Cèpe de Bordeaux

| Règne | Fungi |
|---|---|
| Division | Basidiomycota |
| Sous-division | Agaricomycotina |
| Classe | Agaricomycetes |
| Sous-classe | Agaricomycetidae |
| Ordre | Boletales |
| Sous-ordre | Boletineae |
| Famille | Boletaceae |
| Genre | Boletus |
| Section | Boletus |
- Boletus bulbosus Schaeff.[1]
- Boletus edulisf. arcticus Vassilkov[1]
- Boletus edulis f. edulis Vassilk[1]
- Boletus edulis f. laevipes (Massee) Vassilkov[1]
- Boletus edulis subsp. trisporus Watling[1]
- Boletus edulis var. arcticus (Vassilkov) Hlaváček[1]
- Boletus edulis var. laevipes Massee[1]
- Boletus edulis var. trisporus (Watling) anon. ined.[1]
- Boletus esculentus Pers.[1]
- Boletus filiae Gillet.[1]
Boletus edulis ou Cèpe de Bordeaux est une espèce de champignons basidiomycètes de la famille des Boletaceae que l'on rencontre dans l'hémisphère nord. Elle fait partie, avec Boletus aereus, Boletus reticulatus et Boletus pinophilus des quatre cèpes « royaux » les plus estimés. C'est l'une des espèces parmi les bolets qui est aussi appelée cèpe, entre autres noms vernaculaires ou commerciaux. La classification de cette espèce est en constante révision, notamment depuis que la phylogénie distingue une branche américaine et une branche européenne au sein du genre Boletus. Cet excellent champignon comestible très recherché est commercialisé et cuisiné mondialement sous diverses formes.
Généralités
Origine
Le Cèpe de Bordeaux, de son nom scientifique Boletus edulis, est un champignon des forêts, excellent comestible. Il est récolté par les amateurs avec peu de risque de confusion avec un bolet non comestible ou faiblement toxique qui ne sont de toute façon jamais mortels. Il pousse dans les clairières ombragées, et se développe de l'été à l'automne. Sa biologie, une des plus complexes, ne permet pas à l'homme de maîtriser son développement et encore moins sa culture. Il occupe une place de choix dans la gastronomie française où les recettes se déclinent par centaines. Il tiendrait son nom de sa présence massive sur les marchés de la ville bordelaise. Il fait partie d'un très vaste genre de plusieurs centaines de champignons à tube, les boletus. Au XXIe siècle, la phylogénie démontre que ses nombreuses variétés sont des espèces formant le vaste clade des edulis sensu lato, dont l'un est celui des edulis sensu stricto, maintenant divisé en une branche américaine et une branche européenne où se trouve le Cèpe de Bordeaux.
Les Boletus comestibles sont commercialisés en France sous le nom de Cèpe, dont le Cèpe de Bordeaux, maître élément de la gastronomie française.
Histoire

Les Romains consommaient déjà des bolets, mais le terme latin bōlētus englobait l'Amanite des Césars. Oublié, il ne revient sur les tables des nobles gourmets que vers 1760, par Stanislas Leszczynski, roi de Pologne installé en Lorraine[2]. Il règne depuis sans partage dans le Sud-Ouest depuis le début du XVIIIe siècle. Sa conserverie au naturel débute vers 1930, dans une gamme comprenant le foie gras, les escargots ou les haricots verts extra-fins au sein de sociétés comme Bordes, Bizac ou encore Rougié. Le Cèpe de Bordeaux frais ne sort de sa province que vers les années cinquante. Aujourd'hui sa consommation a pris une ampleur mondiale, mais le marché français n'utilise que la production régionale[3].
Edulis signifie en latin bon à manger[4].
Description générale
Le champignon est formé d'un chapeau, d'un pied et du mycelium qui lui permet pendant des années d'échanger avec certains arbres des substances nutritives. La surface du chapeau présente une couleur de brun clair à brun foncé, avec une marge ornée d'un fin liseré blanc. Adulte, le chapeau va s'étaler, prendre la forme d'un coussinet, ses bords vont se relever et son pied va s'enfler. La chair est blanche et ne change pas de couleur quand on tranche le champignon[5].
Récolte et préparation
En France, l'article 547 du code civil précise que les champignons appartiennent toujours au propriétaire du terrain sur lequel ils poussent, (forêt, prés ou champs), ce qui s'applique aussi aux forêts domaniales, bien que, dans la pratique, il existe une certaine tolérance limitée selon les circonstances aux besoins de la consommation familiale [6].
Le Cèpe de Bordeaux se récolte jeune - on l'appelle alors bouchon de champagne - ou adulte, mais il devient alors vite la proie des vers. Une petite brosse permet de nettoyer les restes de terre. Il se coupe à la base du pied, et il est déposé calé dans un panier pour le garder aéré. Les facteurs climatiques sont déterminants. Les cèpes de Bordeaux poussent après une période chaude interrompue par une forte averse ou une chute de grêle. Ils apparaissent alors après une dizaine de jours. Un petit champignon, le meunier, pousse parfois au même endroit. Il pousse sur des versants bien exposés au soleil couchant et bien abrités du vent, surtout si la période a manqué de soleil, dans les Ardennes par exemple. Il ne pousse pas souvent sous l'arbre mais dans le coupe-feu, probablement parce qu'il fait plus sec sous l'arbre et que le mycélium recherche ses radicelles en vue de l'association symbiotique : la mycorhize. La récolte débute une quinzaine de jours après celle des champignons des prés. Enfin, ils poussent rarement seuls. Le Cèpe de Bordeaux ne se lave pas et ne s'épluche pas. S'il roule, il se blesse et moisit. Dans un sachet de plastique, il macère et fermente. Lavé, il gonfle et ne peut pas être conservé[3].
Adulte, il faut retirer la mousse du bolet qui, si elle est comestible, devient spongieuse à la cuisson. Les tubes verts s'enlèvent facilement. Jeunes, les cèpes de Bordeaux sont blancs et restent fermes. Frais, jeunes et sans coups, ils garderont toute leur saveur au frigo pendant une semaine, sans se dégrader. Les cèpes peuvent être séchés mais deviennent fibreux. En conserve, ils gardent moins leur texture ferme et, surgelés, ils prennent du goût mais peuvent devenir spongieux. Les cèpes de Bordeaux sont poêlés quelques minutes dans un corps gras, agrémentés ou non par des herbes, et ensuite intégrés dans de très nombreuses recettes. Ils peuvent être consommés crus en carpaccio.
Confusion possible avec les autres cèpes
L'avis d'un spécialiste est nécessaire (pharmacien formé, association mycologique) avant d'en consommer cuits. Consommer les autres bolets crus, (sauf le seul véritable Cèpe de Bordeaux, ses quatre espèces proches et les espèces apparentées des autres continents), comme pour beaucoup de champignons est formellement déconseillé. Comme pour la plupart des champignons, la consommation d'exemplaire trop vieux n'est pas recommandée.

Quelques champignons ressemblent au Cèpe de Bordeaux, surtout lorsqu'ils sont jeunes. Si la confusion avec le Bolet Satan, dont la consommation ne provoque que des troubles digestifs, est peu probable car il a un pied bien rouge, le Bolet de fiel et le Bolet à beau-pied sont davantage sources de confusion car la distinction visuelle est difficile et la meilleure technique reste de goûter un morceau cru du champignon, l'amertume de ces deux derniers est évidente. Adultes les différences se marquent : le Bolet de fiel est moins trapu, et son pied est brun, le Bolet beau-pied garde sa mousse blanche et ferme et le pied rougit à sa base.
Taxonomie
Nom binomial accepté
- Boletus edulis (Bulliard 1781)
Synonymes
- Boletus edulis var. arcticus ((Vassilkov) Hlavácek 1994[7]) (synonyme)
- Boletus edulis f. arcticus (Vassilkov 1966[8]) (synonyme)
- Boletus edulis f. laevipes ((Massee) Vassilkov 1966[9]) (synonyme)
- Boletus edulis var. laevipes (Massee 1892[10]) (synonyme)
- Boletus edulis trisporus (Watling 1974[11]) (synonyme)
- Boletus solidus (Sowerby 1809[12]) (synonyme)
- Dictyopus edulis ((Bull.) Forq. 1890) (synonyme)
- Leccinum edule ((Bull.) Gray 1821[13]) (synonyme)
- Boletus esculentus Persoon (1825).
- Tubiporus edulis Karsten[14]
Historique

Boletus edulis a été décrit en 1782 par le botaniste français Pierre Bulliard. L'espèce porte encore son nom d'origine[15]. La date de début de la taxonomie fongique ayant été fixée au , pour coïncider avec la date de parution des travaux du « père de la mycologie », le naturaliste suédois Elias Magnus Fries, le nom avait besoin de l'aval de Fries (indiqué dans le nom par « : ») pour être considéré comme valide, car le travail de Bulliard était antérieur à cette date. On écrivait donc Boletus edulis Bull: Fr. Cependant, une révision de 1987 du Code international de nomenclature botanique fixe la date de départ de la classification au , date de la publication par Linné de son travail de pionnier, le Species Plantarum[16]. Le nom n'a plus besoin de la ratification de Fries pour être valide. Au début, d'autres noms ont été proposés comme Boletus solidus par le naturaliste anglais James Sowerby en 1809[17] et Leccinum edule par Samuel Gray[18]. Le transfert par Gray de l'espèce dans le genre Leccinum s'est révélé incompatible avec les règles de la nomenclature botanique et il est évident que Gray n'était pas familier avec les œuvres antérieures de Fries quand il a publié son classement des espèces de Bolet[19].
Nom normalisé
- Cèpe de Bordeaux[20]
Autres noms vernaculaires français

Noms vernaculaires dans d'autres langues
- Anglais : Penny bun, Cep ;
- Allemand : Gemeine Steinpilz, Fichtensteinpilz (pins), Herrenpilz, Edelpilz (précieux) ;
- Italien : Porcino ;
- en Ligurie, Funzo de castagna, Funzo neigro, Servajlo ;
- en Lombardie, Regùlat, Biancon, Brisott, Cappellet, Ferré levrin, Fonz ferré, Légorzéla, Legorsela, Nonna, Nÿna, Vairol, Nivariö ;
- dans le Piémont, Anvrioel, Bolé caréi, Bolé porcin, Funs capelet ;
- en Toscane, Ceppatello, Ghezza, Moccione, Moccolone ;
- en Vénétie, Boleo, Brisa, Brisot, Carpanote, Pressanella, Sbrisa ;
- dans le canton du Tessin, Copín, Cupít ;
- dans le Val d'Aoste, Fiorone ;
- Espagnol : Seta calabaza, Seta de calabaza, Calabaza.
Étymologie
Première espèce du genre Boletus, nom générique dérivé du latin bōlētus, « champignon » qui dérive à son tour du grec ancien βωλίτης, « champignon terrestre »[25]. Ce dernier mot dérive de βῶλος, bolos, signifiant « motte de terre » et, métaphoriquement, champignon[26]. Toutefois, le βωλίτης de Galien, comme les cèpes des écrivains latins comme Martial, Sénèque et Pétrone[27] sont souvent identifiés comme correspondant au plus prisé Amanita caesarea[28]. Le mot latin pourrait dériver du nom de la ville espagnole Boletum dont le nom actuel est Boltaña dans le Sud des Pyrénées qui est toujours connue pour ses fameux champignons[29].
L'épithète spécifique latine edulis signifie comestible[30].
Systématique
S'il apparaît que les mycologues étaient proches quant à leur classification classique, ils ne sont jamais parvenus à préciser les nuances entre variétés et espèces. Les analyses phylogénétiques ont permis une plus grande précision. Certaines espèces qui étaient considérées comme telles sont maintenant vues comme des variétés de Boletus edulis, par exemple selon Beugelsdijk et al Bolletus edulis var. pusteriensis, Boletus persoonii et Boletus venturii ne sont pas d'un point de vue moléculaire distinguables de Boletus edulis[31].
Classification classique ou morphologique
Boletus edulis est l'espèce type du genre Boletus, classée dans la famille des Boletaceae et dans l'ordre des Boletales. Dans la classification de champignons développée par Rolf Singer en 1986[32], il devient également l'espèce type du genre Boletus, un regroupement d'environ une trentaine de bolets unis par un certain nombre de caractéristiques :
- un goût doux ;
- une chair blanche qui ne change pas de couleur lorsqu'elle est exposée à l'air ;
- un dessin en mailles légèrement réticulées sur au moins la partie supérieure du pied ;
- une sporée jaune-brun ou brun olive ;
- des tubes blancs à l'état jeune, devenant jaunâtres à maturité, puis verdâtres au vieillissement ;
- des cystides peu colorées[33],[34].
Classification phylogénétique
Une analyse moléculaire[35] montre que les bolets dérivent tous d'un ancêtre commun et que les Boletales sont un clade distinct des Agaricales.
Cette classification phylogénétique, grâce à l'analyse moléculaire, confirme Boletus edulis dans le clade des Boletales, mais le révèle plus proche de Porphyrellus porphyrosporus ou de Strobylomyces flocoppus que de Boletus satanas. Il est également fort proche de Paxilus involutus. Chez Boletus edulis en Europe, quelques clades se précisent[31].
La position de l'espèce Boletus edulis au sein des bolétales est détaillée dans l'article Boletales.
Boletus edulis appartient à la section Boletus du genre Boletus. En Europe on rencontre quatre espèces dans cette section. Ce sont ces quatre espèces que l'on appelle des cèpes.
| Cèpe |
| ||||||||||||||||||
Origine tropicale et ancêtre commun
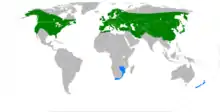
Boletus edulis est un champignon récent [36], la diversification des Cèpes est située entre 34 Mo d'années[37] et 44 Mo d'années pendant la période géologique de l'Éocène[38], la diversification entre agaricales et boletales étant datée entre 139 Mo et 178 Mo d'années.
En 2010[39], apparaissent les nouveaux clades des edulis sensu lato et le clade des edulis sensu stricto qui précisent la situation de toute une série de variétés qui deviennent des espèces spécifiques et de quatre nouvelles espèces dans le sud de la Chine, en Corée, aux Philippines et en Amérique centrale.
L'étude précise qu'un spécimen de Boletus variipes récolté aux Philippines est défini comme le plus récent ancêtre commun du groupe edulis et conclut à l'origine paléotropicale probable de la symbiose ectomycorhizienne.
Description morphologique
Sporophore
Chapeau

Le champignon forme un grand chapeau brun convexe, sa forme ronde rappelle celle d'un bouchon de champagne ; sa marge est enroulée. En vieillissant, il s'étale tout en restant épais, charnu et convexe (8 à 20 cm de diamètre), parfois pulviné. Rarement, dans des circonstances hygrométriques favorables, il peut atteindre 35 cm de diamètre et peser jusqu'à 3 kg.
Chair

La chair du cèpe, l'hyménophore et le stipe, sont de couleur blanche et brun-vineux sous la cuticule. La chair est épaisse et ferme, souvent attaquée par des larves de diptère, comme Cordyla crassipalpis et des morsures de limaces.
Cuticule
La cuticule du chapeau a une couleur qui varie du brun noisette au brun ochracé. Lisse jeune, elle peut devenir viscidule avec l'humidité. Sur les sujets âgés on peut la peler.
Marge
Le chapeau est souvent recouvert d'une pruine blanchâtre qui disparaît avec l'âge en laissant un bord ourlé de blanc et devient appendiculée.
Hyménium
L'hyménium se présente, comme chez tous les bolets, sous forme de tubes, appelés parfois foin en français. Les pores et les tubes des jeunes cèpes sont blancs, fermes et comestibles, puis deviennent jaunes et ensuite vert olive avec l'âge. Ces tubes ont une insertion adnée sur le pied. Vieux, les tubes, s'ils restent comestibles, ne sont plus mangeables, car déliquescents à la cuisson. On peut utiliser cette particularité et les surgeler en cubes pour parfumer naturellement un plat.
.jpg.webp) |
 | |
Le stipe peut avoir une forme allongée ou bulbeuse | ||
Stipe
Le stipe (pied) de couleur blanche ou jaunâtre, peut atteindre jusqu'à 25 cm de hauteur et 10 cm d'épaisseur. Il est trapu à son jeune âge et devient ensuite cylindrique. Il peut devenir assez long et fin si la végétation ne fait pas obstacle à sa pousse. Il reste cependant toujours renflé à la base (6 à 20 x 2 à 5 cm). D'une couleur beige pâle, il est partiellement recouvert d'un réseau de réticulations (maillage beige).
Sporée
Les tubes libèrent des spores brun-olivâtre.
Variétés

La variété grandedulis correspond peut-être à une autre espèce de californienne, Boletus grandedulis, qui pousse en mycorhize avec Pinus muricata[40]
Habitat - Écologie
Habitat
Le cèpe de Bordeaux est un champignon mycorhizien, c'est-à-dire qu'il vit en symbiose avec certains arbres hôtes comme le chêne, le châtaignier, le hêtre, l'épicéa et le sapin pectiné (mais pas avec le sapin de douglas, dont les plantations en masse constituent une menace pour le cèpe). Il pousse le plus souvent sur sols acides, dans les endroits dégagés ou aérés, les clairières, les talus bordés d'arbres et les bords des chemins. Cependant, il affectionne également les sous-bois denses et peu exposés à la lumière, comme ceux formés par les jeunes plantations d'épicéas.
Développement
Au printemps, le mycélium commence à pousser dans le sol à partir des mycorhizes.
En été, les filaments se développent de façon plus ou moins importante selon le niveau des précipitations.
À la fin de l'été et jusqu'à la fin de l'automne, les chocs thermo-hydriques (chute rapide de la température sous 15 °C et grosse pluie) permettent de concentrer les filaments en paquets qui donnent naissance dix jours plus tard aux sporophores, qui est la partie cueillie pour être consommée. Pour que les champignons se développent bien, la température ne doit pas être trop chaude lors de la semaine suivant le choc thermo-hydrique.
C'est pour cette raison que l'on trouve souvent les cèpes sous leurs arbres hôtes dix jours après une pluie importante.
Le sporophore pousse jusqu'à épuisement du mycélium. Suivant les conditions climatiques, dans le Nord de la France par exemple, ils peuvent apparaître en fin d'été et développer de nouveaux sporophores fin d'automne.
Espèces associées

La poussée du cèpe de Bordeaux comme celle des chanterelles est fréquemment précédée, de quelques jours, de celle de l'Amanite tue-mouches ou l'Amanite rougissante qui appartiennent à la guilde écologique des champignons ectomycorhiziens[41].
Boletus edulis est aussi souvent associé à Clitopilus prunulus, de son nom vernaculaire le Meunier ou mère des cèpes qui peut soit le précéder, soit lui succéder, et qui pousse dans les bois clairs de feuillus ou de conifères (notamment les jeunes plantations d'épiceas)[42].
Légendes urbaines
Un cèpe ne pousse pas du jour au lendemain. Du petit bouchon de champagne visible jusqu'à maturité (tubes verts, marge relevée), on compte généralement 5 à 6 jours[43].
Contrairement aux croyances populaires, les phases lunaires n'influencent pas l'apparition des cèpes et ils ne poussent pas plus vite pendant la nuit.
Risques de confusion
- On peut décrire deux formes de Boletus edulis : la forme des montagnes et des résineux, au stipe bulbeux et au chapeau brun sombre pouvant faire penser à Boletus aereus et la forme des feuillus de la plaine, au stipe allongé et au chapeau pâle pouvant faire penser à Boletus aestivalis.
- Bolet amer Tylopilus felleus, comestible mais immangeable par son goût amer.
Anciennes espèces et variétés
Les études actuelles tendent à diviser le clade Boletus edulis en de nombreuses espèces, ce que la morphologie avait détecté. Voici quelques exemples européens et américains, mais il est possible que l'on découvre de nouvelles différences en Asie. Le Phylogramme du clade edulis sensu stricto présente certaines de ces variétés, maintenant considérées comme des espèces
Europe
D'anciennes classifications parlaient de variétés, par exemple Boletus edulis var. pinicola. Ces variétés sont devenues des espèces à part entière comme Boletus pinicola, etc. Plusieurs champignons européens et américains qui étaient autrefois considérés comme des variétés de Boletus edulis se sont révélés par l'analyse phylogénétique moléculaire des espèces distinctes.
- Bolet albinos - Bolet de Persoon
Boletus persoonii, qui était érigé autrefois en espèce, est maintenant rattaché à Boletus edulis.[44] Une espèce similaire existe aux États-Unis, nommée Boletus barrowsii.
Amérique du Nord
Une sous-espèce de l'Ouest nord-américain de son nom vernaculaire Bolet roi de Californie, Boletus edulis var. grandedulis, est une grande variété de couleur foncée a été publiée en 2008[45]. D'autres espèces sont également proposées en Amérique du Nord. Le Cèpe d'Amérique, Boletus chippewaensis[46] est fort proche par son aspect du Boletus edulis européen. Enfin, Boletus subcaerulescens qui présente une cuticule plissée et alvéolée caractéristique unique.
Intérêt culinaire

Conservation
Disponible frais à l'automne, Boletus edulis est un excellent champignon comestible à la douce saveur de noisette que l'on peut consommer aussi bien cru, comme un carpaccio, que cuit, conservé surgelé, en boite ou bocal, en saumure ou dans l'huile et même séché.
Sa consistance est très bonne en conserve. Les Italiens le conservent dans de l'huile. Boletus edulis est aussi l'un des rares champignons qui peuvent être confits au vinaigre.
Il est aussi séché et garde sa saveur après séchage, il est alors reconstitué et utilisé en cuisine, mais devient un peu fibreux.
Richesse nutritive et risques pour la santé
Boletus edulis est pauvre en glucides digestibles et lipides mais riche en protéines, vitamines, sels minéraux et fibres alimentaires. Séché, on peut aussi l'utiliser au moulin.
Ce champignon produit une variété de composés organiques à large spectre d'activité biologique, comme l'ergostérol, une lectine, protéine de liaison spécifique à un sucre, des composés antiviraux, des anti-oxydants et des phytochélatines, qui lui confèrent une grande résistance aux Élément-trace métallique à des doses mortelles pour de nombreux autres organismes[47]. Comme les autres macrochampignons, il concentre plus fortement ces élément toxiques dans son carpophore. Ce pourquoi, comme tous les champignons, il ne devrait jamais être récolté dans les zones polluées ou à teneurs naturellement haute en métaux lourds et métalloïdes toxiques.
Tous les macrochampignons sont en effet connus pour facilement bioaccumuler des métaux et des métalloïdes toxiques, dont le Mercure[48], notamment dans les zones polluées (par l'activité minière par exemple)[49], plus ou moins selon le substrat surlequel ils se nourrissent[50]. Une étude a comparé les teneurs des carpophores de Boletus edulis mercure total (HgT) et en méthylmercure (MeHg, beaucoup plus toxique) selon de leur stade de développement (stade bouton, jeune-blanc, grand-blanc et grand-jaune). Pour chaque stade, on a comparé les teneur de mercure (méthylé ety non méthylé) dans le champignon cru et une fois cuit (braisés dans ce cas).
Le mercure pur est un métal très volatil quand on le chauffe (mais sa vapeur est hautement toxique). Cette étude a cherché à évaluer l'effet de la cuisson (braisage) sur ces teneurs, et d'estimer l'apport en mercure et méthylmercure pour une personne consommant un seul repas de bolets (sur la base du poids total (humide) (ww) et poids sec (dw). Elle a montré que dans les carpophores braisés, les taux de HgT variaient de 0,2668 ± 0,0090 à 0,5434 ± 0,0071 mg kg−1 ww selon le stade de développement du champignon ... et de 0,1880 ± 0,0247 à 0,2929 ± 0,0030 mg kg−1 en poids humide cru (ww).
Les jeunes carpophores bruts (au stade bouton) présentaient les teneurs les plus haute en mercure total (par rapport aux stades ultérieurs de développement), mais la part du MeHg y étaient plus faible. Et, après braisage, le jeune carpophore présentait plus de MeHg qu'aux stades de maturité plus avancés. Le MeHg comptait pour 1,9 ± 0,7 % du mercure total (HgT) dans les champignons crus et pour 1,4 ± 0,3 % dans le champignon cuit (braisés). La cuisson augmente les teneurs moyennes en HgT et MeHg dans les farines de champignons frais de 52 ± 31 % et 53 ± 122 % respectivement, mais une réduction de 40 ± 14 % et 40 ± 49 % respectivement est observée sur une base sèche. Concernant le seul mercure, selon les auteurs, dans ce contexte, « Les apports potentiels de THg et de MeHg provenant des repas braisés de B. edulis étudiés étaient donc faibles peuvent être considérés comme sûrs ».
Mais une étude de risque complète doit aussi tenir compte des autres métaux lourds et des métalloïdes que ce champignon peut bioconcentrer.
Culture et récolte

Tentative de culture
Comme d'autres champignons à mycorhize stricte, Boletus edulis a échappé aux tentatives de culture[51],[52]. Les résultats de certaines études suggèrent que des composantes inconnues de la microflore du sol pourraient être nécessaires à Boletus edulis pour réussir à développer sa relation mycorhizienne avec la plante hôte[53],[54],[55].
En France
L’état de la situation amène à une certaine prudence en matière de prévision sur la domestication des cèpes. L’acquisition de données écologiques et physiologiques plus précises va certainement aider à prendre en compte la composante mycologique dans la gestion de territoires à forte vocation forestière, comme le Limousin, l’Auvergne, le Morvan et les Vosges… La plantation d’arbres mycorhizés demeure du domaine expérimental. Les rendements de mycorhization obtenus par les différents auteurs restent assez faibles, même si chaque laboratoire progresse dans une certaine discrétion vers des résultats plus applicables sur le plan économique. De toute façon, le choix des souches, la qualité de l’inoculum, les conditions de réalisation de l’association restent des éléments à améliorer. Les éventuelles relations avec les micro-organismes de la rhizosphère[56] n’ont pas été précisées pour les cèpes. Enfin, pratiquement rien n’est connu sur les relations entre le champignon introduit précocement et le développement fongique qui s’établit après la plantation. Or, la domestication du cèpe n’a d’intérêt qu’avec le développement des sporophores, attendue 10, 15, voire 20 ans après la plantation[57].
Culture aléatoire et récolte

Les cèpes sont donc toujours des champignons sauvages impossibles à cultiver de façon industrielle. L'amateur peut simplement se contenter de composer un environnement favorable à leur développement. Les récoltes se font dans les bois composés de chênes et châtaigniers vieux d'au moins 15 ans.
Si le terrain ne donne pas naturellement des cèpes, il est possible de tenter de l'ensemencer en disséminant de vieux cèpes sur le sol ou en mettant des épluchures dans une taupinière en s'abstenant de faire la chasse aux taupes car elles propagent le mycélium[58].
Lors de la récolte, on conseille souvent de couper le cèpe plutôt que de l'arracher afin d'en retrouver au même endroit la saison suivante. Toutefois, la meilleure solution semble être de saisir le cèpe par son pied, et de le faire tourner d'un quart de tour et de tirer, ce qui évite une éventuelle infection du mycélium[59].
Mettre le champignon nettoyé dans un panier (et non dans un sac plastique). En effet, cela permet au cours de la récolte de disséminer les spores de champignon passant à travers les mailles d'un panier en osier. Par ailleurs cela permet une meilleure préservation de la récolte[60].
Paramètres climatiques observés
Analyse des paramètres climatiques dans le Sud de la France
Il est évident, pour tout observateur qu’il y a de bonnes et de mauvaises années pour le développement des sporophores. Parmi les explications parfois originales avancées par les ramasseurs de champignons[61], la relation avec le climat local ressort comme une constante. Pour aller plus loin dans l’analyse du phénomène, un dispositif de collecte de données agro-météorologiques a été mis en place en Aquitaine, Midi-Pyrénées, Limousin, Poitou-Charentes. Une cinquantaine de parcelles ont été choisies d’abord pour être des sites connus de pousse de Boletus edulis, Boletus aereus ou de Boletus reticulatus. Elles sont équipées de matériel d’agro-météorologie, pluviomètres, thermomètres (sol et air) et de tensiomètres. À côté des relevés climatiques et des analyses de sols, les pousses de champignons sont notées (dates et quantités, identification, localisation). Le contexte (précédent cultural, flore du sous-bois, autres espèces de champignons, état du peuplement forestier…) fait l’objet de relevés réguliers.
Développement en période pluvieuse chez Boletus aereus et Boletus reticulatus
Les événements qui ont conduit au développement de ces deux espèces apparaissent assez nettement dans une chênaie des Landes. En juillet, une période pluvieuse (80 mm) d’une semaine est suivie d’une baisse de température du sol (à –10 cm); les premiers sporophores apparaissent 7 jours après, la pousse se prolongeant pendant 12 jours (avec une pluviométrie importante). Un scénario voisin se reproduit à la fin août. Dans les deux cas, une remontée des températures coïncide avec l’arrêt du développement des sporophores. Les données collectées dans d’autres régions françaises (Dordogne, Lot, Gironde, Tarn et Garonne) confirment ces observations pour Boletus edulis. Toutefois, pour cette espèce un peu plus tardive, une baisse de la température pendant la phase de développement des sporophores peut arrêter définitivement la production.
Nécessité d'un choc thermique par cumul d’heures “froides” chez Boletus edulis
Les résultats actuellement connus peuvent être résumés[62] comme suit :
- une pluviométrie forte, supérieure à 50 mm sur une décade, assure une humidité persistante dans le sol (tensiomètre saturé plus de 5 jours) ;
- une chute des températures, air et sol, entraîne un refroidissement du sol dans les 10 premiers centimètres; le plus souvent, l’induction du développement des sporophores est liée à un écart de 5 °C environ par rapport à la température moyenne initiale ; le nombre d’heures pendant lequel cet écart se manifeste doit être, en cumulé, de 20 heures pouvant se répartir sur 3 à 5 nuits ; l’analyse des situations conduit à la notion de cumul d’heures “froides” ;
- aucun sporophore n’apparaît si les températures du sol sont supérieures à 20 °C ou inférieures à 10 °C pour Boletus edulis, à 12 °C pour les autres espèces ;
- en général, le choc thermique a lieu pendant ou après la période de pluie; il a cependant été observé quelques cas où la pluie a eu lieu 3 à 5 jours après la baisse de température ;
- les premiers sporophores sont observés 6 à 10 jours après le choc thermique si la température est supérieure à 15 °C, 8 à 15 jours si elle est inférieure;
- un nouvel abaissement de la température du sol d’au moins 3 °C pendant la période de développement arrête le processus.
Pendant la période de développement de fin d’été ou d’automne, il n’est pas encore possible de distinguer nettement les événements inducteurs selon les espèces de cèpes et leur éventuelle thermophilie[63].
Cependant, Boletus aereus et Boletus aestivalis commencent à se développer plus tôt, de juin à mi-août, le rôle du choc thermique semble moindre, avec effet majeur de l’eau que ce soit des pluies ou des arrosages.
Propriétés
Composition nutritive

Source nutritionnelle
Boletus edulis constitue une source nutritionnelle qui, bien que n'étant pas riche en glucides ou en lipides facilement absorbables, contient des vitamines, des minéraux et des fibres alimentaires. Les champignons frais contiennent plus de 80 % d'eau[64]. Bien que les valeurs déclarées aient tendance à différer quelque peu, la teneur en eau peut être affectée par la température ambiante et l'humidité relative pendant la croissance et le stockage, ainsi que la quantité relative d'eau qui peut être produite à la suite des processus métaboliques normaux pendant le stockage[65].
Composition biochimique
Boletus edulis frais présente les valeurs nutritives suivantes basées sur des analyses biochimiques[note 2]
- Valeur nutritive par 100 g :
- Énergie : 342,4 kJ (81,8 kcal)
- Lipides : 1,70 g
- Protéines : 7,39 g
- Thiamine (vitamine B1) : 0,105 mg (9 %)
- Riboflavine (vitamine B2) : 0,092 mg (8 %)
- Niacine (vit. B3) : 6,07 mg (40 %)
- Acide pantothénique (B5) : 2,64 mg (53 %)
- Vitamine B6 : 0,051 mg (4 %)
- Folate (Vitamine B9) : 290 mg (73 %)
- Vitamine C : 4,21 mg (5 %)
- Calcium : 1,195 mg (0 %)
- Fer : 0,739 mg (6 %)
- Phosphore : 22,26 mg (3 %)
- Potassium : 203,3 mg (4 %)
- Zinc : 4,172 mg (44 %)
Les pourcentages sont relatifs à des recommandations américaines spécifiques aux adultes[66].
Glucides
Les glucides constituent la majeure partie des organes du sporophore, comprenant 9,23 % du poids frais comme la liste le présente, et 65,4 % du poids sec[67]. Le volet contient les glucides glucoses monosaccharides, le mannitol et de l'α-tréhalose, le polysaccharide de glycogène, et insoluble dans l'eau des polysaccharides structuraux comme la chitine, qui représente jusqu'à 80-90 % de matières sèches des parois cellulaires du champignon. La chitine, l'hémicellulose et la pectine - comme les glucides non digestibles - par les humains, contribuent à la proportion nutritionnellement souhaitable, renforcée de fibres insolubles chez Boletus edulis[68].
Lipides
Le total des lipides[69],[note 3], ou matières grasses brutes, représente 2,6 % des matières sèches du champignon. La proportion d'acides gras (exprimée en % du total des acides gras) sont :
- l'acide linoléique, 42,2 %
- l'acide oléique, 36,1 %
- l'acide palmitique, 9,8 %
- l'acide stéarique, 2,7 %
- l'acide linolénique, 0,2 %
Acides aminés
Une étude comparative de la composition des acides aminés de onze espèces portugaises de champignons montre que Boletus edulis possède la plus haute teneur totale en acides aminés[note 4], d'environ 2,3 g par 100 g de champignons séchés. Ce total comprend un échantillonnage complet d'une vingtaine d'acides aminés essentiels et non essentiels[70]. L'analyse des acides aminés libres (c'est-à-dire ceux non liés en protéines) ont révélé que la glutamine et l'alanine en sont les principaux acides aminés (chacun pour environ 25 % des composés du total) ; une analyse séparée a conclu que la lysine est un autre composé prédominant[71]
Métaux et minéraux
Les valeurs rapportées sur la composition et les concentrations de traces de métaux et des minéraux chez Boletus edulis ont tendance à différer considérablement, tout comme dans le sporophore s'accumule différents éléments à différents degrés. Comme c'est généralement le cas pour les autres champignons, cette concentration des éléments dans le sporophore reflète souvent la concentration du sol en ces éléments bio disponibles, là où le champignon a été récolté[72].
En général, Boletus edulis contient des quantités appréciables de sélénium (13-17 ppm), un oligo-élément essentiel pour une bonne santé[73]. Cependant, la biodisponibilité du sélénium présent dans le champignon est faible[74].
Vitamines D
Les sporophores entiers et frais contiennent également environ 200 mg de vitamine D2 pour 100 g de poids sec[75]. La teneur relativement élevée en ergostérol contenu (voir prochaine section) par les sporophores peut en faire un champignon nutritionnellement intéressant pour les végétariens et les végétaliens, qui ne pourrait pas autrement recevoir un apport limité de vitamine D[76].
Ergostérol
Les sporophores de Boletus edulis contiennent environ 500 mg d'ergostérol par 100 g de champignons séchés[75]. L'ergostérol est un stérol composé commun dans les champignons. De plus, les sporophores ont environ 30 mg de peroxyde d'ergostérol par 100 g de champignons séchés. Le peroxyde d'ergostérol est un stéroïde dérivé avec un large spectre d'activité biologique, y compris d'activités antimicrobiennes et anti-inflammatoires, et une cytotoxicité de lignées de cellules de diverses tumeurs cultivées en culture en laboratoire[77].
Résistance aux métaux lourds
La phytochélatine confère au Boletus edulis une résistance aux métaux lourds toxiques comme le cadmium[78].
Lectine
Le champignon contient également un sucre à protéines contraignantes, appelée lectine, qui a une affinité pour les sucres xylose et mélibiose. La lectine est mitogène - c'est-à-dire qu'elle peut stimuler les cellules pour démarrer un processus de division cellulaire, d'où résultera la mitose. En outre, la lectine a des propriétés antivirales. Elle inhibe l'immunodéficience humaine du virus enzyme transcriptase inverse[79].
Activité antivirale
D'autres études suggèrent que Boletus edulis a également une activité antivirale contre les virus[80] et le virus de la mosaïque du tabac cultivé dans la culture[81]. Ces composés antiviraux de champignons sont un sujet d'intérêt de la recherche biomédicale pour leur potentiel à faire progresser la connaissance de la réplication virale, et de la mise au point de nouveaux médicaments dans le traitement de maladies virales[82].
Capacités antioxydantes
Les sporophores ont une haute capacité antioxydante, dont la raison probable est la combinaison de différents acides organiques (comme les acides oxalique, citrique, malique, succinique et fumarique), composés phénoliques et des alcaloïdes. L'activité antioxydante la plus élevée se trouve dans l'hyménophore des champignons[83]. Par ailleurs, les sporophores contiennent 528 mg d'ergothionéine par kilogramme de champignons frais de ce composé antioxydant. Cette valeur est la plus élevée parmi les nombreux produits alimentaires testés dans une étude[84].
Propriétés anti-cancéreuses
On pensait avoir trouvé des propriétés anti-cancéreuses en fonction des résultats de recherches hongroises menées dans les années 1950[85], mais d'autres études, menées plus tard aux États-Unis n'ont malheureusement pas confirmé ces résultats[86].
Allergies
De très rares cas d'allergies[87] par inhalation et ingestion de Boletus edulis ont été signalées, créant des réactions anaphylactiques manifestes et confirmées par des tests cutanés avec Boletus édulis cru, démontrant in vivo la capacité des mastocytes cutanés à réagir au contact de cet allergène spécifique à travers la liaison à l'immunoglobuline E (IgE), les symptômes étaient reproductibles après un test par inhalation[88]. Il n'a pas encore été déterminé quelle était la molécule responsable.
Galerie d'images
 Pied ventru
Pied ventru Pores blancs,
Pores blancs,
stipe réticulé.jpg.webp) Pores olivacés,
Pores olivacés,
marge relevée. deux cèpes
deux cèpes
Voir aussi
Articles connexes
- Boletus pinophilus
- Boletus chippewaensis
- Boletus subcaerulescens
- Boletus persoonii
- Anastomose (mycologie)
- Cèpe
Lien externe
- (en) Référence Index Fungorum : Boletus edulis (+ MycoBank)
- Référence Cèpe de Bordeaux
Bibliographie en français
- Becker G. et all[89] : Le guide des champignons, Reader's Digest, 1982 - (ISBN 2-7098-0031-4)
- Bon Marcel : Champignons de France et d'Europe occidentale, Flammarion, Édition de 2004, (ISBN 978-2082013215)
- Courtecuisse Régis & Duhem Bernard : Guide des champignons de France et d'Europe, Delachaux & Niestlé, 1994-2000.
- Gerthard Ewaldt : Guide Vigot des champignons, Vigot, 1999 - (ISBN 2-7114-1413-2)
- Jordan Peter & Steven Wheeler : Larousse saveurs - Les champignons, Larousse, 1996 - (ISBN 2-03-516003-0)
- Laessoe Thomas & Del Conte Anna : L'Encyclopédie des champignons, Bordas, 1996, (ISBN 2-04-027177-5)
- Phillips Roger : Les champignons, Solar, 1981 - (ISBN 2-263-00640-0)
- Romagnesi Henri : Petit atlas des champignons, Bordas, 1970.
Notes et références
Notes
- ↑ Les données sont basées sur les descriptions de la littérature, en prenant compte des plantes hôtes et des limites des zones climatiques. Ont été utilisées les sources suivantes : Hall et al, p. 45, Wang et al., p. 227, Ciesla : p. 86, Fischer et al. p. 96, Masuka: p. 69, Adhikary et al. p. 119, Giri et al. p. 68
- ↑ Les valeurs nutritives sont basées sur des analyses biochimiques sur des spécimens turcs, conduites par Çaglarlrmak et ses collègues à la Faculté d'agriculture, Food Engineering Department, Gaziosmanpasha University. Source : Çaglarlrmak N, Ünal K, Ötles S.. "Nutritional value of edible wild mushrooms collected from the Black Sea region of Turkey" (PDF). Micologia Aplicada International 14 (1): 1–5. 2001
- ↑ Une analyse indienne comparable présente des valeurs quelque peu différentes: total des lipides: 3,3 % de matière sèche; acide palmitique: 21,6 % ; acide stéarique: 9,1 % ; acide oléïque: 31,1 % ; acide linoléique : 33,8 %, et acide linolénique : 1,7 %. Source: Kavishree S, Hemavathy J, Lokesh BR, Shashirekha MN, Rajarathnam S., Fat and fatty acids in Indian edible mushrooms. Food Chemistry 106: 597–602. 2008
- ↑ Les autres espèces étudiées sont Suillus bellinii, Suillus luteus, Suillus granulatus, Tricholomopsis rutilans, Hygrophorus agathosmus, Amanita rubescens, Russula cyanoxantha, Tricholoma equestre, Fistulina hepatica, et Cantharellus cibarius.
Références
- 1 2 3 4 5 6 7 8 9 10 BioLib, consulté le 10 novembre 2017
- ↑ Pierre Thibaud, Éloge du Cèpe, 1990, cite Stanislas Leszczynski.
- 1 2 Le Cèpe, Spécial Champignon magazine, collectif, Secret et plaisir, Bordas, 2000.
- ↑ Dictionnaire Gaffiot
- ↑ Jean-Louis Lamaison, Jean-Marie Polese, Encyclopédie visuelle des champignons, éditions Artemis, , p. 57-58
- ↑ L'ami des jardins, À la découverte des champignons 1995.
- ↑ Hlavácek Jiri, Prehled našich hub hribotvarých - Boletales, (24), In: Mykologický Sborník 71(1):1–9 + 1 pl. 1994,
- ↑ Vassilkov, Bekyi Grib: 16, 1966
- ↑ Vassilkov, in: Bekyi Grib:13, 1966
- ↑ Massee, in: Brit. Fung.-Fl. 1:284, 1892
- ↑ Watling, in: Notes R. bot. Gdn Edinb. 33(2) p. 326, 1974
- ↑ Sowerby, in, Col. fig. Engl. Fung. Mushr. (London) 3, pl. 419, 1809
- ↑ Gray, in: Nat. Arr. Brit. Pl. (London) 1, p. 647, 1821
- ↑ Maublan A. 6e Édition revue par Viennot-Bourgin, II Atlas, p. 166
- ↑ Bulliard JBF., Herbier de la France. Vol 2, Paris, France, P.F. Didot, (lire en ligne), p. 49–96, plate 60
- ↑ (en) Esser K, Lemke PA., The Mycota : A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research, Heidelberg, Germany, Springer, , relié (ISBN 978-3-540-66493-2, LCCN 94019413), p. 81
- ↑ (en) Sowerby J., Coloured Figures of English Fungi, vol. Volume 4, Londres, J. Davis, , p. 199
- ↑ Gray SF., A Natural Arrangement of British Plants, Londres, Baldwin, Cradock, and Joy, Paternoster-Row, (lire en ligne), p. 647
- ↑ (en) Šutara J., « Leccinum and the question of superfluous names (Fungi: Boletaceae) », Taxon, vol. 34, no 4, , p. 678–86 (DOI 10.2307/1222214, JSTOR 1222214)
- ↑ SMF, Société Mycologique de France, Comité des noms Français
- ↑ Maublan, A. 6e Édition revue par Viennot-Bourgin, II Atlas, p. 166
- ↑ Maublan, A. ibidem, p. 166
- ↑ Philips, Solar
- ↑ Jean-Marie Polese, Champignons de France=éditions Artemis, , p. 47.
- ↑ (la) DP. Simpson, Cassell's Latin Dictionary, Londres, Cassell Ltd, , 5e éd. (ISBN 978-0-304-52257-6), p. 883Langue originale : allemand
- ↑ (en) Liddell HG, ScottR., A Greek-English Lexicon, United Kingdom, Oxford University Press, (ISBN 978-0-19-910207-5, LCCN 33031054)Langue originale : grec ancien
- ↑ Howell Peter, A Commentary on Book One of the Epigrams of Martial, The Athlone Press, 1980 p. 152-3. Howell doute de cette identification, et mentionne l'idée avancée par Augusta A. Imholtz Jr.
- ↑ (en) Ramsbottom J., Mushrooms & Toadstools, London, England, Collins, (ISBN 978-1-870630-09-2), p. 6Langue originale : grec ancien
- ↑ Imholtz Augusta A. Jr., Fungi and piace-names, the origin of boletus, in AJP Vol.98, 1977 p. 71 f., that the Latin word may derive from the Spanish town Boletum, modern-day Boltaña, south of the Pyrénées, which is still famous for its mushrooms
- ↑ (en) Jamieson A, Ainsworth R, Morell T., Latin Dictionary: Morell's Abridgment, Londres, Moon, Boys & Graves, (lire en ligne), p. 121, 596
- 1 2 (en) Beugelsdijk DCM et al., « A phylogenetic study of Boletus section Boletus in Europe », Persoonia, vol. 20, no 6, , p. 971–981 (DOI 10.3767/003158508X283692)
- ↑ Singer, Rolf, The Agaricales in modem taxonomy" 4e édition, Sven Koeltz Scientific Books, Germany1986
- ↑ (en) Rolf Singer, The Agaricales in Modern Taxonomy, vol. 1, Koenigstein, Koenigstein Königstein im Taunus, Germany: Koeltz Scientific Books, , 4e éd., 981 p., 73 pls. (ISBN 3-87429-254-1), p. 779
- ↑ (en) Smith AH, Thiers HD., The Boletes of Michigan, Ann Arbor, Michigan, University of Michigan Press, (lire en ligne), p. 221
- ↑ (en) Binder M, Hibbett DS., « Molecular systematics and biological diversification of Boletales », Mycologia, vol. 98, no 6, , p. 971–981 (PMID 17486973, DOI 10.3852/mycologia.98.6.971, lire en ligne [PDF], consulté le )
- ↑ (en) BTM Dentinger et al., « Molecular phylogenetics of porcini mushrooms (Boletus section Boletus) », Molecular Phylogenetics and Evolution, vol. 57, no 3, , p. 1276–1292 (lire en ligne)
- ↑ Taylor J.W., Berbee M.L., 2006. Dating divergences in the Fungal Tree of Life: review and new analyses. Mycologia 98, 838.
- ↑ Lucking, R., Huhndorf, S., Pfister, D.H., Plata, E.R., Lumbsch, H.T., Fungi evolved right on track. Mycologia 101, 810; 2009
- ↑ Dentinger B.T.M. et al., 2010, ibidem
- 1 2 Arora D. (2008). California Porcini: Three New Taxa, Observations on Their Harvest, and the Tragedy of No Commons. Economic Botany 62(3): 356-375
- ↑ (en) I. R. Hall, A. J. E. Lyon, Y. Wang & L. Sinclair, « Ectomycorrhizal fungi with edible fruiting bodies 2. Boletus edulis », Economic Botany, vol. 52, no 1, , p. 44–56 (DOI 10.1007/BF02861294).
- ↑ Giuseppe Pace, Mushrooms of the World, Firefly Books, , p. 122
- ↑ Les différents stades de la pousse d'un cèpe
- ↑ (en) D.C.M. Beugelsdijk, S. van der Linde, G.C. Zuccarello et H.C. den Bakker, « A phylogenetic study of Boletus section Boletus in Europe », Persoonia - Molecular Phylogeny and Evolution of Fungi, vol. 20, no 1, , p. 1–7 (ISSN 0031-5850, PMID 20467482, PMCID PMC2865352, DOI 10.3767/003158508x283692, lire en ligne, consulté le )
- ↑ Arora D. & Simonini, Economic Botany 62(3): 374. 2008
- ↑ Smith A.H. & Thiers, in Persoonia 7(2): 366, 1973.
- ↑ Jerzy Falandysz, Aneta Frankowska, Grazyna Jarzynska et Anna Dryzałowska, « Survey on composition and bioconcentration potential of 12 metallic elements in King Bolete (Boletus edulis) mushroom that emerged at 11 spatially distant sites », Journal of Environmental Science and Health, Part B, vol. 46, no 3, , p. 231–246 (ISSN 0360-1234, DOI 10.1080/03601234.2011.540528, lire en ligne, consulté le )
- ↑ J. Alonso, M. J. Salgado, M. A. García et M. J. Melgar, « Accumulation of Mercury in Edible Macrofungi: Influence of Some Factors », Archives of Environmental Contamination and Toxicology, vol. 38, no 2, , p. 158–162 (ISSN 0090-4341 et 1432-0703, DOI 10.1007/s002449910020, lire en ligne, consulté le )
- ↑ Július Árvay, Ján Tomáš, Martin Hauptvogl et Miriama Kopernická, « Contamination of wild-grown edible mushrooms by heavy metals in a former mercury-mining area », Journal of Environmental Science and Health, Part B, vol. 49, no 11, , p. 815–827 (ISSN 0360-1234 et 1532-4109, DOI 10.1080/03601234.2014.938550, lire en ligne, consulté le )
- ↑ R. Bargagli et F. Baldi, « Mercury and methyl mercury in higher fungi and their relation with the substrata in a cinnabar mining area », Chemosphere, vol. 13, no 9, , p. 1059–1071 (ISSN 0045-6535, DOI 10.1016/0045-6535(84)90065-1, lire en ligne, consulté le )
- ↑ Chang S-T, Miles PG., (2004). Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact (2d ed.). Boca Raton, Florida: CRC Press. p. 131.
- ↑ Arora D., (1986). Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi. Berkeley, California: Ten Speed Press. p. 30.
- ↑ Veselkov IM. (1975). "Artificial propagation of Boletus edulis in forests". Растительньіе Ресурсы России 11: 574–78.
- ↑ Ceruti A, Tozzi M, Reitano G. (1988). "Micorize di sintesi tra Boletus edulis, Pinus sylvestris e Picea excelsa" (in Italian). Allionia (Turin) 28: 117–24.
- ↑ Fitter AH, Garbaye J.: "Interaction between mycorrhizal fungi and other organisms". In Robson AD, Abbott LK, Malajczuk N. Management of Mycorrhizas in Agriculture, Horticulture and Forestry. Dordrecht, Netherlands: Kluwer Academic Publishers. 1994
- ↑ Garbaye et al. ibidem, p. 110
- ↑ Olivier JP, Guinberteau J., Rondet J. Mamoun M.: Vers l'inoculation contrôlée des cèpes et bolets comestibles, Rev. For. Fr. XLIX -n° sp. 1997
- ↑ Bon Marcel Champignons de France et d'Europe occidentale (Flammarion, 2004)
- ↑ Philips Roger : Les champignons (Solar, 1981)
- ↑ Courtecuisse Régis, Duhem Bernard: Guide des champignons de France et d'Europe (Delachaux & Niestlé, 1994-2000).
- ↑ Larrère et de La Soudière, Cueillir la montagne. Lyon: La Manufacture Ed., 1987. 254 pp.
- ↑ Poitou N, Mamoun M., Delmas J. Cultures mycéliennes de champignons mycorhiziens en milieu optimal. , in Mushroom Science, vol. 11, p. 885-892,1981
- ↑ Poitou N., Mamoun M., Ducamp M., Delmas J., Premiers résultats de fructification in situ d’un bolet mycorhizien comestible en conditions contrôlées, inPHM Revue horticole, vol. 239, p. 45-47,1983
- ↑ Ouzouni PK, Riganakos KA., "Nutritional value and metal content profile of Greek wild edible fungi". Acta Alimentaria 36 (1): 99–110. 2006
- ↑ Crisan EV, Sands A, "Nutritional value". In Chang ST, Hayes WA. The Biology and Cultivation of Edible Mushrooms. New York: Academic Press. p. 727–93. 1978
- ↑ Source: USDA Nutrient Database
- ↑ Ouzouni PK, Riganakos KA., "Nutritional value and metal content profile of Greek wild edible fungi". Acta Alimentaria 36 (1): 99–110., 2006
- ↑ Kalač P., "Chemical composition and nutritional value of European species of wild growing mushrooms: a review". Food Chemistry 113 (1): 9–16, 2009
- ↑ Pedneault K, Angers P, Gosselin A, Tweddell RJ., "Fatty acid composition of lipids from mushrooms belonging to the family Boletaceae". Mycological Research 110 (Pt 10): 1179–83. 2006
- ↑ Ribeiro B, Andrade PB, Silva BM, Baptista P, Seabra RM, Valento P., "Comparative study on free amino acid composition of wild edible mushroom species". Journal of Agricultural and Food Chemistry 56 (22): 10973–79. 2008
- ↑ Tsai S-Y, Tsai H-L, Mau J-L., "Non-volatile taste components of Agaricus blazei, Agrocybe cylindracea and Boletus edulis". Food Chemistry 107 (3): 977–83, 2008
- ↑ Kalač P., "Chemical composition and nutritional value of European species of wild growing mushrooms: a review". Food Chemistry 113 (1): 9–16. 2009
- ↑ Falandysz J., "Selenium in edible mushrooms". Journal of Environmental Science and Health Part C–Environmental Carcinogenesis & Ecotoxicology Reviews 26 (3): 256–99, 2008
- ↑ Mutanen M., "Bioavailability of selenium in mushrooms". International Journal for Vitamin and Nutrition Research 56 (3): 297–301., 1986
- 1 2 Mattila P, Lampi A-M, Ronkainen R, Toivo J, Piironen V., "Sterol and vitamin D2 contents in some wild and cultivated mushrooms". Food Chemistry 76 (3): 293–98. 2002
- ↑ Kalač P.. ibidem 2009
- ↑ Krzyczkowskia W, Malinowskaa E, Suchockib P, Klepsa J, Olejnikd M, Herold F., "Isolation and quantitative determination of ergosterol peroxide in various edible mushroom species". Food Chemistry 113 (1): 351–55, 2008
- ↑ (en) Christian Collin-Hansen, Sindre Sindre A. Pedersen,, Rolf A. Andersen et Eiliv Steinnes, « First report of phytochelatins in a mushroom: induction of phytochelatins by metal exposure in Boletus edulis », Mycologia, vol. 99, no 2, , p. 161-174 (lire en ligne)
- ↑ Zheng S, Li C, Ng TB, Wang HX., "A lectin with mitogenic activity from the edible wild mushroom Boletus edulis". Process Biochemistry 42 (12): 1620–24, 2007
- ↑ Kandefer-Szersen M, Kawecki Z, Salata B, Witek M., "Mushrooms as a source of substances with antiviral activity" (in Polish). Acta Mycologica 16 (2): 215–20, 1980
- ↑ Li D, Zhao W-H, Kong B-H, Ye M, Chen H-R., "Inhibition effects of the extract and polysaccharide in macrofungus on TMV". Journal of Yunnan Agricultural University 24 (2): 175–80, 2009
- ↑ Piraino FF., "Emerging antiviral drugs from medicinal mushrooms". International Journal of Medicinal Mushrooms 8 (2): 101–14, 2006
- ↑ Ribeiroa B, Lopesa R, Andradea PB, Seabraa RM, Gonçalvesa, Baptistab P, Quelhasa I, Valentão P., "Comparative study of phytochemicals and antioxidant potential of wild edible mushroom caps and stipes". Food Chemistry 110 (1): 47–56. 2008
- ↑ Ey J, Schömig E, Taubert D., "Dietary sources and antioxidant effects of ergothioneine". Journal of Agricultural and Food Chemistry 55 (16), 2007
- ↑ Lucas EH, Montesono R, Pepper MS, Hafner M, Sablon E., "Tumor inhibitors in Boletus edulis and other holobasidiomycetes". Antibiotics and Chemotherapy 7: 1–4.
- ↑ Lamaison J-L, Polese J-M., The Great Encyclopedia of Mushrooms. Koln, Germany: Könemann. p. 28. (1957) 2005
- ↑ un total de 4 cas sur 10 ans à hôpital universitaire de Zurich et à l'hôpital Karolinska de Stockholm
- ↑ Torricelli R., Johansson S. G. O., Wiithrich B., Ingestive and inhalative allergy to the mushroom "Boletus edulis", Allergy, Volume 52, Issue 7, pages 747–751, July 1997
- ↑ Dr Giacomoni L., Nicot J., Pautot S., Redeuihl G., Branchu G., Hartog D., Herubel A., Marxmuller H., Millot U. et Schaeffner C.