
| Empire | Eukaryota |
|---|---|
| Super-règne | Opisthokonta |
Bartl., 1830
- Mycota Alexop., 1962
_white_spores_a_-_panoramio.jpg.webp)

Les Fonges[1] (Fungi)[2], également appelés Mycètes (Mycota)[3], sont un règne d'organismes eucaryotes appelés communément champignons. Ce règne est diversifié, regroupant des organismes microscopiques, invisibles à l'œil nu, microchampignons unicellulaires (levures) ou pluricellulaires (moisissures), jusqu'aux « champignons supérieurs »[4] dotés le plus souvent d'un pied et d'un chapeau, que le promeneur récolte.
Leur succès évolutif est en grande partie dû à leur plasticité génétique associée à des caractéristiques biologiques extrêmement diversifiées, depuis leurs modes de vie (symbiose de type lichens ou mycorhizes, parasitisme biotrophe ou nécrotrophe, saprotrophie) jusqu'à leur développement qui implique divers processus : la sporulation, la germination des spores, la croissance de l'appareil végétatif (mycélium composé d'hyphes), et la reproduction par voie asexuée ou sexuée en différenciant un appareil reproducteur, le sporophore.
L'existence simultanée d’une paroi cellulaire périphérique et de vacuoles turgescentes dans le cytoplasme, les rapproche des végétaux auxquels on les rattachait autrefois, alors que leur corps végétatif non différencié et leur paroi peptido-polyosidique les distingue des plantes. L'absence de chloroplastes, de chlorophylle et d'amidon en fait, comme les animaux, des organismes hétérotrophes au carbone. Sur la base de ces caractères particuliers, l'Américain Robert Harding Whittaker classe en 1959 les champignons dans un règne à part, celui des Mycota ou Mycètes. D'après des substances typiques du règne animal retrouvées chez les champignons lors d'études chimiotaxonomiques (chitine, mélanine, bufoténine, etc.) et l'analyse des séquences ADN, la classification phylogénétique actuelle les rend plus proches des animaux, formant avec eux l'essentiel du super-règne des Opisthochontes.
La mycologie est la science qui les étudie.
Environ 120 000 espèces de Mycètes ont été décrites à ce jour (avec un rythme moyen de 1 300 espèces nouvelles décrites annuellement)[5], mais la majorité sont des microorganismes incultivés et les estimations des mycologues de leur nombre total, basées notamment sur des analyses métagénomiques d'un grand éventail de sols, vont de 0,5 à 10 millions, l'établissement de l'inventaire fongique qui se voudrait exhaustif prenant plusieurs milliers d'années au rythme de description actuelle[6],[7],[5].
Étymologie
Il existe une grande dispersion étymologique pour désigner les champignons, ce qui peut laisser penser que les hommes pré- et proto-historiques consommaient rarement ces organismes[8]. Cependant, la découverte en 1991 d'Ötzi révèle que cet homme vivant vers 2500 av. J.-C. transportait dans son sac deux types de champignons, des polypores du bouleau, probablement à usage médicinal (consommés pour ses propriétés antibiotiques, vermifuges et vulnéraires) et de l'amadou, probable allume-feu, ce qui suggère que les hommes préhistoriques qui vivaient de chasse et de cueillette, ont récolté des champignons pour leur consommation, comme le font encore de nos jours maintes peuplades exploitant la nature[9].
Le terme champignon vient de l'ancien français du XIIIe siècle champignuel (par substitution du suffixe -on*) du bas latin campinolius[10] « petits produits des campagnes » ou « qui pousse dans les champs » (dérivé en -ŏlu de campania, « campagne, champ »), lui-même issu de la racine latine campus, « campagne », qui donne le champ[alpha 1], la plaine.
Le mousseron (nom vernaculaire du Tricholome de la Saint-Georges), perçu comme poussant dans la mousse, a donné en anglais le nom générique du champignon, mushroom. La racine de ce mot semble être la mousse, mais est plus probablement l'indo-européen *meu qui l’apparente au latin muscus (« mousse »), mucus (« morve »), mucor (« moisissure ») et au grec mykès (d'où les Mycètes) désignant d'abord les champignons en général[11]. Les termes grec et latin sont ainsi une allusion possible aux champignons qui se protègent contre la dessication par une couche de mucus qui recouvre leur chapeau et parfois aussi leur pied, ou à la mycophobie ancestrale, les champignons étant associés aux mucosités nasales repoussantes[11].
Selon une étymologie populaire, fungus et fongus sont la contraction du latin funus, « funérailles » et d'ago, « produire », rappelant les nombreux décès provoqués par les champignons toxiques[12]. Une origine plus probable de ce terme serait une allusion à l'aspect poreux ou spongieux des champignons : les mots espagnol (hongo) et italien (fungo) remontent en effet à une racine méditerranéenne[alpha 2], *sfong-/*fung-, qui a donné en grec spongos et en anglais sponge, signifiant « éponge », et en latin fungus qui signifie en même temps « champignon » et « éponge »[14].
Définition mycologique

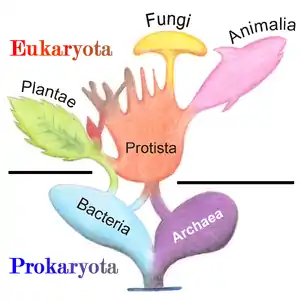
Parmi les Eucaryotes, les Mycètes ne sont ni des plantes (puisqu'ils n'effectuent pas de photosynthèse) ni des animaux (bien qu'ils soient, comme ces derniers, des Opisthocontes), mais forment un règne à part entière. Autrefois classés avec les algues dans les végétaux « sans rameaux feuillés » : « cryptogames thallophytes non-chlorophylliens », les Mycètes constituent à présent un règne autonome, le cinquième règne ou « règne fongique » (du latin fungus = champignon).
Selon la classification phylogénétique, qui affine encore plus les liens de parenté, ce règne des Fungi est rattaché aux Unikonta, une des deux divisions des Eukaryota, et plus précisément aux Opisthokonta (ce qui signifie qu'ils sont plus proches des animaux, Opisthocontes également, que des plantes, Bikontes) : les champignons ont originellement des cellules avec un flagelle postérieur mais ont perdu ce flagelle à plusieurs reprises au cours de l'évolution[15].
Comparée à celle des végétaux, la définition de l'organisme fongique est d'abord négative : dépourvus de tiges, de feuilles et de racines. Il est formé d'un appareil végétatif appelé thalle, sans tissus fonctionnels ni organes différenciés, constitué de cellules végétatives allongées et cloisonnées nommées hyphes. Ces hyphes s'associent le plus souvent en mycélium, sorte de feutrage difficile à voir à l'œil nu et le plus souvent impossible à identifier en l'état. Parfois, le thalle est un simple tube sans cloisons ; on parle alors de structure cœnocytique et de siphon.
Leur reproduction est très discrète et d'apparence capricieuse, tantôt asexuée, tantôt sexuée, au moyen de cellules spéciales, les spores. Le champignon ne produisant pas de fleurs, il ne peut être un fruit ou carpophore au sens botanique, aussi l'appareil de « fructification » portant les spores et permettant la reproduction est aujourd'hui désigné par le terme de « sporophore ». La dispersion des spores est assurée par divers mécanismes. La majorité des champignons utilise l'anémochorie (dissémination par le vent[16]). Les autres modes de dissémination sont l'hydrochorie (par les éclaboussures de la pluie)[17], la barochorie (par la seule gravité) et la zoochorie (par les animaux : endozoochorie des limaces[18], ectozoochorie et endozoochorie[19] des mammifères[20], des insectes mycophages qui sont attirés par les belles couleurs du chapeau[21] ou les odeurs)[22].
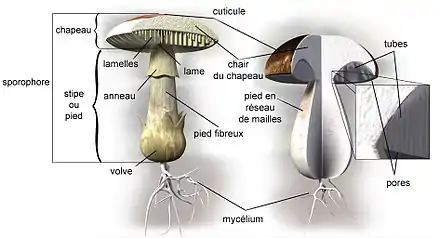
Chez les « champignons supérieurs » (30 000 espèces de macromycètes dont le sporophore est visible à l'œil nu[23], dont plus de 15 000 espèces connues en France[24]), cet appareil (souvent constitué d'un pied et d'un chapeau et encore appelé champignon dans la langue courante[25]) est particulièrement développé. Le reste du champignon (le mycélium) correspondant à sa forme trophique est souterrain ou dans le cœur du bois ou de l'hôte animal et donc invisible. Les champignons « inférieurs » peuvent aussi produire des sporophores, mais ceux-ci demeurent microscopiques.
La plupart des champignons ont une structure multicellulaire, mais il y a des exceptions notables : ainsi les levures sont unicellulaires.
Définition du règne fongique
Les organismes du règne des Fungi présentent les caractéristiques suivantes :
- ils sont eucaryotes (organismes possédant des cellules et dont les chromosomes sont enfermés dans un noyau) ;
- ils sont hétérotrophes vis-à-vis du carbone, qu'ils doivent donc trouver dans leur environnement immédiat. Incapables d'utiliser l'énergie solaire, ils absorbent de nombreuses molécules carbonées fabriquées par d'autres êtres vivants ;
- ils sont absorbotrophes, se nourrissant par absorption (décomposition) et non par ingestion (caractère animal). Ce mode d'alimentation implique la sécrétion d'enzymes extracellulaires (en) (ces exoenzymes étant des enzymes hydrolytiques libérées par exocytose) assurant une digestion extracellulaire (en) (exodigestion)[26],[27]. Dépourvus de racines, tiges et feuilles, leur appareil végétatif, appelé mycélium, est diffus, ramifié et tubulaire, constitué de filaments fins enchevêtrés, les hyphes, à croissance apicale, permettant la nutrition par absorption. Dans la nature, la plupart des plantes ont recours aux mycorhizes, qui est une symbiose entre les racines d'une plante et le mycélium d'un champignon. Les racines de la plante produisent des sucres pour le champignon. Le mycélium procure en retour de l'eau et des sels minéraux inaccessibles aux racines de la plante ;
- comme les animaux, ils sont opistochontes, c'est-à-dire que, comme on peut le voir chez certains champignons primitifs (chytridiomycètes), ils dérivent d'espèces ayant une phase aquatique avec des gamètes à un seul flagelle propulseur (type spermatozoïde). Les groupes modernes sont tous affranchis de la phase aquatique obligatoire et possèdent des spores sans flagelle ;
- l'existence simultanée d'une paroi cellulaire périphérique et de vacuoles turgescentes dans le cytoplasme, rapproche les Fungi des végétaux mais l'absence de chloroplastes en fait, comme les animaux, des organismes hétérotrophes ;
- ils fabriquent des substances qui leur sont propres. Leur paroi contient des marqueurs taxonomiques : hémicelluloses (callose, α et β-glucanes, mannanes), polyosides aminés (chitosane, chitine voisine de la chitine des insectes, caractère animal, alors que les végétaux possèdent une paroi pectocellulosique)[28]. L'ergostérol est la forme majoritaire de stérol retrouvée dans la membrane cellulaire fongique (c'est le cholestérol chez les animaux et les phytostérols chez les végétaux)[29]. Le glycogène est le principal polysaccharide, comme chez les animaux. La synthèse de la lysine est réalisée par la voie de l'α-aminoadipate. D'autres substances typiques du règne animal sont retrouvées chez les champignons (mélanine pour durcir les parois fongiques, bufoténine à l'action toxique, etc.)[30]. Ils produisent une variété de métabolites secondaires spécifiques associés à d'importants mécanismes de défense et de résistance. Ils émettent notamment des composés organiques volatils qui ont des propriétés antibactérienne, antifongique, insecticide, et qui peuvent être un critère important pour leur identification[31] ;
- ils n'ont pas de différences sexuelles. Ils sont capables de produire un nombre considérable de spores haploïdes après une dicaryophase plus ou moins longue entre la plasmogamie et la caryogamie. Les mycologues utilisent plutôt le terme de bipolarité sexuelle que celui de sexualité.
Histoire évolutive

.png.webp)
Les premiers groupes modernes d'organismes eucaryotiques (algues, champignons aquatiques) apparaissent dans l'océan puis dans les eaux douces il y a 1,3 milliard d'années. Des microchampignons aquatiques apparaissent ainsi probablement au fond des océans dans des sources hydrothermales, l'hydromycoflore ayant depuis gardé un mode de vie benthique (tel le genre Tappania (en) apparu il y a 1,6 milliard d'années)[34].
Contrairement aux plantes et aux animaux, les fossiles de champignons mycéliens sont rares car leur matière fragile se prête mal à la fossilisation. Jusqu'à récemment, les fossiles les plus anciens étaient des hyphes fossilisés, spores de Glomeromycota de 420 Ma[35] et un ascocarpe d'Ascomycète de 400 Ma découvert dans la flore de Rhynie. Les « champignons supérieurs » étaient donc bien individualisés dès cette époque[36], ce qui indique une origine des champignons mycéliens beaucoup plus ancienne, sans doute entre 0,76 et 1,06 milliard d'années[37], voire 2,4 milliards d'années mais ces études se basent sur des critères morphologiques (donc ambigus) pour avancer une affinité fongique[38]. Des microfossiles multicellulaires d'affinité fongique ont été découverts en 2019 dans un schiste argileux de la formation de Grassy Bay (Arctique canadien), daté à 1,01–0,89 Ga[39].
Trois hypothèses relatives à la terrestrialisation fongique sont proposées : selon le scénario « vert », les champignons ont coévolué avec les ancêtres des plantes terrestres, peut-être en tant que symbiotes ou parasites biotrophes des algues Streptophytes qui ont conquis des habitats d'eau douce après leur séparation d'anciennes algues vertes chlorophytes. Le scénario « brun » suppose que les champignons zoosporiques (spores flagellées, caractère qui semble restreindre ces organismes aux milieux aquatiques et dans les sols humides) ont colonisé des sédiments ou des terres humides grâce à un mode de vie saprotrophe (bactéries et algues en décomposition échoués sur la côte ou les berges de plans d'eau, de rivières), puis ont perdu leur flagelle et ont développé une croissance hyphale. La voie « blanche » suggère que les champignons zoosporiques se sont adaptés à des environnements gelés qui ont servi de transition entre les milieux aquatiques et terrestres[40].
Selon le mycologue Paul Stamets, c'est la conquête des terres par les champignons à mycélium avant l'arrivée des plantes terrestres qui a permis à ces dernières de se développer il y a plus de 500 millions d'années. Une étude de 2020 identifie le plus ancien fossile présumé dans un schiste dolomitique du supergroupe Mbuji-Mayi en République démocratique du Congo. Ce microfossile, daté d'environ 715 à 810 millions d'années, serait apparu dans une zone de transition entre milieux aquatique et terrestre[38],[41].
Alors que les organismes vivant dans l'eau baignent dans une solution d'éléments nutritifs, ceux ayant conquis les terres (analogues à des déserts minéraux) ont développé des stratégies adaptatives pour leur alimentation et hydrique et minérale. Le mycélium fongique contient des acides oxaliques et des exoenzymes extracellulaires qui auraient altéré les roches des premiers sols qui devaient sans doute se présenter comme des croûtes semblables à celle des déserts chauds ou froids actuels. Les champignons, associés à des bactéries ou à des algues dans les lichens (hypothèses que soulèvent le fossile Prototaxites), auraient ainsi été très actifs dans la colonisation des roches, la minéralisation des sols et la pédogenèse, facilitant l'alimentation minérale des plantes[42].
Selon une théorie[43], les champignons symbiotiques de type lichen sont parmi les premiers organismes à avoir colonisé la terre durant le Cambrien et l'évolution des plantes terrestres au cours du Dévonien n'aurait ainsi pas été possible sans eux. La conquête terrestre nécessitant plusieurs adaptations simultanées, l'établissement de symbioses a permis d'apporter des solutions et d'effectuer ce saut macroévolutif afin de coloniser ce milieu plus hostile par les plantes. Des algues ont ainsi probablement établi des associations mutualistiques avec des microchampignons[44] (mycophycobioses, lichens). Par provocation, il est tentant d'écrire selon Marc-André Selosse, que la majorité des plantes terrestres sont des sortes de champignons lichénisés dans lesquels l'algue occupe la partie aérienne et visible de l'association, et le champignon pluricellulaire la partie souterraine[45]. Selon une autre hypothèse, ce sont les endomycorhizes à arbuscules primitives qui ont permis aux plantes de sortir de l'eau et de s'adapter au milieu terrestre[46].
Sur les 100 000 espèces de champignons répertoriées en 2015, « près de 10 000 produisent des fructifications à l'œil nu, un peu plus de 1 100 sont comestibles et consommés comme aliments, et environ 500 sont utilisés comme remèdes dans la médecine traditionnelle de tous les pays en développement »[47].
| Opisthokonta 1300 Ma |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Classification

Classifications classiques (morphologiques)
Historique
Les champignons ont été classés dans le passé comme faisant partie du règne végétal du fait de la présence d'une paroi cellulaire et de plusieurs similitudes entre leurs cycles de reproduction et ceux des algues. L'idée de classer les champignons dans un règne à part est émise dès 1783 par Noël Martin Joseph de Necker dans son Traité sur la mycitologie[48]. Mais ce n'est qu'en 1959, que l'écologue américain Robert Whittaker les classe dans un règne à part, celui des Mycota, sur la base de plusieurs caractères particuliers comme l'absence de chlorophylle et d'amidon[49].
Une des classifications les plus répandues est celle de Geoffrey Clough Ainsworth (1905-1998) et Guy Richard Bisby (1889-1958) dans leur Dictionary of Fungi (1971), bien qu'elle soit aujourd'hui profondément remaniée (9e édition en 2001), on trouve encore les anciennes versions de cette classification dans certains ouvrages. Elle suit manifestement les recommandations des études phylogénétiques actuelles.
La classification phylogénétique et la classification traditionnelle ont fait l'objet d'une synthèse qui reprend les divisions classiques phylum, sous-phylum, sur-phylum, classe, sous-classe et ordre. Ce document de 2007, cosigné par 67 auteurs reste une référence[50].
Ancienne classification d'Ainsworth
Exemple d'une ancienne classification morphologique[51] :
- Règne des Fungi
- Division des Myxomycota (présentent des plasmodes)
- Acrasiomycètes
- Myxomycètes
- Plasmodiophoromycètes
- Division des Eumycota (ne présentent pas de plasmode)
- Subdivision des Mastigomycotina (présentent des spores mobiles -zoospores-)
- Chytridiomycètes
- Hyphochytridiomycètes
- Oomycètes
- Subdivision des Deuteromycotina
- Subdivision des Zygomycotina
- Subdivision des Ascomycotina
- Subdivision des Basidiomycotina
- Subdivision des Mastigomycotina (présentent des spores mobiles -zoospores-)
- Division des Myxomycota (présentent des plasmodes)
Ce premier règne des champignons comprenait un certain nombre d'organismes qui, par la suite, ont été replacés dans d'autres règnes :
- les Oomycètes et les Hyphochytridiomycètes qui sont maintenant classés dans les Straménopiles ;
- les Myxomycota qui sont maintenant classés dans plusieurs groupes de protistes.
Les Chytridiomycota ou Chytridiomycètes sont des espèces dont les spores portent un flagelle. On les considère comme les ancêtres de tous les autres champignons.
Les Ascomycota ou Ascomycètes ont des spores qui sont produites à l'intérieur de sacs (les asques) et sont projetées, à maturité, à l'extérieur par ouverture de l'asque.
Les Basidiomycota ou Basidiomycètes ont des spores qui se développent à l'extrémité de cellules spécialisées (les basides) et sont dispersées par le vent à maturité.
Les Glomeromycota ou Glomeromycètes étaient autrefois classés dans les Zycomycota. Ils sont maintenant considérés comme constituant une division à part.
Les Deuteromycota ou Deuteromycètes formaient le groupe des champignons imparfaits, il s’agissait de champignons que l'on ne connaissait que sous forme anamorphe, il est maintenant en général possible de rattacher chaque genre à diverses classes d'ascomycètes.
La classification de Geoffrey Clough Ainsworth (division des Fungi en Eumycota et Myxomycota) est aujourd'hui remplacée par le groupe Eumycota (« champignons vrais » proches génétiquement des animaux) et le groupe Pseudomycota (« faux champignons » apparentés à des algues donc aux plantes)[52].
Liste des embranchements
Selon Catalogue of Life (6 novembre 2020)[53] :
- Ascomycota ;
- Basidiomycota ;
- Blastocladiomycota ;
- Chytridiomycota ;
- Entomophthoromycota ;
- Glomeromycota ;
- Zygomycota.
 Morchella conica (Ascomycota)
Morchella conica (Ascomycota)
.jpg.webp) Physoderma pulposum (Blastocladiomycota)
Physoderma pulposum (Blastocladiomycota) Cladochytrium menyanthis (Chytridiomycota)
Cladochytrium menyanthis (Chytridiomycota) Rozella allomycis (Cryptomycota)
Rozella allomycis (Cryptomycota)_Fresen_204248.jpg.webp) Entomophthora muscae (Entomophthoromycota)
Entomophthora muscae (Entomophthoromycota) Gigaspora margarita (Glomeromycota)
Gigaspora margarita (Glomeromycota) Monoblepharis polymorpha (Monoblepharidomycota)
Monoblepharis polymorpha (Monoblepharidomycota)_Figure_1.jpg.webp) Buwchfawromyces eastonii (Neocallimastigomycota)
Buwchfawromyces eastonii (Neocallimastigomycota) Spinellus fusiger (Zygomycota, parasite)
Spinellus fusiger (Zygomycota, parasite)
Classification phylogénétique (cladistique)
Les premières études de portions d'ADN et de chromosomes tendent à proposer une nouvelle classification, dite « classification systématique des champignons » (terme confus car déjà utilisé pour classification systématique classique), et coïncide de plus en plus avec la classification phylogénétique, donc de moins en moins avec la classification morphologique.
- Fungi
- Microsporidia
- clade
- Kickxellomycotina
- Zoopagomycotina
- Entomophthoromycotina
- Blastocladiomycota
- Mucoromycotina
- Neocallimastigomycota
- Chytridiomycota
- Symbiomycota
- Glomeromycota
- Symbiomycota
- Dikarya
Le cladogramme qui suit est basé sur les travaux de Philippe Silar[54], « The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research »[55] et Tedersoo et al. 2018[56].
| Cladogramme détaillé de la phylogénie des Mycètes. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Stratégies écologiques pour l'obtention de nutriments

Grâce au métabolisme de la photosynthèse, les végétaux verts peuvent fixer directement le gaz carbonique de l'air : on dit qu'ils sont autotrophes. Ce n'est pas le cas des champignons, qui sont hétérotrophes : ils doivent trouver le carbone nécessaire à leur vie dans leur environnement immédiat, sous la forme de matières organiques. Ils se nourrissent à partir de substances dissoutes par absorbotrophie selon différentes stratégies écologiques, la saprotrophie, la biotrophie mutualiste ou parasitaire (le mycologue et microbiologiste Francis Martin les associe aux trois grandes guildes d'espèces fongiques des bois, les mutualistes ou symbiotes, les décomposeurs ou fossoyeurs, et les parasites, surnommés « Le Bon, la Brute et le Truand »)[59], le commensalisme et la carnivorie.
- Saprotrophie : les champignons peuvent se nourrir de matière organique morte ou en décomposition (feuilles mortes, débris végétaux ou animaux, excréments) : on les appelle alors des « saprotrophes » (et également « coprophiles » pour ceux qui croissent sur les excréments). On les trouve notamment en forêt, là où cette nourriture, sous forme d'humus, existe en grande quantité (champignons corticioïdes et polyporoïdes, décomposeurs de la litière, agents de dégradation du bois responsables de leur pourriture). En dégradant ainsi la matière organique morte, les champignons saprophytes remettent à la disposition des autres organismes des éléments minéraux essentiels de nouveau assimilables (azote, phosphore, carbone). Les moisissures sont également des champignons saprotrophes décomposeurs de matière organique. Ils participent ainsi au recyclage de la matière organique.
- Symbiose mutualiste : les champignons peuvent vivre en symbiose avec d'autres êtres vivants autotrophes, au point que l'un ne peut vivre sans l'autre (endosymbiose extracellulaire comme les champignons endophytes, les lichens et les associations mycorhiziennes). Ainsi, les lichens sont des associations de champignons (essentiellement des Ascomycètes, mais aussi quelques Basidiomycètes) et de cyanobactéries ou d'algues vertes. Le champignon fournit à l'algue protection, eau et sels minéraux et, en retour, celle-ci l'approvisionne en glucides, produits de la photosynthèse. Mais la plupart des plantes vertes vivent également en symbiose avec des champignons du sol, formant une association symbiotique appelée mycorhize. Il existe des cas de symbiose avec des animaux : les champignons aident ainsi fourmis et termites à digérer la cellulose.
- Parasitisme : les champignons peuvent également tirer parti de la matière organique vivante. Ils sont parasites et vivent aux dépens d'un être vivant à leur propre compte (biotrophie parasitaire). Souvent pathogènes, ils provoquent des maladies et entraînent parfois la mort de leurs hôtes (mycoparasites nécrotrophes d'autres champignons, parasites des algues, champignons zoopathogènes et phytopathogènes). Les anthracnoses, les oïdiums[60] sont des maladies cryptogamiques des végétaux. Chez l'humain et les animaux, des mycoses comme les dermatophytose dues à Trichophyton, les candidoses dues aux levures Candida, les aspergilloses dues aux champignons du genre Aspergillus, les cryptococcoses de Cryptococcus, la teigne, le muguet, la pneumonie, etc. sont des maladies dues à de tels champignons parasites. De nombreux parasites sont biotrophes au départ, puis nécrotrophes (notion d'agent pathogène hémibiotrophe).
- Commensalisme : Un champignon est dit « commensal » s'il tire profit de son hôte sans nuire à ce dernier (il s'en sert par exemple comme d'un support), mais sans non plus lui apporter d'avantage. Exemples : commensalisme cutané de levures du genreMalassezia ; commensalisme digestif, respiratoire et digestif de levures du genre Candida. Il y a un continuum dynamique entre le commensalisme et le parasitisme. Si cet équilibre dynamique est rompu, ces levures commensales peuvent devenir parasites et induire des pathologies.
- Carnivorie : 140 espèces de champignons (Arthrobotrys, pleurote en huître) sont carnivores, déployant des pièges (filament à mucus collant, cils, hyphes formant un lasso, phéromones sexuelles attitrant la proie) pour capturer protistes, amibes et vers (nématodes, rotifères)[61].
Développement des mycètes

Le développement des mycètes est caractérisé par divers processus : la sporulation, la germination des spores, la croissance de l'appareil végétatif (thalle composé d'hyphes), et la reproduction par voie asexuée ou sexuée en différenciant un appareil reproducteur, le sporophore.
Habitats
Bien qu’ils passent souvent inaperçus, les champignons sont présents dans presque tous les compartiments de l'environnement terrestre, y compris au sein d'organismes vivant avec lesquels ils peuvent entretenir des interactions durables dont le parasitisme n'est qu'une des formes. Mais leur activité est le plus souvent aérobie (certains comme de nombreuses levures peuvent facultativement vivre en anaérobie). Ils sont donc plus rares dans les eaux douces ou salées, dans les couches abiotiques de la croûte terrestre et en haute altitude.
Le vocabulaire mycologique concernant l'habitat des champignons est riche ; en voici un petit florilège[62] :
- Abiéticole : se développe dans les sapinières
- Betulicole : se développe dans les bétulaies, tel Boletus betulicola
- Caducicole : se développe sur les arbres à feuilles caduques
- Calcicole : se développe préférentiellement sur un substrat calcaire
- Carbonicole : se développe sur les débris carbonisés (charbonnières)
- Coprophile : se développe sur des excréments animaux
- Corticole : se développe sur les écorces
- Graminicole : se développe au milieu ou sur les graminées
- Fimicole : se développe sur le fumier ou les endroits fumés
- Fongicole : se développe sur un autre champignon qu'il parasite
- Humicole : se développe sur l'humus
- Lignicole : se développe sur le bois
- Muscicole : se développe sur les mousses
- Paludicole : se développe dans les marécages
- Pinicole : se développe dans les pinèdes, tel Boletus pinicola
- Quercicole : se développe dans les chênaies, tel Boletus quercicola
- Ramicole : se développe sur les rameaux
- Rudéral : se développe dans les milieux transformés par les activités humaines (surtout enrichis en azote)
- Stercoral : se développe sur les excréments
- Silicole : se développe sur le sol siliceux
- Terricole : se développe sur la terre
- Thermophile : se développe dans les endroits chauds
- Ubiquiste : se développe dans des endroits variés d'un point de vue géographique et écologique

.jpg.webp) Conocybe pubescens, espèce coprophile sur crottin de cheval.
Conocybe pubescens, espèce coprophile sur crottin de cheval.
Coloration
La coloration des champignons et des spores est due à des pigments fongiques. Ces pigments sont des métabolites secondaires classés habituellement autour des voies métaboliques dont ils proviennent : la majorité sont des quinones qui donnent des pigments orangés, rouges ou bruns, ou des dérivés de l'acide pulvinique donnant des pigments jaune orange (principalement dans les champignons de l'ordre des Bolétales). Les caroténoïdes et les pigments azotés (bétalaïnes, alcaloïdes) sont également présents chez les macromycètes mais sont moins fréquents que chez les plantes à fleurs[63],[64].
Ces pigments constituent un élément important de la stratégie écologique pour lutter contre les stress biotiques et abiotiques : rôle de photoprotection contre les UV, couleur plus ou moins vive avertissant les mycophages de la toxicité d'un champignon (théorie du signal honnête et aposématique) ou au contraire attirant des vecteurs biologiques qui favorisent la dispersion active et spécifique des spores, coloration cryptique ou de camouflage, défense chimique contre les herbivores et contre les pathogènes, protection contre des polluants bioaccumulés[65]…
Les champignons présentent une plasticité thermique. Cette plasticité adaptative est utilisée par les macromycètes ectothermes qui produisent dans les climats plus froids la mélanine. Ce pigment brun qui les rend plus sombres leur permet, en absorbant les rayons solaires, de se réchauffer plus vite. De plus, les champignons ectomycorrhiziens sont en moyenne plus sombres que les champignons saprotrophes. Une interprétation est que les premiers ont un accès facilité au carbone (leurs hôtes leur en fournissent une partie) pour la production de pigments coûteuse en énergie, tandis que les seconds investissent surtout leur énergie dans la production de complexes enzymatiques destinés à décomposer le bois mais. Les saprotrophes présentent cependant un léger mélanisme thermique saisonnier (ils sont ainsi plus sombres lors des saisons froides, et plus clairs lors des saisons chaudes)[66].
- Les bétalaïnes sont responsables de la couleur rouge des sporophores d'amanite tue-mouches.
%252C_white_spores%252C_causes_brownrot_at_Schaarsbergen_-_panoramio.jpg.webp) Les couleurs claires du Polypore du bouleau sont en lien avec son mode de vie saprotrophe.
Les couleurs claires du Polypore du bouleau sont en lien avec son mode de vie saprotrophe. Trompette de la mort, champignon ectomycorrhizien encore plus sombre par temps humide[67].
Trompette de la mort, champignon ectomycorrhizien encore plus sombre par temps humide[67].
Rôle dans l'altération météorique des roches

Les champignons sont des agents météoriques qui altèrent la structure superficielle et la composition chimique des roches[alpha 3] et de leurs minéraux. Ils contribuent au développement des sols et aux cycles géochimiques globaux[68].
- Les champignons libres (non symbiotiques) trouvent à la surface des roches des nutriments d'origine variée : poussières transportées par le vent, produits émis par l'industrie et les habitations, et exsudats des micro-organismes et des insectes ou autres animaux.
- Les lichens, organismes symbiotiques associant un champignon et une algue et/ou une cyanobactérie, pratiquent la photosynthèse donc ne dépendent pas de la présence préalable de nutriments : ils sont généralement les premiers à coloniser les roches nouvellement formées, et occupent une fraction notable de la surface terrestre. Plusieurs milliers d'espèces de lichens sont concernées.
- Les champignons microcoloniaux, symbiotiques aussi à l'occasion, forment un troisième groupe de champignons saxicoles, qui envahissent des fissures au creusent des cavités dans la roche. On en connaît plusieurs centaines d'espèces mais leur diversité est sans doute encore sous-estimée, leur étude étant encore récente.
Les champignons altèrent les roches et les minéraux suivant différents mécanismes :
- les mécanismes physiques comprennent la pénétration des structures d'ancrage des champignons (notamment les rhizines des lichens foliacés) associée à des cycles d'expansion et de contraction du thalle (lors de cycles d'humidification et de dessèchement), et le forage direct. Dans le cas des champignons microcoloniaux la destruction mécanique implique la turgescence intracellulaire et la production extracellulaire de polysaccharides[69]. De petits grains rocheux ou minéraux peuvent être incorporés au thalle, de même que des minéraux néo-formés chimiquement. Cette détérioration, amplifiée par l’arrachage d'une partie des lichens par certains animaux, peut être visible et significative au bout de seulement quelques années ;
- l'altération biochimique est principalement due à l'excrétion de CO2, d'acides organiques, de sidérophores et d'autres métabolites, qui dissolvent les cations de la roche par complexation. Le principal de ces agents est sans doute l'acide oxalique, mais les lichens produisent aussi une grande variété d'acides spécifiques très corrosifs[70].
L'activité fongique a aussi pour effet le dépôt de nouveaux minéraux qui résultent de réactions d'oxydoréduction à la surface des minéraux, ainsi que de l’excrétion de minéraux par les champignons. Ces néo-minéraux (ou minéraux secondaires) sont notamment des carbonates, des phosphates, des oxydes et des oxalates. Les oxydes de manganèse, notamment, sont des composants communs du vernis noir recouvrant les roches altérées.
Rôle écologique

Les champignons jouent un rôle central dans beaucoup d’écosystèmes, notamment en tant que symbiotes des arbres, mais surtout en tant que décomposeurs bouclant le cycle du carbone et de nombreux éléments. Avec les bactéries, ils sont les décomposeurs qui participent le plus à la dégradation de la matière organique et à la production d'humus dans les écosystèmes terrestres et jouent un rôle primordial dans les cycles biogéochimiques et les chaînes alimentaires, accélérant le recyclage de nombreux éléments comme l'azote, le phosphore et le potassium[71]. Certains champignons sont actifs dans les milieux humides et aquatiques, d'autres champignons mycorhiziens ont un rôle clé dans les environnements arides en assurant principalement la pérennité du couvert végétal[72]. Le mycélium fongique peut atteindre dans le sol des forêts une biomasse de 12 tonnes par hectare, constituant alors un feutrage blanc très dense d'ascomycètes et de zygomycètes[73]. La décomposition de la matière organique végétale par les champignons est une étape essentielle du cycle du carbone. Les champignons sont une source majeure de nourriture pour de nombreux animaux, invertébrés (ex : certaines espèces de fourmis qui les cultivent) mais aussi quelques mammifères dont l'écureuil, le sanglier ou l'ours brun.
Quelques champignons, comme les Zoopagales (voir Zoopagomycotina), sont des prédateurs de Nématodes qu’ils capturent au moyen d’anneau ou de pièges adhésifs.
Les champignons peuvent provoquer des biodétériorations posant problème, comme lors de contamination et d’altérations organoleptiques de produits alimentaires ou lors de dégradation ou altération de l'aspect physique de divers produits tels que le bois, le papier, des textiles, les peintures, les métaux, la pierre ou même le verre. Divers mécanismes intervenant dans la sélection naturelle leur permettent de s'adapter à certains biocides antifongiques quand ces derniers sont utilisés systématiquement.
Les champignons jouent également un rôle important au niveau du mycobiome, autrement dit la composante fongique du microbiome, notamment le microbiote de l'organisme humain. Le mycobiome dans la bouche ou sur la peau des humains contient une centaine de genres de champignons tandis que celui de l'intestin contient des centaines d'espèces de champignons, la plupart commensaux mais certains pouvant devenir pathogènes, tel Candida albicans, principal responsable de la candidose[74].
Biomasse fongique
Contrairement à une idée répandue qui veut que le poids global de tous les organismes vivant dans l'eau, l'air et le sol, soit majoritairement représenté par des êtres « visibles » de la planète, ce sont les êtres « invisibles » qui constituent la majorité. La biomasse végétale — composée des champignons microscopiques dans le sol, des racines[75] et des plantes « visibles[76] » en surface — constitue 50 % de la biomasse terrestre[77] et 75 % de la biomasse terrestre est dans le sol (champignons microscopiques et bactéries constituant l'essentiel de la biodiversité du sol)[78]. La biomasse fongique, constituée essentiellement de microchampignons, représente près de 25 % de la biomasse terrestre, soit l'équivalent de deux mille milliards d'humains[79]. Cette composante microbienne du sol se traduit ainsi par une implication forte dans les fonctions et les services écosystémiques assurés par les matrices environnementales[80].
Équilibre forestier
L'attitude des forestiers à l'égard du champignon est parfois ambiguë, car il est tantôt l'indispensable auxiliaire de la forêt et de son sol (rôle pédologique majeur), tantôt un facteur de dégradation commerciale et technique du bois (coloration, biodégradation, maladies fongiques…) et tantôt un aliment ou une source de revenu parfois importante[81] (truffes notamment).
En outre de très nombreux champignons interagissent fortement avec les radionucléides présent dans le sol ou l'eau du sol, au point d'être probablement le plus important facteur biotique intervenant dans leur mobilité[82], même pour des champignons qui ne fructifient jamais hors du sol, tels les truffes Elaphomyces granulatus qui bioacumulent fortement certains radionucléides (Césium notamment), D'autres champignons terricoles comestibles font de même (dont laccaire améthyste (Laccaria amethystina) et le bolet bai (Xerocomus badius) ou de Cortinarius caperatus (anc. Rozites caperatus), la pholiote ridée, très consommée dans les pays de l'Est (et interdite d'importation en France depuis la catastrophe de Tchernobyl)[83].
Rôle économique
Les champignons jouent un rôle considérable dans nos sociétés, pas tellement parce qu'on les mange, car ils constituent rarement un aliment de base, mais parce que les maladies des plantes, dite maladies cryptogamiques, qu'ils occasionnent (surtout les ascomycètes) réduisent fortement l'offre alimentaire : ils sont responsables de 85 % des maladies de plantes et de 30 % des maladies émergentes actuelles[84]. Par ailleurs, les champignons en produisant des mycotoxines altèrent gravement les aliments et ils ont été à l'origine de graves intoxications qui ont marqué l'histoire de l'humanité. Ces toxines sont responsables selon la FAO de la perte en rendement de 25 % des récoltes mondiales[85].
En formant des mycorhizes, ils jouent un rôle essentiel dans la production végétale que ce soit dans le domaine forestier (surtout les basidiomycètes) ou les cultures annuelles (surtout les gloméromycètes). 90 % de toutes les espèces de plantes supérieures sont associées à ces mycorhizes[86].
Leurs métabolites secondaires sont sources de nombreux médicaments ou drogues.
Dans l’élaboration des aliments (pain, vin, bière, fromage…) et dans les industries biotechnologiques, ils jouent un rôle essentiel. Ils interfèrent aussi dans la dégradation ou le recyclage de matériaux. On trouve d'autres usages plus anecdotiques, comme le « briquet préhistorique » dit amadouvier.
Utilisation en alimentation
De nombreuses espèces comestibles et charnues sont utilisées à des fins alimentaires, notamment en soupes, sautées, en omelette, en friture (tenpura) ou en fricassée. La plupart des champignons n'ont pas d'intérêt culinaire ou sont toxiques[24], mais certaines espèces comestibles sont très recherchées pour leur saveur : le cèpe de Bordeaux, la truffe noire, l'oronge, etc. Le ramassage des champignons, activité encore vivace et populaire, constitue une subsistance des systèmes socio-économiques de cueillette. Celle-ci n'est pas sans risques car diverses espèces sont toxiques, voire mortellement vénéneuses, à l'origine de mycétisme, empoisonnement par méconnaissance des champignons. Des données récentes laissent penser qu'on a pu sous-estimer la toxicité naturelle ou acquise (bioconcentration de métaux lourds, accumulation de toxines avec l'âge) pour nombre de champignons[87] dont certains encore considérés comme comestibles[83]. C'est le cas par exemple de deux champignons très consommés en Chine et de plus en plus dans le monde ; l'oreille de Judas (Auricularia auricula-judae et espèces voisines) cause du syndrome de Szechwan, une atteinte plaquettaire, découverte par des dentistes intrigués par des saignements répétés chez des patients consommateurs réguliers de cuisine chinoise. C'est aussi le cas du shiitaké (Lentinula edodes) qui peut provoquer une « rare et sévère toxidermie » (éruptions cutanées liées à un mécanisme immunologique) autrefois uniquement connue au Japon et maintenant décrite en Europe (Royaume-Uni, France)[88].
Champignons cultivés

Si la culture des champignons est attestée dès l'Antiquité, peu d'espèces en Europe, malgré les différents progrès réalisés au cours du XXe siècle, se révèlent intéressantes pour une culture de type industriel ou semi-industriel. Le champignon de Paris (Agaricus bisporus) est le plus cultivé dans le monde. Il représente plus du tiers des champignons comestibles produits annuellement, suivi par le shiitaké et les pleurotes[89]. Par contre en Extrême-Orient, les espèces cultivées se multiplient au fil des années, avec des champignons tels que le shiitaké, l'éringî (nom japonais), la poule de bois, la collybie à pied de velours ou le champignon noir. La culture des champignons est appelée la myciculture (à ne pas confondre avec la mycoculture, une technique de culture utilisée en laboratoire pour les mycètes d'intérêt médical ou vétérinaire).
Principales espèces cultivées
- Agaricus bisporus — champignon de Paris
- Lentinula edodes — shiitaké (椎茸 cultivé à grande échelle en Extrême-orient)
- Pleurotus ostreatus — pleurote en huître et autres espèces voisines
- Pleurotus eryngii — pleurote du panicaut, éringi cultivé au Japon (エリンギー)
- Grifola frondosa — poule de bois le maitaké cultivé au Japon (舞茸 マイタケ)
- Hypsizygus tessulatus — buna-shimeji ブナシメジ rare en Europe, cultivé au Japon.
- Auricularia auricula-judae — oreille de Judas, 木耳 le « champignon noir » de la cuisine chinoise.
- Flammulina velutipes — Collybie à pied de velours énokidaké (エノキダケ les primordiums sont cultivés en bouteilles au Japon, leurs pieds très allongés sont alors très tendres).
- Volvariella volvacea — volvaire asiatique (袋竹)
- Cyclocybe aegerita — pholiote du peuplier (柳松茸 ヤナギマツタケ)
- Pholiota nameko — pholiote asiatique naméko (舐子 ナメコ)
Production
Il s’agit de champignons alimentaires sans distinction d’espèce.
|
Production en tonnes. Chiffres 2003-2004 | |||||
| Chine | 1 309 455 | 42 % | 1 359 335 | 42 % | |
| États-Unis d'Amérique | 391 000 | 12 % | 391 000 | 12 % | |
| Pays-Bas | 263 000 | 8 % | 260 000 | 8 % | |
| France | 165 647 | 5 % | 170 000 | 5 % | |
| Pologne | 120 000 | 4 % | 120 000 | 4 % | |
| Espagne | 115 165 | 4 % | 115 165 | 4 % | |
| Italie | 90 000 | 3 % | 90 000 | 3 % | |
| Canada | 78 018 | 2 % | 80 000 | 2 % | |
| Royaume-Uni | 77 100 | 2 % | 80 000 | 2 % | |
| Irlande | 69 000 | 2 % | 70 000 | 2 % | |
| Japon | 67 000 | 2 % | 67 000 | 2 % | |
| Autres pays | 403 726 | 13 % | 404 238 | 13 % | |
| Total | 3 149 111 | 100 % | 3 206 738 | 100 % | |
Mycotoxines
Les mycotoxines sont responsables d'intoxications plus ou moins graves. Trois genres de champignons peuvent causer le syndrome phalloïdien qui se traduit par des troubles digestifs et une hépatite aigüe pouvant devenir fulminante : Amanita (9 espèces), Lepiota (24 espèces) et Galerina (9 espèces)[90].
Les toxines du Cortinaire couleur de rocou et du Cortinaire très joli (en) s'attaquent au rein, certains intoxiqués devant subir une transplantation rénale ou des séances de dialyse à vie[91].
Chez l'humain et d'autres mammifères, les spores du Schizophylle commun ont la possibilité de germer dans ou sur différents organes et d'être à l'origine d'infections du système respiratoire (graves œdèmes, notamment chez les personnes immunodéficientes)[92], mais aussi des yeux, de la bouche, et d'onychomycoses[93].
Effet sur la santé
Influence économique
Utilisations diverses


Près de 700 espèces de champignons, tel le shiitake ou le polypore en touffes, sont utilisés à des fins médicinales. La consommation de ces champignons médicinaux ou de leurs extraits a donné naissance à une branche de la phytothérapie, la mycothérapie[95].
Les champignons contiennent souvent des molécules organiques très complexes, plus ou moins toxiques. La pénicilline et de nombreux médicaments sont tirés de champignons. L'amadouvier, puissant hémostatique, est utilisé en médecine chinoise traditionnelle[96]. D'autres peuvent avoir des vertus psychotropes (voir l'article détaillé Champignon hallucinogène), contenant des substances dites psychédéliques.
Les « champignons filamenteux » (basidiomycètes surtout) intéressent les acteurs des biotechnologies de par leurs éventuelles capacités à rapidement biotransformer les lignocelluloses grâce à des enzymes spécialisés, ou à dépolluer certains matériaux[97] (INRA Avignon, en France Divers programmes de recherche visent à comprendre et maîtriser certains mécanismes de biotransformation fongique pour les utiliser industriellement, dont pour produire des carburants biosynthétisés. Là encore, certains craignent un risque en cas de fuite dans l'environnement d'organismes génétiquement modifiés (OGM) susceptibles de s'attaquer à des ligneux ou autres végétaux (vivants et/ou morts).
Technologies vertes Dépollution Paul Stamets et d'autres mycologues préconisent de développer la permaculture de champignons, et considèrent la fungiculture comme une source importante de nourriture et de molécules utiles pour le futur. Elle semble aussi intéressante pour la bioremédiation et la dépollution de certains sols ou matériaux ; en accompagnement de la phytoremédiation ou de l'utilisation de divers micro-organismes ; utilisées seuls ou en association èpuratrice, etc. Certaines espèces captent et stockent remarquablement bien les métaux. La mycoremédiation (parfois traduit par fongoremédiation), via la mycofiltration notamment, permettrait ainsi de détoxiquer des milieux (eau, air, sol) de façon moins coûteuse qu'avec les techniques physico-chimiques classiques et plus rapide que via la phytoremédiation. Elle demande encore cependant une meilleure connaissance et maîtrise de la culture des mycéliums dans un sol ou un substrat pollué ou dans un matériau filtrant un air ou une eau polluée.
Beaucoup d'espèces bioconcentrent fortement les métaux lourds et certaines les radionucléides (Elaphomyces granulatus par exemple), contribuant à remettre en circulation des métaux qui ont été provisoirement piégés dans des organismes animaux ou végétaux, ou naturellement présents dans le sol sur certains sites métallifères.
Champignons extrêmes
Les mycètes peuvent atteindre des tailles insoupçonnées. Les fossiles de prototaxites sont de nos jours classés comme des anciens champignons de deux à neuf mètres de hauteur pour un mètre de circonférence.
Ils auraient été les plus grands organismes terrestres du Silurien et du Dévonien, entre -420 et -350 Ma.
De nos jours, c'est également un champignon qui détient le record de plus grand être vivant au monde (bien que la notion d'organisme soit discutable dans ce cas) : un mycélium de l'espèce Armillaria ostoyae couvrant près de 9 km2 (880 hectares) et pesant près de 2 000 tonnes a été identifié en 2000 dans l'Oregon, par des tests d'ADN[98],[99]. Son âge est estimé entre 2 000 et 10 000 ans[100] selon l'évaluation de sa vitesse de croissance. Le record précédent datant de 1992 était « seulement » de 600 hectares.
État des populations, menaces
Comme pour de nombreuses autres espèces, beaucoup d'espèces de champignons sont en régression. Il existe dans un nombre croissant de pays et régions des listes rouges d'espèces fongiques menacées.

À titre d'exemple, la liste rouge des champignons menacés de Suisse (limitée aux champignons supérieurs), mise à jour par l’Office fédéral suisse de l’Environnement en 2007, alerte sur le fait que sur 2 956 espèces et sous-espèces à propos desquelles des données fiables et suffisantes existent, 937 espèces (32 %) ont été classées comme menacées par l’Institut fédéral de recherche sur la Forêt, la Neige et le Paysage (WSL). Une espèce est éteinte, 3 % sont « en danger critique d'extinction », 12 % sont « en danger » et 17 % sont vulnérables. 63 % sont considérées comme non menacées, mais l'état des populations de 2004 des autres espèces (40 % du total des champignons supérieurs connus en Suisse) n'a pu être évalué, faute de données. En toute logique, les espèces les plus menacées sont celles dont les milieux ont le plus rapidement ou le plus fortement régressé (champignons des prés et pâturages maigres, des marais et liés au bois mort). Les espèces sont également jugées plus menacées en altitude où elles sont moins nombreuses. Sur les 937 espèces menacées, 15 % sont des champignons forestiers. C'est probablement moins que dans d'autres pays voisins grâce à la Liste rouge. Cela est certainement dû à une sylviculture plus « proche de la nature » (de type prosilva) qui a su conserver une relative naturalité aux forêts et du gros bois mort.
Deux études françaises ont montré que les fongicides, étaient dans le Nord de la France présents plusieurs jours par an en quantité très significative dans la pluie et l'air, au point qu'on ne peut plus parler de traces.
De nombreuses espèces de lichens ont également fortement régressé, même si celles qui étaient indicatrices de pollution acide réapparaissent.
Valeur de bioindication
Les communautés fongiques se montrent très sensibles à certains facteurs environnementaux, dont au pH[101] et à d'autres facteurs édaphiques, ce qui leur confère une intéressante valeur bioindicatrice[102] ou donne des informations sur ses plantes-hôte en forêt[103].
Dans ou sur les sols agricoles et forestiers notamment, ou sur les bois-morts, le degré de pauvreté ou richesse (en terme d'espèces, mais aussi de diversité génétique) en champignons (dont mycorhiziens ou associés aux algues dans le cas des lichens), est bioindicatrice de la qualité du milieu, et dans une certaine mesure de sa naturalité ou son ancienneté[104].
Très résistants à la plupart des métaux lourds, ils sont de mauvais bioindicateurs de leur présence, mais ce sont souvent d'excellent biointégrateurs qui peuvent informer sur les métaux bioaccumulables dans les sols pollués et le degré de bioconcentration de certains polluants, par exemple les métaux lourds ou les radionucléides[105].
Comestible ou toxique ?
On a identifié à ce jour une vingtaine de champignons mortels dans le monde, une trentaine d'excellents comestibles et une grande masse de champignons immangeables car trop amers, âcres, nauséabonds, coriaces, fibreux ou trop minuscules. Comme il n'existe aucune méthode fiable pour les identifier, il importe d'abord de connaître les champignons dangereux et ensuite sélectionner les seuls champignons comestibles sûrs et savoureux, idéalement lors de sorties sur le terrain avec un connaisseur. La liste de champignons toxiques et comestibles peut être consultée chez un pharmacien (en France), ou sur le site de la Société mycologique de chaque région.
Deux types de toxicité sont à considérer :
- La toxicité intrinsèque de certaines espèces liée à des toxines organiques produites par le champignon, qui provoquent par exemple des hallucinations, des douleurs abdominales, nausées, diarrhée sanglante, coliques, paralysies pouvant conduire à la mort (voir les détails sur la page mycotoxicologie) ;
- La toxicité induite par la forte capacité de certains champignons (dont des espèces comestibles et recherchées) à bioaccumuler certains métaux lourds toxiques (dont mercure, plomb, cadmium, sélénium, et, à un moindre degré cobalt, nickel et chrome (le chrome VI est très toxique)[106]). Selon une méta-analyse récente (2021) consacrée à la quantité de métaux lourds présente dans les champignons comestibles, « bien que les champignons soient considérés comme des aliments sains et précieux, ils peuvent être une source de métaux lourds éventuellement toxiques susceptibles de nuire à la santé »[107] ; selon 59 études publiées de 1970 à mi-2020, les métaux les plus bioaccumulés sont, par ordre d'importance décroisante : Fe (204,5716 mg/kg) > Zn (154,6076 mg/kg) > Cu (32,1996 mg /kg) > Mn (25,4007 mg/kg) > Ni (10,4375 mg/kg) > Cr (4,934 mg/kg) > Pb (2,4844 mg/kg) > Cd (1,3925 mg/kg) ; les trois derniers (Cr > Pb > Cd) étant les plus toxiques[107] ; et il a été constaté ue Pb, Cu, Fe, Cd, Cr et Zn s'accumulent plutôt dans les chapeau alor que Mn et Ni sont plutôt concentrés dans les stipes. Au vu des taux de métaux lourds bioaccumulés par les champignons, dans certains pays leur consommation peut exposer à « des risques non cancérigènes importants »[107].
Chez 15 espèces différentes de champignons sauvages comestibles échantillonnés en milieu urbain et en milieu forestier sous « influence anthropique limitée », pour 46 éléments chimiques recherchés, les taux de nutriments et de polluants variaient beaucoup selon les cas, mais avec en général pas ou peu de différence entre la ville et la forêt. Selon cette étude (2021), les taux d'éléments majeurs varient plutôt selon les affinités propres à l'espèce (facteur génétique), mais pour les éléments traces métalliques, c'est le contexte environnemental (fond géochimique et/ou pollution locale) qui semble décisif[108] ; pour le plomb par exemple, la pollution routière ne semble pas ou plus avoir d'influence : des taux similaires de Pb sont retrouvées dans le champignon, qu'il provienne de la forêt ou d'une zone urbaine. Et la quantité de métaux bioaccumulée varie peu selon que le substrat soit le bois ou le sol hormis pour l'As et le Mo moins concentrés dans les champignons lignicoles[108].
L'évaluation des risques pour la santé indique un « risque élevé non cancérigène » pour les métaux dans 14 des 19 espèces de champignons étudiés les zones urbaines et forestières. Selon les auteurs, tant en ville qu'en forêt, « la consommation de quantités excessives des champignons étudiés peut avoir des effets néfastes sur la santé »[108].
La teneur en éléments toxiques était faible et aucune influence de la pollution routière n'a pu être détectée, car des concentrations similaires de Pb ont été trouvées dans des champignons provenant de sites forestiers et urbains. Alors que les concentrations de métaux ne présentaient pas de variations selon le substrat (bois et sol), il a été observé que les champignons poussant sur bois présentaient généralement des teneurs plus faibles en As et Mo.
Néanmoins, l'évaluation des risques pour la santé a révélé un risque élevé non cancérigène des métaux dans 14 des 19 champignons étudiés dans les zones urbaines et forestières, ce qui suggère que ces champignons doivent être consommés avec prudence.
Les taux de cadmium mesurés dans les champignons de certaines régions où le sol est naturellement riche en cadmium ou pollué par du cadmium anthropique sont suffisants pour poser de graves problèmes de néphrotoxicité (attaque du système rénal), voire exceptionnellement pour tuer par empoisonnement aigu[106]. L'exposition à des doses souvent faibles à moyennes de radionucléides via l'exposition de champignons a des effets qui sont encore très discutés pour les faibles doses, mais les études qui ont suivi la catastrophe de Tchernobyl ont montré que le champignon était l'une des premières sources de radioactivité dans l'alimentation dans les zones de retombées du nuage[109].
En cas d'empoisonnement, le médecin peut confondre ces deux types d'intoxications.
La contamination et l'empoisonnement occasionnels d'animaux tels que vaches, chevaux, chèvres moutons par les métaux lourds pourraient en partie être dus à la consommation de champignons, y compris d'espèces à fructification souterraines, qui passent inaperçues, telles que la truffe du cerf ou les truffes recherchées par les sangliers, les écureuils ou quelques micromammifères.
Comme le rappelle Didier Michelot du CNRS, la possibilité d'empoisonnements graves, distincts de ceux produits par les toxines organiques, et dus à la consommation de spécimens appartenant aux genres (Agaricus, Pleurotus, etc.) n'est pas exclue en raison de leur capacité à concentrer des métaux toxiques (dont cadmium, plomb, mercure…) à des doses très supérieures aux seuils toxicologiques.
À titre d'exemple et à partir des analyses faites par D. Michelot (CNRS) en France, on peut retenir qu'un repas typique composé de 200 g (portion moyenne) d'Agaricus arvensis frais, espèce très appréciée des cuisiniers, contenait en France 2 mg de cadmium, soit 100 fois la dose permise par les autorités de santé publique[110].

Des risques similaires sont posés par d'autres champignons, dont certains recherchés par les amateurs :
- Agaricus silvicola (30,6 ppm de cadmium),
- Agaricus bresadolianus (10,7 ppm de cadmium) et, moindrement ;
- Suillus variegatus (4 ppm de cadmium).
Les Agaricales accumulent les plus grandes quantités.
La teneur la plus élevée en mercure est détectée chez
- Suillus variegatus (94 ppm)
- Agaricus aestivalis (87,4 ppm),
- Agaricus arvensis (84,1 ppm),
- Pleurotus eryngii (82 ppm).
Le plomb a été détecté à des taux très élevés chez
- Agaricus bresadolanus (52,2 ppm),
- Morchella esculenta (44,2 ppm),
- Fistulina hepatica (42,7 ppm),
- Clitocybe nebularis (43 ppm),
- Leccinum crocipodium (Boletus) (42,1 ppm).
Il est par ailleurs probable que les champignons symbiotes jouent un rôle dans l'accumulation de métaux dans le bois.
Aspects culturels
Notes et références
Notes
- ↑ On trouve la trace du vieux verbe eschamper, « fuir le champ de bataille, se sauver » dans l'expression figée « prendre la poudre d'escampette » et dont témoigne aussi l'italien scampo, « fuite ».
- ↑ Mot d'origine très ancienne dont la filiation remonte, selon Robert Gordon Wasson, aux appellations paléosibériennes Ponk, Pongas, Hango[13].
- ↑ Sont concernées les roches en place dans la nature, mais aussi les pierres des monuments et ces roches artificielles que sont le ciment et le béton.
Références
- ↑ Au pluriel pour le taxon Fungi. Le terme existait déjà en français comme synonyme de champignon (XVIe siècle, = fungusses en vieil anglais), et aussi au singulier féminin désigne l'ensemble des champignons d'un même territoire, comme pour la faune et la flore pour les animaux et les plantes.
- ↑ C'est un des noms latin du « règne » en biologie (mycologie), au sens large de champignons ; exemples Hypocrea fungicola P. Karst., renvoie à son écologie inféodée au Fomes pinicola, ce polypore étant lui même dépendant du Pin. Cf Paul Escallon 1988, Précis de Myconymie ; SMBC & FMDS [ouvrage dédié à Henri Romagnesi... le Mycologue, le Philologue... et le Poète>]. Thonon les Bains, France
- ↑ Probablement de l'indo-européen *meu qui l’apparente au latin muscus (« mousse »), mucus (« morve »), mucor (« moisissure ») et au grec mykès (d'où les Mycètes) désignant d'abord les champignons en général. Les termes grec et latin sont ainsi une allusion possible aux champignons qui se protègent contre la dessication par une couche de mucus qui recouvre leur chapeau et parfois aussi leur pied, ou à la mycophobie ancestrale, les champignons étant associés aux mucosités nasales repoussantes.
René Pomerleau, Flore des champignons du Québec, Éditions la presse, 1980, p. 87. [> fonge du Québec?].
On retrouve les mêmes étymologies en japonais, sauf que le gluant (Naba, Namé) est associé à un délice gastronomique (mycophilie). Pholiota nameko. [Imazeki, Rokuya. 1973. Japanese mushroom names. The Transactions of the Asiatic Society of Japan, 3rd series, 11:26–80.]
(étymon qui a également donné mucus, mucilage, muqueux, muqueuse, avec deux caractères opposés :
- matière gluante (collagène, « plus facile à saisir qu'à lâcher » cf. la célèbre tirade de Raimu dans Marius de Marcel Pagnol) ou
- visqueuse et gélatineuse (difficile à saisir) phénomène à double action comme la savonnette, par exemple, qui échappe des mains, en fonction de la proportion d'eau.
- ↑ macromycète, subst. masc.,bot. Champignon de taille importante. E. Fries (1794-1878) (Hist. gén. sc.,t.3, vol.1, 1961, p.434).
- 1 2 (en) Joseph Heitman, Barbara J. Howlett, Pedro W. Crous, Eva H. Stukenbrock, Timothy Yong James, Neil A. R. Gow, The Fungal Kingdom, John Wiley & Sons, , p. 31
- ↑ (en) Meredith Blackwell, « The Fungi. 1, 2, 3 … 5,1 Million species ? », American Journal of Botany, vol. 98, no 3, , p. 426–438 (DOI 10.3732/ajb.1000298).
- ↑ (en) Leho Tedersoo, Mohammad Bahram, Sergei Põlme, Urmas Kõljalg, Nourou S. Yorou, Ravi Wijesundera, Luis Villarre, « Global diversity and geography of soil fungi », Science, vol. 346, no 6213, (DOI 10.1126/science.1256688).
- ↑ Louis-Jean Calvet, Histoires de mots : étymologies européennes, Éditions Payot, , p. 137.
- ↑ (en) Philip G. Miles, Shu-Ting Chang, Mushrooms. Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, CRC Press, , p. 1
- ↑ Informations lexicographiques et étymologiques de « Champignon » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales
- 1 2 René Pomerleau, Flore des champignons au Québec et régions limitrophes, Éditions la presse, , p. 87.
- ↑ Philippe Bouchet, Jean-Louis Guignard, Yves-François Pouchus, Les champignons. Mycologie fondamentale et appliquée, Elsevier Masson, , p. 2
- ↑ René Pomerleau, Flore des champignons au Québec et régions limitrophes, Éditions la presse, , p. 87.
- ↑ Louis-Jean Calvet, Histoires de mots : étymologies européennes, Éditions Payot, , p. 138
- ↑ (en) Stefanie Pöggeler et Johannes Wöstemeyer, Evolution of Fungi and Fungal-Like Organisms, Springer, , p. 5
- ↑ Les ballistospores (en) sont par exemple des basidiospores détachées de la baside qui les porte par des courants d'air qui les lancent dans l'environnement.
- ↑ (en) Galante TE, Horton TR, Swaney DP. –. Mycologia 103(6),., « 95% of basidiospores fall within 1 m of the cap: a field–and modeling–based study », Mycologia, vol. 103, no 6, , p. 1175–1183
- ↑ Les enzymes digestives des limaces stimulent la germination des spores. Cf (en) HW Keller, KL Snell, « Feeding activities of slugs on Myxomycetes and macrofungi », Mycologia, vol. 94, no 5, , p. 757–760
- ↑ Les spores qui utilisent ce type de vecteur ont des parois plus épaisses, souvent renforcée par des pigments mélaniques qui évitent aux spores d'être digérées. Cf (en) Rast D, Stussi H, Hegnauer H, Nyhlen L., « Melanins », in The Fungal, Spore: Morphogenetic Controls, Turian G, Hohl HR (Eds), Academic Press, 1981, p. 507–531.
- ↑ Rongeurs, écureuils, cerfs.
- ↑ Ces pigments fongiques peuvent à l'inverse avoir un rôle de défense contre les herbivores.
- ↑ (en) Erik Lilleskov, Tom D. Bruns, « Spore dispersal of a resupinate ectomycorrhizal fungus, Tomentella sublilacina, via soil food webs », Mycologia, vol. 97, no 4, , p. 762-769
- ↑ Francis Martin, Tous les champignons portent-ils un chapeau ?, Quae, , p. 18.
- 1 2 « Ni animal ni végétal, tout simplement à part », sur webzine-biodiversite.developpement-durable.gouv.fr (consulté le )
- ↑ cnrtl
- ↑ (en) A. Tsuneda, Fungal Morphology and Ecology, Tottori Mycological Institute,
- ↑ Jean-Claude Callen et Roland Perasso, Biologie cellulaire. Des molécules aux organismes, Dunod, , p. 184
- ↑ (en) José Ruiz-Herrera, Fungal Cell Wall: Structure, Synthesis, and Assembly, CRC Press, , 256 p. (lire en ligne)
- ↑ (en) Kevin Kavanagh, Fungi: Biology and Applications, John Wiley & Sons, (lire en ligne), p. 6
- ↑ (en) J K Misra et S. K. Deshmukh, Fungi from different environments, Science Publishers, , p. 260, 361.
- ↑ Guy Claus, « Des odeurs en mycologie ! », Documents mycologiques, vol. 8, nos 30-31, , p. 31-63
- ↑ Marc-André Selosse, François Le Tacon, « Les stratégies symbiotiques de conquête du milieu terrestre par les végétaux », Année Biol., vol. 40, , p. 3-20 (lire en ligne)
- ↑ (en) Andrew W. Wilson, Manfred Binder et David S. Hibbett, « Effects of gasteroid fruiting body morphology on diversification rates in three independent clades of fungi estimated using binary state speciation and extinction analysis », Evolution, vol. 65, no 5, , p. 1305–1322 (DOI 10.1111/j.1558-5646.2010.01214.x, lire en ligne, consulté le ).
- ↑ Jean-Christophe Guéguen et David Garon, Biodiversité et évolution du monde fongique, EDP Sciences, , p. 92.
- ↑ (en) Stefanie Pöggeler et Johannes Wöstemeyer, Evolution of Fungi and Fungal-Like Organisms, Springer, , p. 171
- ↑ (en) T. N. Taylor, H. Hass & H. Kerp, « The oldest fossil ascomycetes », Nature, vol. 399, no 648, (DOI 10.1038/21349)
- ↑ (en) Lucking R, S Huhndorf, Pfister D, Plata ER, Lumbsch H., « Fungi evolved right on track », Mycologia, vol. 101, no 6, , p. 810-822 (DOI 10.3852/09-016)
- 1 2 Lise Loumé, « Les champignons sont apparus sur Terre 300 millions d'années plus tôt que prévu », sur sciencesetavenir.fr,
- ↑ (en) Corentin C. Loron, Camille François, Robert H. Rainbird, Elizabeth C. Turner, Stephan Borensztajn et Emmanuelle J. Javaux, « Early fungi from the Proterozoic era in Arctic Canada », Nature, (DOI 10.1038/s41586-019-1217-0).
- ↑ (en) Miguel A. Naranjo‐Ortiz Toni Gabaldón, « Fungal evolution: major ecological adaptations and evolutionary transitions », Biological Letters, vol. 94, no 4, , p. 1443-1476 (DOI 10.1111/brv.12510)
- ↑ (en) S. Bonneville, F. Delpomdor, A. Préat, C. Chevalier, T. Araki, M. Kazemian, A. Steele, A. Schreiber, R. Wirth and L. G. Benning, « Molecular identification of fungi microfossils in a Neoproterozoic shale rock », Science Advances, vol. 6, no 4, (DOI 10.1126/sciadv.aax7599)
- ↑ (en) Paul Stamets, Mycelium running. How mushrooms can help save the world, Potter/TenSpeed/Harmony, , p. 47.
- ↑ (en) Mary E. White, Earth Alive ! From Microbes to a Living Planet, Rosenberg Pub, , p. 93
- ↑ Marc-André Selosse, François Le Tacon, « Les stratégies symbiotiques de conquête du milieu terrestre par les végétaux », Année Biol., vol. 40, , p. 16-17 (lire en ligne)
- ↑ Marc-André Selosse, op. cit., p. 16
- ↑ (en) Mark C. Brundrett, « Coevolution of roots and mycorrhizas of land plants. 154, », New Phytologist, vol. 154, no 2, , p. 275-304 (DOI 10.1046/j.1469-8137.2002.00397.x)
- ↑ Francis Martin, Tous les champignons portent-ils un chapeau ?, Éditions Quae, , p. 41
- ↑ Jean Després, L'univers des champignons, Les Presses de l'Université de Montréal, (lire en ligne), p. 123.
- ↑ (en) R.H. Whittaker, « On the broad classification of organisms », The Quarterly Review of Biology, vol. 34, no 3, , p. 210-226 (DOI 10.1086/402733).
- ↑ D.S. Hibbett, et al., « A higher-level phylogenetic classification of the Fungi », Mycological Research, vol. 111, no 5, , p. 509-547 (lire en ligne [PDF])
- ↑ (en) Geoffrey Clough Ainsworth, The Fungi : An Advanced Treatise, Volumes 1 à 5, Academic Press, , 805 p.
- ↑ (en) David J. McLaughlin, Paul A. Lemke, Esther G. McLaughlin, Meredith Blackwell, Joseph W. Spatafora, Systematics and Evolution : the Mycota, Springer, , p. 3-4
- ↑ Catalogue of Life Checklist, consulté le 6 novembre 2020
- ↑ Silar P, Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes, HAL, (ISBN 978-2-9555841-0-1, lire en ligne [archive du ]), p. 462
- ↑ Esser K, The Mycota VII A: Systematics and Evolution (2nd ed.), Springer, coll. « The Mycota », (ISBN 978-3-642-55317-2, DOI 10.1007/978-3-642-55318-9, S2CID 46141350, lire en ligne), p. 461
- ↑ Leho Tedersoo, Santiago Sanchez-Ramırez, Urmas Koljalg, Mohammad Bahram, Markus Doring, Dmitry Schigel, Tom May, Martin Ryberg et Kessy Abarenkov, « High-level classification of the Fungi and a tool for evolutionary ecological analyses », Fungal Diversity, vol. 90, no 1, , p. 135–159 (DOI 10.1007/s13225-018-0401-0
 )
) - ↑ Les mycorhizogène se caractérisent généralement par une base de pied « propre » : les hyphes secrètent la glomaline, colle biologique qui enchevêtre et emmêle les particules minérales et organiques de sol, facilitant la formation de micro-agrégats (<250 µm) agglomérés en macro-agrégats (>250 µm) qui se cassent doucement à la main. Chez les saprotrophes, des débris organiques sont plus agglomérés à cette base (pied « sale ») dont les hyphes se prolongent dans la litière où elles agissent en tant que pourritures blanches de l'humus (distinctes de la pourriture blanche du bois). Cf (en) JM Tisdall, « Fungal hyphae and structural stability of soil », Australian Journal of Soil Research, vol. 29, no 6, , p. 729-743 (DOI 10.1071/SR9910729), Pierre Davet, Vie microbienne du sol et production végétale, éditions Quae, , p. 27
- ↑ (en) Judith Becker, Christoph Wittmann, « A field of dreams: Lignin valorization into chemicals, materials, fuels, and health-care products », Biotechnology Advances, vol. 37, no 6, (DOI 10.1016/j.biotechadv.2019.02.016)
- ↑ Hugues Demeude, Les incroyables pouvoirs de la nature, Flammarion, , p. 32
- ↑ Les mildious, les rouilles sont des maladies induites par des parasites obligatoires de la famille des Oomycètes.
- ↑ Jean-Marie Pelt, Franck Steffan, Les Langages secrets de la nature. La communication chez les animaux et les plantes, Fayard, , p. 227
- ↑ Régis Courtecuisse et Bernard Duhem, Guide des champignons de France et d'Europe : 1752 espèces décrites et illustrées, Paris, Delachaux et Niestlé, , 157 p. (ISBN 978-2-603-01691-6)
- ↑ (en) James R Hanson, The Chemistry of Fungi, Royal Society of Chemistry, , p. 127-138
- ↑ (en) Laurent Dufossé, Yanis Caro, Mireille Fouillaud, « Fungal Pigments: Deep into the Rainbow of Colorful Fungi », Journal of Fungi, vol. 3, no 3, , p. 45 (DOI 10.3390/jof3030045).
- ↑ (en) Jan Velisek, Karel Cejpek, « Pigments of higher fungi – a review », Journal of Food Sciences, vol. 29, no 2, , p. 87-102 (DOI 10.17221/524/2010-CJFS).
- ↑ (en) Franz-Sebastian Krah, Ulf Büntgen, Hanno Schaefer et al., « European mushroom assemblages are darker in cold climates », Nature Communications, vol. 10, no 2890, (DOI 10.1038/s41467-019-10767-z).
- ↑ Ce mélanisme constituerait une défense renforcée contre les pathogènes dont le développement est favorisé par le temps humide. Cf Franz-Sebastian Krah, op. cit.
- ↑ (en) Geoffrey M. Gadd, « Fungi, Rocks, and Minerals », Elements, vol. 13, , p. 171-176 (DOI 10.2113/gselements.13.3.171).
- ↑ (en) M. Marvasi et al., « Black microcolonial fungi as deteriogens of two famous marble statues in Florence, Italy », International Biodeterioration and Biodegradation, vol. 68, , p. 36-44.
- ↑ (en) P. Adamo et P. Violante, « Weathering of rocks and neogenesis of minerals associated with lichen activity », Applied Clay Science, vol. 16, , p. 229-256.
- ↑ (en) G.M. Gadd, Fungi in Biogeochemical Cycles, Cambridge University Press, , 469 p. (lire en ligne)
- ↑ (en) Brundrett MC. 1991. Mycorrhizas in natural ecosystems. In: Macfayden A, Begon M, Fitter AH, eds. Advances in ecological research, vol. 21. London, UK: Academic Press, 171 – 313.
- ↑ (en) L. Boddy, J.C. Frankland & P. Van West, « Ecology of Saprotrophic Basidiomycetes », British Mycological Society, vol. 28, , p. 277-300
- ↑ (en) Lijia Cui, Alison Morris et Elodie Ghedin, « The human mycobiome in health and disease », Genome Medicine, vol. 5, no 7, , p. 63
- ↑ « De 10 à 30 % dans une forêt, la biomasse racinaire peut atteindre 75 à 95 % de la biomasse totale dans des prairies, des toundras ou des pelouses arides d'altitude ». Cf Claire Tirard, Luc Abbadie, David Laloi, Philippe Koubbi, Écologie, Dunod, (lire en ligne), p. 423.
- ↑ Sachant que la biomasse végétale « visible » est cent fois plus importante que la biomasse animale. Cf (en) William B. Whitman, David C. Coleman & William J. Wiebe, « Prokaryotes: The unseen majority », PNAS, vol. 95, no 12, , p. 6578-6583 (DOI 10.1073/pnas.95.12.6578, lire en ligne).
- ↑ (en) Walter Larcher, Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, Springer Science & Business Media, , p. 10.
- ↑ (en) Garry Willgoose, Principles of soilscape and landscape evolution, Cambridge University Press, (lire en ligne), p. 163.
- ↑ (en) Author links open overlay panelJ.DavidMiller, « Fungi as contaminants in indoor air », Atmospheric Environment, vol. 26, no 12, , p. 2163–2172 (DOI 10.1016/0960-1686(92)90404-9)
- ↑ Lionel Ranjard, Philippe Cuny, Pierre-Alain Maron, Elisabeth d'Oiron Verame, La microbiologie moléculaire au service du diagnostic environnemental, ADEME, (lire en ligne), p. 13
- ↑ Olaf Schmidt, Wood and Tree Fungi Biology, Damage, Protection, and Use 10.1007/3-540-32139-X_8
- ↑ John Dighton, Tatyana Tugay et Nelli Zhdanova, « Interactions of Fungi and Radionuclides in Soil », dans Microbiology of Extreme Soils (résumé, lire en ligne), p. 333-355
- 1 2 Les champignons dits comestibles le sont-ils encore ? (D'après une conférence de Paul Pirot (de la Société Mycologique de Belgique) faite à Nismes (B) et Bellême (F) à l'automne 2010
- ↑ (en) Anderson PK, Cunningham AA, Patel NG, Morales FJ, Epstein PR, Daszak P., « Emerging infectious diseases of plants : Pathogen pollution, climate change and agrotechnology drivers », Trends in Ecology & Evolution, vol. 19, no 10, , p. 535–544
- ↑ (en) Mahendra Rai et Ajit Varma, Mycotoxins in Food, Feed and Bioweapons, Springer, , p. 201
- ↑ (en) Diana Marco, Metagenomics : Current Innovations and Future Trends, Horizon Scientific Press, , p. 162
- ↑ Dr L. Giacomoni, Les champignons. Intoxications, pollutions, responsabilités. Une nouvelle approche de la mycologie, Les éditions billes, Malakoff, 1989, 197 pp
- ↑ Dr Lucien Giacomoni, Un syndorme méconnu : la toxicodermie du Shiitaké, lentinula edodes (Berkeley) ; in bulletin de la S.M.F (Société Mycologique de France), tome 125, fascicules 3 et 4, 2009, p. 197-212, avec illustrations), http://cat.inist.fr/?aModele=afficheN&cpsidt=23113796 Résumé/Fiche INIST-CNRS]
- ↑ (en) R.B. Beelman, D.J. Royse et N. Chikthimmah, « Bioactive components in Agaricus bisporus of nutritional, medicinal or biological importance », in Science and Cultivation of Edible and Medicinal Fungi, Romaine C.P., Keil C.B., Rinker D.L. et Royse D.J. (Eds.), 2004, Pennsylvania State Univ., p. 1
- ↑ C. Bruneau, « Syndrome phalloïdien : quoi de neuf en 2018 ? », Toxicologie Analytique et Clinique, vol. 30, no 3, , p. 158–159 (DOI 10.1016/j.toxac.2018.07.080)
- ↑ Guillaume Eyssartier, Les 50 règles d'or du cueilleur de champignons, Larousse, , p. 39
- ↑ (en) Chowdhary A, Kathuria S, Agarwal K, Meis JF., « Recognizing filamentous basidiomycetes as agents of human disease: A review. », Medical Mycology, vol. 52, no 8, , p. 782-797 (PMID 25202126, DOI 10.1093/mmy/myu047).
- ↑ (en) J. P. Z. Siqueira, D. Sutton, J. Gené, D. García, M. Guevara-Suarez, C. Decock, N. Wiederhold et J. Guarro, « Schizophyllum radiatum, an Emerging Fungus from Human Respiratory Tract », Journal of clinical microbiology, vol. 54, no 10, , p. 2491–2497 (DOI 10.1128/JCM.01170-16, lire en ligne, consulté le ).
- ↑ Mirko Svrček, J. Kubička, Le multiguide nature des champignons d'Europe, Elsevier Sequoia, , p. 80.
- ↑ Alain Tardif, La mycothérapie : Médecine des champignons, Amyris, , 190 p. (ISBN 978-2-930353-45-6)
- ↑ B. Roussel, S. Rapior, C. Charlot, C.-L. Masson et P. Boutié, « Histoire des utilisations thérapeutiques de l'amadouvier », Revue d'histoire de la pharmacie, vol. 90, (lire en ligne)
- ↑ Exemple
- ↑ Canadian Journal of Forest Research, avril 2003
- ↑ (en) Rebecca Stefoff, The Fungus Kingdom, Marshall Cavendish, , p. 7-8
- ↑ (en) Judy Wearing, Fungi. Mushrooms, Toadstools, Molds, Yeasts, and Other Fungi, Crabtree Publishing Company, , p. 43
- ↑ Baath E, Anderson TH. 2003. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 35: 955-963.
- ↑ Lilleskov EA, Fahey TJ, Horton TR, Lovett GM. 2002. Below ground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology 83: 104-115
- ↑ Buée M, Maurice JP, Zeller B, Andrianarisoa S, Ranger J, Courtecuisse R, Marçais B, Le Tacon F. 2011. Influence of tree species on richness and diversity of epigeous fungal communities in a French temperate forest stand. Fungal Ecology 4: 22-31
- ↑ Diédhiou AG, Dupouey JL, Buée M, Dambrine E, Laüt L, Garbaye J. 2009. Response of ectomycorrhizal communities to past Roman occupation in an oak forest. Soil Biol. Biochem. 41: 2206-2213
- ↑ Universalia, Encyclopaedia Universalis France, , p. 166
- 1 2 http://toxicologie.pagesperso-orange.fr/metaux_6.htm
- 1 2 3 (en) Mohsen Dowlati, Hamid Reza Sobhi, Ali Esrafili et Mahdi FarzadKia, « Heavy metals content in edible mushrooms: A systematic review, meta-analysis and health risk assessment », Trends in Food Science & Technology, vol. 109, , p. 527–535 (DOI 10.1016/j.tifs.2021.01.064, lire en ligne, consulté le )
- 1 2 3 (en) Maja Ivanić, Martina Furdek Turk, Zdenko Tkalčec et Željka Fiket, « Distribution and Origin of Major, Trace and Rare Earth Elements in Wild Edible Mushrooms: Urban vs. Forest Areas », Journal of Fungi, vol. 7, no 12, , p. 1068 (ISSN 2309-608X, PMID 34947050, PMCID PMC8706631, DOI 10.3390/jof7121068, lire en ligne, consulté le )
- ↑ http://mycologia34.canalblog.com/archives/10___tchernobyl_et_les_champignons/index.html
- ↑ « Champignons - Définition et recettes de "Champignons" - Supertoinette », sur supertoinette.com (consulté le ).
Voir aussi
Bibliographie
- Dr_Lucien_GiacomoniNicotPautot1982">[Becker et al. 1982] Georges Becker, Dr Lucien Giacomoni, Jacqueline Nicot, Serge Pautot, Guy Redeuihl, Geneviève Branchu, Domiique Hartog, Anke Hérubel, Helga Marxmuller, Ursula Millot et Claude Schaeffner, Le guide des champignons, Reader's Digest, (ISBN 2-7098-0031-4).
- [Bon 2004] Marcel Bon, Champignons de France et d'Europe occidentale, Flammarion, .
- [Courtecuisse & Duhem 1994] Régis Courtecuisse et Bernard Duhem, Guide des champignons de France et d'Europe, Delachaux et Niestlé (1re éd. 1994).
- Dr_Ewaldt_Gerhardt1999">[Gerhardt 1999] Dr Ewaldt Gerhardt (trad. de l'allemand par Michel Cuisin), Guide Vigot des champignons : plus de 1200 espèces, plus de 1000 photos couleur [« Der Grosse BLV Pilzführer für unterwegs »], Paris, éd. Vigot, , 714 p. (ISBN 2-7114-1413-2 et 978-2-711-41413-0, BNF 37177286).
- [Jordan & Wheeler 1996] Peter Jordan et Steven Wheeler, Larousse saveurs - Les champignons, éds. Larousse, (ISBN 2-03-516003-0).
- [Laessoe & Del Conte] Thomas Læssøe et Anna Del Conte (trad. de l'anglais), L'Encyclopédie des champignons, Paris, éd. Bordas, coll. « Dorling Kindersley Book », , 256 p. (ISBN 2-04-027177-5).
- [Phillips 1982] Roger Phillips (trad. Odile Ricklin et Samuel Grosman), Les champignons, Paris, Solar, , 288 p. (ISBN 978-2-263-00640-1, BNF 34738816).
- [Redeuilh et al. 2004] Guy Redeuilh, Guillaume Eyssartier, Isabelle Masson-Deblaize et Philippe Joly, Larousse des champignons, Paris, éd. Larousse, (réimpr. 2015), 408 p. (ISBN 2-03-560338-2, EAN 2000144354227, présentation en ligne).
- [Romagnesi 1970-1971] Henri Romagnesi, Petit atlas des champignons (2 vol. : vol. 1 : 1970, LXXVI p. + 148 pl. ; vol. 2 : 1971, 150 p. + 348 pl.), Paris, Bordas, (réimpr. 1978, 1983) (ISBN 2-04-007940-8 et 204007970X, présentation en ligne).
- [Vajda 2012] (en) Vivi Vajda, « Fungi, a Driving Force in Normalization of the Terrestrial Carbon Cycle Following the End-Cretaceous Extinction », dans J.A. Talent (éd.), Earth and Life (International Year of Planet Earth), Springer, , sur academia.edu (DOI 10.1007/978-90-481-3428-1_27, lire en ligne).
Articles connexes
- Champignon
- Mycologie
- Mycorhize
- Sporophore
- Histoire évolutive des champignons (en)
Liens externes
- Ressources relatives au vivant :
- AlgaeBase
- Global Biodiversity Information Facility
- (da + en) Danmarks svampeatlas
- (sv) Dyntaxa
- (en) EPPO Global Database
- (en) Paleobiology Database
- (mul + en) iNaturalist
- (en) Index Fungorum
- (en) MycoBank
- (en + en) New Zealand Organisms Register
- (nl + en) Plant Parasites of Europe
- (en) Système d'information taxonomique intégré
- (en) World Register of Marine Species
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Société Mycologique de France
- Société Mycologique du Nord de la France. Site assez complet sur les Champignons et leur étude.
- Atlas des champignons, véritable encyclopédie des champignons
- Le memento des champignons
- Le projet Tree of life (en anglais)
- Champignons De Franche-Comté
- MycoDB : Base de données de champignons
- Société Mycologique de Strasbourg
- Reconnaissance visuelle des champignons
- Champignons forestiers (Suisse, PDF)