الموت التي هي متأصلة في كل خلية من خلايا الجسم. في السرطان، ويتم تبديل نسبة الخلايا في تقسيم الخلايا. علاج السرطان بواسطة العلاج الكيميائي والإشعاع يقتل الخلايا المستهدفة في المقام الأول عن طريق حفز الخلايا.
النشاط المفرط في عملية الاستماتة
من ناحية أخرى، فقدان السيطرة على موت الخلايا (مما يؤدي إلى زيادة في موت الخلايا المبرمج (الاستماتة) ) يمكن أن يؤدي إلى أمراض الاعصاب والأمراض الدموية، وتلف الأنسجة. تطور فيروس نقص المناعة البشرية يرتبط ارتباطا مباشرا ، موت الخلايا المبرمج الغير المنظم والمفرط به . في الشخص السليم، عدد من CD4 + الخلايا الليمفاوية هي في توازن مع الخلايا التي تم إنشاؤها بواسطة نخاع العظام. ومع ذلك، في المرضى المصابين بفيروس نقص المناعة البشرية، يتم فقدان هذا التوازن بسبب عدم قدرة نخاع العظم على تجديد خلايا CD4 +. في حالة فيروس نقص المناعة البشرية، اللمفاويات CD4 + يموتون بمعدل متسارع من خلال الخلايا غير المتحكم فيها، عندما يتحفز.
العلاجات
العلاجات التي تهدف إلى منع عمل caspases محددة. وأخيرا، فإن البروتين كيناز AKT يعزز بقاء الخلية من خلال مسارين. AKT phosphorylates ويمنع Bas (أحد أفراد عائلة Bcl-2)، مما Bas يتسبب للتفاعل مع سقالة 14-3-3، مما يؤدي إلى تفكك Bcl، وبالتالي بقاء الخلية حية . AKT أيضا ينشط IKKα، الأمر الذي يؤدي إلى تفعيل NF-kB وبقاء الخلية حية . تنشيط NF-kB يحث على تفعيل الجينات المضادة مثل Bcl-2 ينتج في تهبيط مفعول الاستماتة ، . وقد وجد أن NF-kB يلعب على حد سواء دورا في مقاومة للاستماتة ودور في الاستماتة الاولية اعتمادا على المحفزات المستخدمة ونوع من الخلايا.
تطور فيروس نقص المناعة البشرية
هو التقدم من عدوى فيروس نقص المناعة البشرية إلى الإيدز يرجع في المقام الأول إلى استنزاف CD4 + الخلايا الليمفاوية التائية T المساعدة بطريقة سريعة جدا لنخاع العظام في الجسم لتجديد الخلايا، مما يؤدي إلى الجهاز المناعي للخطر. واحدة من الآليات التي تنضب خلايا T المساعدة هو موت الخلايا المبرمج (الاستماتة)، الذي ينتج عن سلسلة من المسارات البيوكيميائية:
1- إنزيمات فيروس نقص المناعة البشرية تثبط عمل مكافحة الاستماتة Bcl-2. هذا لا يسبب موت الخلايا مباشرة ولكن ينبغي أن تلقى إشارة مناسبة إلى الخلية لموت الخلايا المبرمج. في موازاة ذلك، هذه الإنزيمات تنشط proapoptotic procaspase-8، والتي تقوم بتفعيل مباشرة أحداث الميتوكوندريا من موت الخلايا المبرمج (الاستماتة) .
2- فيروس نقص المناعة البشرية يمكن أن يزيد من مستوى البروتينات الخلوية التي تسرع موت الخلايا المبرمج (الاستماتة)
3- بروتينات فيروس نقص المناعة البشرية تقلل من كمية CD4 بروتين سكري في علامة موجودة على غشاء الخلية.
4- تحرر الجسيمات الفيروسية والبروتينات الموجودة في السائل خارج الخلية قادرة على إحداث موت الخلايا المبرمج (الاستماتة ) في "المتفرج" الخلايا التائية T المساعدة المجاورة.
5- يقلل فيروس نقص المناعة البشرية انتاج جزيئات تشارك في ترميز الخلية من اجل الاستماتة، وإعطاء الوقت للفيروس إلى تكرار ومواصلة الإفراج والتحرير عن عوامل الاستماتة والفيرونات في الأنسجة المحيطة بها.
6- قد تتلقى خلايا CD4 + المصابة أيضا إشارة الموت من خلية تائية سامة
الخلايا أيضا قد تموت لنتيجة مباشرة من العدوى الفيروسية. HIV-1 الحث الأنبوبي للخلية G2 / M الاعتقال والاستماتة الانتقال من فيروس نقص المناعة البشرية الإيدز ليست فورية أو حتى بالضرورة سريعة. يتم تصنيف نشاط السمية لخلايا فيروس نقص المناعة البشرية تجاه CD4 + الخلايا الليمفاوية وباء الإيدز مرة واحدة CD4 + عدد خلايا المريض تيهبط دون 200.
عدوى فيروسية
يحدث التحريض الفيروسي من الاستماتة عندما يصاب واحد أو عدة خلايا من الكائن الحي مع الفيروس، مما يؤدي إلى موت الخلايا. موت الخلايا في الكائنات الحية ضروري للتطور الطبيعي للخلايا ونضوج دورة الخلية. ومن المهم في الحفاظ على وظائف وأنشطة العادية للخلايا أيضا.
يمكن للفيروسات أن تؤدي إلى موت الخلايا المبرمج (الاستماتة) للخلايا المصابة عبر مجموعة من الآليات بما في ذلك:
1- منطقة ربط المستقبلات .
2- تفعيل بروتين كيناز (R ( PKR .
3- التفاعل مع بروتين p53
4- التعبيرعن البروتينات الفيروسية بالإضافة إلى البروتينات MHC على سطح الخلية المصابة، مما يسمح لخلايا الجهاز المناعي (مثل الخلايا القاتلة الطبيعية والسامة للخلايا T) التعرف عليها، ثم تقوم بتحفيز الخلية المصابة إلى الخضوع للاستماتة
ومن المعروف فيروس حمى الكلاب (CDV) أن يسبب بموت الخلايا المبرمج (الاستماتة) في الجهاز العصبي المركزي والأنسجة اللمفاوية من الكلاب المصابة في الجسم الحي في المختبر. موت الخلايا المبرمج الناجمة عن CDV عادة ما يسببها عبر المسار الخارجي، الذي ينشط caspases التي تعطل وظيفة الخلايا، ويؤدي في نهاية المطاف إلى موت الخلايا. في الخلايا الطبيعية، CDV ينشط أولا caspase-8 ، والذي يعمل البروتين البادئ تليها caspase-3 البروتين الجلاد. ومع ذلك، موت الخلايا المبرمج الناجم عن CDV في خلايا هيلا لا تنطوي على البروتين البادئ caspase-8. هيلا موت الخلايا المبرمج الناجمة عن CDV يتبع آلية مختلفة من ذلك في خطوط الخلايا فيرو. هذا التغيير في caspase تتالي تشير CDV يدفع الخلايا عبر المسار الجوهري، باستثناء الحاجة إلى البادئ caspase-8. بدلا من ذلك تنشيط بروتين الجلاد من المحفزات الداخلية التي تسببها عدوى فيروسية لا تتاليcaspase cascade
تم العثور على فيروس أوروبوش (OROV) في عائلة الفيروسات البنياوية. وقد بدأت الدراسة على موت الخلايا المبرمج الناجمة عن الفيروسات البنياوية في عام 1996، عندما لوحظ أن موت الخلايا المبرمج والناجم عن الفيروس لاكروس في خلايا الكلى من الهامستر الطفل وفي أدمغة اطفال الفئران .
OROV هو المرض الذي ينتقل بين البشر من قبل ذبابة العض (البعضوضيات paraensis). يشار إليها باعتبارها الفيروسة المنقولة بالمفصليات الحيوانية ويسبب مرض الحمى، والتي تتميز بداية من حمى مفاجئة تعرف باسم حمى أوروبوش.
يسبب الفيروس أوروبوش أيضا اضطراب في الخلايا المستزرعة - الخلايا التي تزرع في ظروف متميزة ومحددة. مثال على ذلك يمكن النظر في خلايا هيلا، حيث تبدأ الخلايا لتتحول بعد فترة وجيزة من الإصابة.
مع استخدام الهلام الكهربائي، فإنه يمكن ملاحظة أن OROV يسبب تجزئة الحمض النووي في خلايا هيلا. ويمكن أن تفسر عن طريق عد وقياس وتحليل الخلايا من موع خلية فرعية / G1 عندما يصاب خلايا هيلا مع OROV، يتم تحرير السيتوكروم C من غشاء الميتوكوندريا، في العصارة الخلوية للخلايا. هذا النوع من التفاعل يدل على أن موت الخلايا المبرمج هو تفعيلها عن طريق المسار الجوهري.
من أجل موت الخلايا المبرمج تحدث داخل OROV، الفيروس الغير مغطى ، والاستيعاب الفيروسي، من الضروري جنبا إلى جنب مع تكرار الخلايا . يتم تنشيط الخلايا في بعض الفيروسات من خلال المحفزات خارج الخلية. ومع ذلك، فقد أثبتت الدراسات أن العدوى OROV يسبب موت الخلايا المبرمج ليتم تفعيلها من خلال محفزات الخلايا، وينطوي على الميتوكوندريا.
العديد من الفيروسات تشفريربروتينات التي يمكن أن تحول دون موت الخلايا المبرمج. عدة فيروسات تشفر فيروسات متشابهة من Bcl-2 . ويمكن لهذا التشابه أن يمنع بروتينات الاستماتة مثل BAX وBAK ، التي تعتبر ضرورية لتفعيل موت الخلايا المبرمج. وتشمل الأمثلة على الفيروسية Bcl-2 بروتينات ابشتاين بار بروتين BHRF1 الفيروس واتش البروتين E1B 19K بعض الفيروسات تعبر عن مثبطات caspase التي تمنع النشاط ل caspase ومثال على ذلك هو بروتين CrmA من الفيروسات جدري البقر. بينما عدد من الفيروسات يمكن أن تمنع آثار TNF وFas. على سبيل المثال، البروتين M-T2 من الفيروسات المخاطي يمكن ربط TNF منعه من ربط مستقبلات TNF وتحفزه على الاستجابة لها. وعلاوة على ذلك، العديد من الفيروسات تعبر عن البروتين p53 مثبطات التي يمكن أن تربط البروتين p53 وتمنع النشاط النسخي transactivation لها. ونتيجة لذلك، البروتين p53 لا يمكن أن يحفز الخلايا، لأنه لا يمكن أن يحدث في التعبير عن بروتينات الاستماتة . واتش البروتين E1B-55K وفيروس التهاب الكبد B بروتين HBX أمثلة من البروتينات الفيروسية التي يمكن أن تؤدي هذه الوظيفة.
يمكن للفيروسات أن تظل سليمة من الخلايا وخاصة في المراحل الأخيرة من العدوى. يمكن تصديرها في أسام الاستماتة التي انفصلت من سطح الخلية الميتة ، وحقيقة أنهم ابتلعوعن طريق البلعمة يمنع الشروع في استجابة المضيف. هذا التفضل في انتشار الفيروس.
النباتات
موت الخلية المبرمج في النباتات لديها عدد من أوجه التشابه الجزيئية إلى أن موت الخلايا المبرمج في الحيوان، ولكن لديها أيضا اختلافات، أبرز تلك الفصائل وجود جدار الخلية وعدم وجود نظام المناعة التي تزيل قطعة من الخلايا الميتة. بدلا من الاستجابة المناعية، الخلية الميتة تجمع المواد لتكسير و تحطيم نفسها باستمرار ثم توضع في فجوة كما عملية موت الخلية. إذا كانت هذه العملية برمتها تشبه الخلايا الحيوانية بشكل وثيق بما يكفي لتبرير استخدام الخلايا اسم (على عكس أعم موت الخلية المبرمج) غير واضح.
استقلال عملية الاستماتة
مصطلح ال caspases يسمح في تطوير المثبطات، والتي يمكن أن تستخدم لتحديد ما إذا كانت عملية الخلوية تتضمن caspases النشطة. باستخدام هذه المثبطات اكتشف أن الخلايا يمكن أن تموت أثناء عرض التشكل مماثلة لموت الخلايا المبرمج دون تفعيل كاسباس. دراسات لاحقة ترتبط هذه الظاهرة إلى الإفراج عن الوكالة الدولية للفرنكوفونية (عامل يحفز موت الخلايا المبرمج) من الميتوكوندريا والنبات في داخل النواة بوساطة NLS لها (إشارة توطين النووية). داخل الميتوكوندريا، يرتكز AIF إلى الغشاء الداخلي. من أجل أن يطلق سراحه، هو المشقوق البروتين من قبل calpain البروتيني التي تعتمد على الكالسيوم.
تنبؤ بروتينات الخلايا في المواقع التحت الخلوية
في عام 2003، تم تطوير طريقة للتنبؤ مواقع التحت الخلوية من البروتينات موت الخلايا المبرمج. وبعد ذلك، تم تطوير مختلف وسائل لتكوين الأحماض الأمينية لتحسين نوعية التنبؤ لتوطين المواقع التحت الخلوية من البروتينات موت الخلايا المبرمج على أساس تسلسل المعلومات وحدها.
انظر أيضاً
- نخر
- سرطان
- تحلل ذاتي
- موت الخلية المبرمج
- بروتين موت الخلية المبرمج 1
- موت الخلايا
- قائمة الجينات البشرية
المراجع
- "Quantitative phase contrast microscopy — label-free live cell imaging and quantification". Phase Holographic Imaging AB. مؤرشف من الأصل في 14 فبراير 2019.
- Cotran, Ramzi, S.; Kumar, Collins (1998). Robbins Pathologic Basis of Disease. Philadelphia: W.B Saunders Company. .
- American Heritage Dictionaryنسخة محفوظة June 30, 2008, على موقع واي باك مشين. نسخة محفوظة 30 يونيو 2008 على موقع واي باك مشين.
- Nagata S (April 2000). "Apoptotic DNA fragmentation". Exp. Cell Res. 256 (1): 12–8. doi:10.1006/excr.2000.4834. PMID 10739646.