| Histamine | ||
| ||
 | ||
| Identification | ||
|---|---|---|
| Nom UICPA | 2-(3H-imidazol-4-yl)éthanamine | |
| No CAS | ||
| No ECHA | 100.000.092 | |
| No CE | 200-100-6 | |
| Code ATC | L03 V04 |
|
| PubChem | 774 | |
| SMILES | ||
| InChI | ||
| Apparence | Cristaux incolores | |
| Propriétés chimiques | ||
| Formule | C5H9N3 [Isomères] |
|
| Masse molaire[1] | 111,145 1 ± 0,005 2 g/mol C 54,03 %, H 8,16 %, N 37,81 %, |
|
| pKa | 9,8 (25 °C)[2] | |
| Propriétés physiques | ||
| T° fusion | 86 °C[2] | |
| T° ébullition | 209,5 °C | |
| Solubilité | soluble dans l'eau et l'éthanol, insoluble dans l'éther |
|
| Précautions | ||
| Directive 67/548/EEC | ||
 Xn |
||
| Écotoxicologie | ||
| DL50 | 220 mg·kg-1 (souris, oral) 385 mg·kg-1 (souris, i.v.) 2 500 mg·kg-1 (souris, s.c.) 725 mg·kg-1 (souris, i.p.) [2] |
|
| Unités du SI et CNTP, sauf indication contraire. | ||
L'histamine, amine naturelle, est une molécule de signalisation du système immunitaire, de la peau, de l'estomac et du cerveau des vertébrés. C'est un médiateur chimique synthétisé par les animaux dans de nombreux organes :
- par des cellules impliquées dans la réponse immunitaire, granulocytes basophiles et mastocytes ;
- par des cellules de la paroi de l'estomac régulant la sécrétion acide stomacale et ;
- par des neurones du système nerveux central pour le contrôle de l'éveil.
Selon les récepteurs qu'elle active, l'histamine provoque une réponse immunitaire, une sécrétion de suc gastrique et d'acide chlorhydrique, une dilatation des artérioles (petites artères), une contraction des bronches et des muscles du péristaltisme intestinal, une accélération de la fréquence cardiaque (tachycardie), un relâchement des contractions de l'utérus. Elle est en outre responsable d'un prurit (démangeaison de la peau). Dans le système nerveux central, elle assure le maintien de l'état de veille.
L'un des premiers effets repérés de l'histamine a été un rôle de médiateur dans la réponse allergique immédiate, mais l'ensemble des fonctions qu'elle a dans les cellules qui la contiennent et dans leur environnement est encore mal cerné. On sait qu'à très faible dose (non détectables par les techniques conventionnelles[3]), elle contribue à réguler divers processus physiologiques dont « la sécrétion d'acide gastrique, la neurotransmission, la régulation de la microcirculation et la modulation des réactions inflammatoires et immunologiques »[3]. Ces fonctions lui confèrent tantôt des propriétés pharmacologiques tantôt une certaine toxicité pour l'organisme[3].
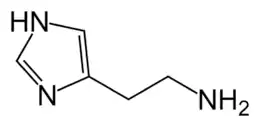
D'un point de vue chimique, l'histamine est la 2-(4-imidazolyl)éthylamine et a pour formule C5H9N3. C'est une amine hydrophile, vasoactive retrouvée dans tous les tissus [d'où son nom provenant du grec ancien ἱστός (histos = « tissu »)].
Localisation
On trouve de l'histamine dans tous les tissus des mammifères, les plus riches étant les poumons, le foie, la peau et la paroi de l'estomac.
- Réponse immunitaire
L’histamine est un médiateur chimique synthétisé par les granulocytes basophiles et les mastocytes (cellules appartenant à une variété de globules blancs). L'histamine est stockée dans les cellules et libérée dans des circonstances telles que les réactions d'hypersensibilité.
- Régulation acide de l'estomac
L'histamine est produite dans les cellules entérochromaffines de la paroi de l'estomac, dites cellules ECL, enterochromaffin-like cells. Ces cellules synthétisent et sécrètent l'histamine lorsqu'elles sont stimulées par l'hormone gastrine et la protéine PACAP (Pituitary adenylate cyclase-activating polypeptide).
- Neuromodulateur central
Les neurones histaminergiques sont localisés pour l'essentiel dans l'hypothalamus postérieur, au niveau du noyau tubéro-mamillaire[4]. Leurs axones innervent l'ensemble du cerveau, du tronc cérébral et de la moelle épinière. Ils sont actifs durant l'éveil mais pas durant le sommeil. Ils représentent le principal centre d'éveil du cerveau.
Métabolisme

L'histamine est synthétisée à partir de la L-histidine, un acide aminé essentiel, par l'enzyme histidine décarboxylase, sous l'effet du coenzyme pyridoxal-5'-phosphate[5]. Un inhibiteur de cette histidine décarboxylase, nommé la tritoqualine, est proposé en traitement d'appoint des affections allergiques[6].
Une fois formée, l'histamine est soit stockée soit dégradée. La dégradation est assurée par l'histamine N-méthyl transférase, localisée dans le cytoplasme de nombreuses cellules, neurones et cellules gliales au niveau central. Le produit formé est ensuite dégradé par la MAO-B (monoamine oxydase B).
Dans les neurones, le renouvellement de l'histamine stockée dans les vésicules présynaptiques est rapide. Sa demi-vie est d'environ 30 minutes. Par contre, dans les mastocytes, l'histamine stockée dans les granules de sécrétion, est complexée avec des résidus d'acides divers (héparine, sulfate de chondroïtine) et sa demi-vie dans ce cas est beaucoup plus longue, de l'ordre de plusieurs semaines.
Libération de l'histamine
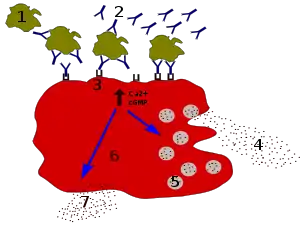
L'histamine est synthétisée par des enzymes du cytosol puis stockée (par transport actif) dans des granules (ou vésicules) de la cellule et ensuite relarguée par exocytose. Le signal déclencheur peut être de nature très différente suivant le type cellulaire.
- Pour les mastocytes tissulaires,
- lors de l'exposition à un PAMP ("Pathogen Associated Molecular Patterns" ou "Motif moléculaire associé aux pathogènes") du récepteur MRGPRX2,
- lors d'une phase de sensibilisation, une immunoglobuline E (IgE) spécifique d'un allergène (pollen, aliment) va s'attacher à sa surface. Lors de l'exposition suivante à l'allergène, la « dégranulation » est provoqué par la liaison de l'IgE avec cet allergène spécifique. Le mécanisme de cette exocytose dépend principalement de l'augmentation de la concentration en Ca2+ cytosolique.
- Pour les cellules ECL de la paroi de l'estomac, c'est la stimulation par la gastrine qui provoque la dégranu-lation (par le phénomène d'exocytose) des vésicules mastocytaires.
- Pour les neurones histaminiques, la fusion des vésicules de sécrétion amarrés à la membrane plasmique est déclenchée par l'augmentation du calcium libre Ca2+ intracellulaire provoqué par l'arrivée d'un potentiel d'action.
Dans la mastocytose, une maladie orpheline, la libération d'histamine provoque des symptômes.
Le syndrome d'activation mastocytaire est une état d'hyperactivation constante des mastocytes en raison d'une infection cryptique de type Bartonella.
Les récepteurs histaminergiques
L'histamine libérée va pouvoir agir sur des cellules spécifiques en se fixant sur celles qui portent des récepteurs histaminergiques.
Quatre types de récepteurs de l'histamine, nommés H1, H2, H3, H4, ont été décrits. Ils comportent tous sept hélices transmembranaires et sont couplés aux protéines G.
| Les quatre types de récepteurs histaminiques | |||
| Type | Protéine G | Localisation | Fonction |
| H1 | Gq/11 | muscles lisses | contraction des muscles lisses, bronchoconstriction |
| endothélium | vasodilatation, séparation des cellules endothéliales, responsable de l'urticaire et des douleurs dues aux piqûres d'insectes | ||
| système nerveux central | maintien de l'état de veille | ||
| H2 | Gs | paroi de l'estomac | régulation de la sécrétion de l'acide de l'estomac |
| H3 | Gi/o | autorécepteurs présynaptiques des neurones centraux | inhibe la libération de l'histamine, acétylcholine, noradrénaline, sérotonine… |
| H4 | Gi/o | cellules hématopoïétiques périphériques et de la moelle osseuse | - chimiotaxie des éosinophiles et mastocytes à l'histamine - libération d'IL-16 des lymphocytes T CD8 |
Histamine mastocytaire et allergies
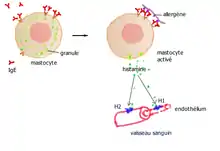
L'histamine libérée par les mastocytes va se fixer dans le voisinage, sur les récepteurs H1 des cellules endothéliales des vaisseaux sanguins et les récepteurs H1 des cellules musculaires lisses[6],[7]. Il en résulte une vasodilatation et la formation d'un œdème local accompagnant les allergies cutanées et nasales. La vasodilatation capillaire, provoquée probablement par la libération de monoxyde d'azote NO est à l'origine de la rougeur du visage et de certaines céphalées.
En cas de libération massive d'histamine, ces effets sont généralisés avec une chute tensionnelle importante.
L'histamine joue un rôle important dans les mécanismes de l'intolérance alimentaire, de l'allergie (elle est responsable de manifestations allergiques telles que les vasodilatations, le prurit et les œdèmes), de l'anaphylaxie, de l'urticaire, des inflammations et augmente pendant la réaction allergique.
Les antihistaminiques sont des médicaments qui inhibent les récepteurs histaminiques H1 (manifestations allergiques) et H2 (estomac). En bloquant les récepteurs de l'histamine, les antihistaminiques empêchent l'histamine d'agir. Il existe au moins un antihistaminique atypique utilisé en thérapeutique qui agit en inhibant la L-histidine décarboxylase (tritoqualine).
Histamine et sécrétion acide de l'estomac

L'histamine libérée par les cellules entérochromaffines-like ECL (après stimulation par la gastrine) se lie sur les récepteurs H2 couplés à une protéine Gs des cellules pariétales. La cascade de réactions qui s'ensuit se termine par la phosphorylation de la pompe à protons qui déverse alors des H+ dans la lumière de l'estomac.
L'agent de stimulation des cellules ECL est un peptide de 17 résidus, nommé la gastrine, sécrétée par les cellules endocrines G de l'antrum en réponse à une prise alimentaire. Elle se lie ensuite aux récepteurs CCK1 ou CCK2.
L'histamine est aussi libérée par les cellules entérochromaffines EC, de la paroi stomacale. De la même manière, l'histamine se lie aux récepteurs H2 des cellules pariétales de la paroi voisine et active les pompes à protons.
L’histamine neuromédiateur du système nerveux central

L'histamine est aussi un neurotransmetteur synthétisé et libéré par les neurones appelés « neurones histaminergiques ». Elle est stockée dans des vésicules (mais le transporteur responsable de cette localisation n'a pas encore été identifié). Elle est libérée à la suite d'un stimulus électrique et va se lier à des récepteurs post- ou présynaptiques.
On trouve le corps cellulaire de tels neurones dans une zone bien précise du système nerveux central : l'hypothalamus et plus précisément dans le noyau tubéromamillaire de l'hypothalamus postérieur. Ce noyau, situé à la périphérie des corps mamillaires, est formé de cellules de grandes tailles qui se projettent de façon diffuse sur le cortex cérébral et en particulier sur les régions ventrales (hypothalamus, le prosencéphale basal, amygdale) qui sont fortement innervées[8].
Les axones de ces neurones histaminergiques, largement répartis dans tout le cerveau, y libèrent de l'histamine, qui va se fixer sur les récepteurs H1 et H2 de neurones, tous postsynaptiques et des récepteurs H3 présynaptiques[6]. Ces derniers sont des autorécepteurs présynaptiques inhibiteurs. Les récepteurs H3 peuvent être aussi postsynaptiques et jouer le rôle de régulateurs de la libération des autres neurotransmetteurs. Ils sont largement distribués dans le striatum, le noyau accumbens et le cortex cérébral.
Le taux de décharge des neurones histaminergiques varie au cours du cycle veille-sommeil. Il est le plus grand durant l'éveil et le plus faible durant le sommeil paradoxal. On observe les mêmes variations cycliques dans la décharge de la noradrénaline des neurones du locus coeruleus ou la sérotonine dans les noyaux du raphé : activité maximale durant l'éveil, diminuée pendant le sommeil lent et disparition presque complète dans le sommeil paradoxal.
Des enregistrements de cellule histaminergique individuelle chez le chat ont montré une absence d'activité durant le sommeil mais une reprise d'activité un peu avant le début de l'éveil[9]. La libération importante d'histamine durant l'éveil et durant la phase précédant l'éveil suggèrent que ces neurones jouent le rôle de promoteur de l'éveil. De même, il a été observé qu'une micro-injection de muscimol, un agoniste puissant du GABA, dans le noyau tubéromamillaire, supprimait l'éveil et faisait entrer dans une phase de sommeil lent.
Finalement, les études montrent que l'activation des cellules histaminergiques dans l'hypothalamus postérieur produit une activation corticale et provoque le réveil.
Un déficit dans ce système histaminergique est probablement responsable de l'hypersomnie rencontrée dans la narcolepsie (maladie dans laquelle l'individu s'endort subitement).
Le rôle de l'histamine dans l'éveil explique aussi pourquoi les antihistaminiques utilisés contre l'allergie ont pour effet secondaire la somnolence : ils bloquent l’action de l’histamine. Mais les nouvelles générations d'antihistaminiques ne passent pratiquement pas la barrière hémato-encéphalique et ne devraient plus provoquer de sédation.
On sait aujourd'hui que, dans le cerveau, un autre type cellulaire est capable de synthétiser l'histamine, il s'agit des cellules microgliales. Ces cellules appartiennent à la famille des monocytes/macrophages.
Le système histaminergique cérébral module le rythme de veille/sommeil, mais aussi la prise alimentaire et/ou la prise de poids, ainsi que l'attention et la vigilance, sous le contrôle de l'hormone mélatonine.
Alimentation et histamine
L’intoxication histaminique, ou syndrome de pseudo-allergie alimentaire, provient de la consommation d’aliments renfermant de fortes quantités d’histamine[10].
Les principaux symptômes observés sont liés à l’effet vasodilatateur de l’histamine. Les symptômes les plus souvent rencontrés sont : rougeur facio-cervicale, éruption cutanée, œdème du visage, bouffées de chaleur, sensation de brûlure dans la gorge, démangeaisons, picotements de la peau. Ces symptômes cutanés sont assez spécifiques de l’intoxication histaminique, ils sont généralement accompagnés de signes généraux à type de céphalées, palpitations cardiaques, étourdissements. Enfin, des symptômes secondaires de nature gastro-intestinale peuvent apparaître : nausées, maux d’estomac, vomissements, diarrhée.
En général, les symptômes apparaissent rapidement, entre quelques minutes et quelques heures, puis disparaissent spontanément en quelques heures. Exceptionnellement, ils peuvent durer plusieurs jours dans les cas les plus graves. Cliniquement, cette intoxication mime une allergie alimentaire vraie avec pour conséquence une sous-estimation de son incidence[11],[12].
Aliments riches en histamine[13]
- Le chocolat.
- Les fromages affinés, comme le roquefort (particulièrement dans les zones les plus humides et salées[14]).
- Les poissons marinés tels le hareng, anchois, mahi-mahi et les sardines ; Les poissons dits scombroïdes, c'est-à-dire de la famille des scombridés (Espadon, marlin, thon, bonite et maquereau), les poissons les plus courants d'intoxication à l'histamine en cas de rupture de la chaine du froid). Il existe en France un "Plan de surveillance de l'histamine dans les produits de la pêche"; en 2006, sur 375 prélèvements, 10 non-conformités ont été mises en évidence par la DGAL[15].
- Le gibier faisandé.
- La levure de bière et d'autres aliments fermentés (vin, bière, choucroute).
Cela ne doit pas être confondu avec la libération d'histamine lors d'une allergie alimentaire.
Certaines formes d'intoxications alimentaires sont dues à la conversion de l'histidine en histamine dans des denrées alimentaires décomposées, comme du poisson, par l'action de décarboxylases microbiennes. Ainsi, le scombrotoxisme est une intoxication alimentaire due à la formation d'histamine après dégradation bactérienne de l'histidine, présente en grande quantité dans certains poissons comme les thons, maquereaux et bonites.
Réglementation[16]
Étant un indicateur de l'altération, les teneurs en histamine des poissons sont réglementées par l'Union européenne : le règlement (CE) n° 2073/2005 définit des critères de sécurité pour quelques espèces de poissons qui présentent de forts taux d'histamine. Ce même règlement indique également la méthode de quantification homologuée par HPLC. À ce jour, aucune réglementation ne limite les concentrations dans les autres denrées alimentaires.
Notes et références
- ↑ Masse molaire calculée d’après « Atomic weights of the elements 2007 », sur www.chem.qmul.ac.uk.
- 1 2 3 (en) « Histamine », sur ChemIDplus, consulté le 8 février 2009
- 1 2 3 L. Galoppin, C. Ponvert 1997) L'Histamine (Original Research Article) ; Revue Française d'Allergologie et d'Immunologie Clinique, Vol.37, no 7, 1997, Pages 865-880 (Résumé)
- ↑ H. Hass, P. Panula, « The role of histamine and the tuberomamillary nucleus in the nervous system », Nature Reviews Neuroscience, vol. 4,
- ↑ Kathleen M. Botham, David A. Bender et al., Biochimie de Harper, De Boeck Superieur, , p. 315
- 1 2 3 Yves Landry, Jean-Pierre Gies, Pharmacologie Des cibles vers l'indication thérapeutique, Dunod, 2009 (2e édition)
- ↑ LEMAITRE Florian, « Anti-histaminiques H1 (sauf comme anxiolytiques ou comme hypnotiques) », sur pharmacomedicale.org (consulté le )
- ↑ R.E. Brown, D.R. Stevens, H.L. Haas, « The physiology of brain histamine », Progress in Neurobiology, vol. 63, , p. 637-672
- ↑ Subimal Datta, Robert Ross MacLean, « Neurobiological mechanisms for regulation of mammalian sleep-wake behavior: reinterpretation of historical evidence and inclusion of contemporary cellular and molecular evidence », Neuroscience and Biobehavioral Reviews, vol. 31,
- ↑ « allergienet.com/fausses-allerg… »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- ↑ Fiche de description de danger transmissible par les aliments: Histamine ; AFSSA juin 2006 ; M. MALLE
- ↑ http://wwwbibli.vet-nantes.fr/theses/1999/boulay995/part1.pdf
- ↑ http://www.infectiologie.com/site/medias/_documents/officiels/afssa/Histamine090207.pdf
- ↑ Hervé This, « Attention à l'histamine dans les aliments ! », Pour la science, no 544, , p. 96.
- ↑ voir p 17, in Bilan des plans de surveillance et de contrôle mis en œuvre par la DGAL en 2006
- ↑ « Histamine », sur Anses (consulté le )
Voir aussi
Articles connexes
- Allergie
- Choc anaphylactique
- Anti-histaminique
- Mastocytose
- Syndrome d'intolérance à l'histamine
- Liste d'hormones
Liens externes
Bibliographie
- (fr) L. Galoppin, C. Ponvert 1997) L'Histamine (Original Research Article) ; Revue Française d'Allergologie et d'Immunologie Clinique, Vol.37, no 7, 1997, Pages 865-880 (Résumé)
- (en) Ritchie E. Brown, David R. Stevens, Helmut L. Haas (2001), The physiology of brain histamine (Review Article), Progress in Neurobiology, Vol.63, no 6, , Pages 637-672 (résumé)
- (en) R Leurs, M.J Smit, H Timmerman (1995) Molecular pharmacological aspects of histamine receptors (Revue de la littérature) Pharmacology & Therapeutics, Volume 66, Issue 3, 1995, Pages 413-463 (résumé)