اكتشف العلماء الذين يعملون في تخصصات مختلفة على مدى سنين أدلة على السلف المشترك لبعض الكائنات الحية. وقد أثبتت هذه الأدلة حدوث التطور وقدمت معلومات وافرة حول العمليات الطبيعية التي تطورت بها الحياة بأنواعها واختلافها على كوكب الأرض. هذه الأدلة تدعم الاصطناع التطوري الحديث، وهو النظرية العلمية التي تشرح كيفية تغير الحياة مع مرور الزمن وأسباب ذلك. يوثق علماء الأحياء التطوريون حقيقة الأصل المشترك بوضع تنبؤات يمكن اختبارها، والتحقق من الفرضيات، وتطوير نظريات تمثل لها وتصف أسبابها.
كشفت مقارنات سلاسل الحمض النووي للكائنات عن أن الكائنات الأقرب وراثيًا تتشابه بدرجة أكبر في سلاسلها عن الكائنات الأبعد وراثيًا. جاءت أدلة أخرى على الأصل المشترك من المخلفات الجينية كالمورثات الكاذبة، وهي مناطق في حمض نووي ريبوزي منقوص الأكسجين تتنادّ مع جين في موجود في متعضية قريبة، لكنها لم تعد فعالة ويبدو أنها تسير في عملية انحلال مستمرة.
الأحافير مهمة لتقدير زمن تطور سلالات مختلفة في الزمن الجيولوجي. وبما أن التحجر حادثة نادرة تتطلب عادة وجود أجزاء صلبة في جسد الكائن وموته قرب موقع تخزَّن فيه الرواسب، فإن سجل الأحافير لا يقدم إلا معلومات قليلة ومتقطعة حول تطور الحياة. أدلة تواجد المتعضيات قبل تطور أجزاء الجسد الصلبة كالأصداف والعظام في غاية الندرة، لكنها موجودة في صورة أحافير دقيقة قديمة، إضافة إلى انطباعات لأجساد مختلف الكائنات لينة الجسد. دراسة التشريح المقارن لمجموعات من الحيوانات تظهر خصائص بنيوية متماثلة أو متناددة، مما يبين العلاقات الوراثية مع متعضيات أخرى، خاصةً عند مقارنتها بأحافير لمتعضيات قديمة منقرضة. تعتبر التركيبات الأثارية ومقارنات النمو الجنيني عوامل مهمة في تحديد مدى التشابه التشريحي بالتوافق مع الأصل المشترك. وبما أن العمليات الأيضية لا تخرج من الأحافير، فإن البحث في تطور العمليات الخليوية البسيطة يجري إلى حد بعيد بمقارنة فسيولوجيا المتعضيات الحالية وكيميائها الحيوية. تفرع العديد من السلاسل في مراحل مختلفة من النمو، ولذلك فإنه من الممكن تحديد زمن ظهور عمليات أيضية محددة بمقارنة صفات أفراد سلالات السلف المشترك. كما يُظهر التنظيم الحيوكيميائي المشترك وأنماط التنوع الجزيئي في المتعضيات علاقة مباشرة باشتراك السلف.
يقدم فرع الجغرافيا الحيوية المزيد من الأدلة؛ لأن التطور والسلف المشترك يقدمان أفضل التفسيرات وأكثرها شمولًا لعدد كبير من الحقائق المتعلقة بالتوزيع الجغرافي للنباتات والحيوانات حول العالم. يتجلى هذا أكثر في حقل الجغرافيا الحيوية للجُزر. تتيح حقيقة السلف المشترك مع تكتونيات الصفائح طريقة لجمع حقائق توزيع أنواع الأحياء حاليًا مع أدلة السجل الأحفوري لتقديم تفسير متسق منطقيًا لكيفية تغير التوزيع الجغرافي للكائنات الحية عبر الزمن.
تطور البكتيريا المقاومة للمضادات الحيوية وانتشارها، وانتشار أنواع النباتات والحشرات المقاومة للمبيدات، يقدمان دليلًا على أن التطور بالاصطفاء الطبيعي عملية مستمرة في العالم الطبيعي. كما أنه قد لوحظت أمثلة لانفصال تجمعات من نوع واحد إلى عدة أنواع جديدة (الانتواع). لوحظ الانتواع كذلك بطريقة مباشرة وغير مباشرة في المختبر وفي الطبيعة. وقد وُصفت عدة من تلك وقائع ووُثقت أمثلةً لأنماط معينة من الانتواع. علاوة على ذلك، فإن الأدلة على السلف المشترك تمتد من التجارب المعملية المباشرة بالاصطفاء الصناعي للمتعضيات حاليًا وفي السابق، إضافة إلى تجارب أخرى مضبوطة تشمل كثيرًا من مواضيع هذا المقال. يشرح هذا المقال الأنواع المختلفة لأدلة التطور من سلف مشترك، ويسرد أمثلة محددة كثيرة لكل من تلك الأدلة.
أدلة من علم وظائف الأعضاء المقارن والكيمياء الحيوية المقارنة
علم الوراثة

دراسة سلاسل المورثات تقدم أحد أقوى الدلائل على الأصل المشترك. يفحص تحليل المتسلسلات المقارن العلاقة بين سلاسل دنا لدى الأنواع المختلفة،[1] مما يقدم عدة أدلة تبرهن فرضية دارون الأصلية بالأصل المشترك. إن كانت فرضية الأصل المشترك صحيحة، فستكون الأنواع التي تشترك في السلف قد ورثت سلاسل دنا من ذلك السلف المشترك إضافة إلى الطفرات التي تميز ذلك السلف. وستشترك الأنواع الأكثر قرابة في جزء أكبر من سلاسل دنا المتطابقة عن ما إذا قورنت بأنواع تصلها بها قرابة أضعف.
أقوى هذه الأدلة وأبسطها يأتينا من الاستبناء الوراثي. عادةً ما تكون تلك الاستبناءات قوية، خاصةً عند استخدام سلاسل بروتين بطيئة التطور في إنشائها، ويمكن استخدامها لاستبناء جزء كبير من التاريخ التطوري للمتعضيات الحديثة، بل والتاريخ التطوري للمتعضيات المنقرضة كما في السلاسل الوراثية المستخرجة من الماموث، والنياندرتال، والتيرانوصور. تثبت تلك العلاقات المستبناة وراثيًا استبناءات العلاقات التي تعتمد على الدراسات التشكّلية أو الحيوكيميائية. أجريت أكثر الاستبناءات دقة في دراسة جينوم الميتوكوندريا الذي تشترك فيه كل المتعضيات حقيقيات النوى، وهو جينوم قصير وسهل الدراسة. أما أكثر الاستبناءات اتساعًا فقد أجريت اعتمادًا على سلاسل بضعة بروتينات قديمة جدًا أو على سلاسل رنا الريبوسومي.
تمتد العلاقات الوراثية كذلك إلى مجموعة كبيرة من العناصر التسلسلية غير الوظيفية كالتكرارات، والينقولات، والمورثات الكاذبة، والطفرات في السلاسل المرمّز للبروتين والتي لا تغير في سلسلة الأحماض الأمينية. بالرغم من أن قلة من هذه العناصر يمكن أن تكتشف لها وظائف لاحقًا، إلا أنها في الجملة توضح أن التطابق لابد أنه نتيجة الأصل المشترك وليس الوظيفة المشتركة.
التنظيم الحيوكيميائي الشامل وأنماط التنوع الجزيئي
تعتمد كل المتعضيات المعاصرة المعروفة على العمليات الحيوكيميائية ذاتها: ترمَّز المعلومات الوراثية في صورة حمض نووي (دي أن إيه، وآر أن إيه في الفيروسات)، وتنتسَخ إلى رنا، ثم تترجَم إلى بروتينات (أي مبلمرات من الأحماض الأمينية) بواسطة ريبوسومات محافظة. يتضح ذلك أكثر عند ملاحظة أن الشفرة الجينية ("مفتاح الترجمة" بين الدنا والأحماض النووية) موحدة في كل المتعضيات تقريبًا؛ أي أنه إن وُجدت قطعة دنا في بكتيريا فإنها ترمّز للحمض الأميني نفسه الذي ترمز له إن وجدت في خلية بشرية. ثلاثي فوسفات الأدينوسين هو عملة الطاقة لدى كل أشكال الحياة المعاصرة. كما أن علم الأحياء التطوري التنموي يظهر أن اشتراك التشكل هو في الواقع نتيجة لاشتراك العناصر الوراثية.[2] مثلًأ: على الرغم من اعتقاد أن العين شبيهة الكاميرا تطورت مرات عديدة منفصلة،[3] إلان أنها تتقاسم مجموعة مشتركة من البروتينات مستشعرة الضوء (أوبسينات)، مما يدل على نقطة أصل مشتركة لكل الكائنات المبصرة.[4][5][6] من الأمثلة الأخرى الجديرة بالذكر خطة جسم الكائنات الفقارية المعروفة، والتي تحدد بنيتها عائلة العلبة المثلية (Hox) من المورثات.
سَلسَلة الحمض النووي
المقال الرئيسي: سلسلة الحمض النووي
تتيح مقارنة سلاسل دنا تجميع المتعضيات حسب تشابه سلاسلها، وعادة ما تتطابق أشجار التطور مع التصنيف التقليدي، وكثيرًا ما تستخدم العلاقات التطورية لإثبات التصنيف أو تصحيحه. تعَد مقارنة السلاسل مقياسًا قويًا بما يكفي لتقويم الافتراضات الخاطئة في شجرة التطور إذا قلت الأدلة الأخرى. مثلًا: تتباين سلسلة دنا الإنسان بمقدار ما يقارب 1,2% عن أقرب أقربائه الوراثيين، الشمبانزي، وبقرابة 1,6% عن الغوريلا، و6,6% عن البابون.[7][8] لذلك يتيح دليل السَلسلة الوراثية استنتاج مدى القرابة الوراثية وتقديرها بين الإنسان والقردة الأخرى.[9][10] استُخدمت سلسلة مورثة رنا الريبوسومي 16 س (مورثة أساسية ترمّز لجزء من الريبوسوم) للعثور على علاقات وراثية واسعة بين كل الأحياء المعاصرة. أجرى ذلك البحث أولًا كارل ووز عام 1977،[11][12] وكانت نتيجة ذلك البحث إيجاد النظام ثلاثي النطاقات الذي ينص على أنه حدث انقسامان رئيسيان في بداية تطور الحياة؛ نتج عن الانقسام الأول البكتيريا الحديثة وعن الانقسام الآخر نتجت العتائق وحقيقيات النوى.
الفيروسات القهقرية الداخلية
الفيروسات القهقرية الداخلية (ERVs) هي سلاسل في الجينوم بقيت من عدوى فيروسية قديمة في المتعضية. دائمًا ما توَرَّث تلك الفيروسات القهقرية إلى الجيل التالي من المتعضية المصابة. هذا يحافظ على المورثة الفيروسية في جينوم المتعضية. ولأن هذا الحدث نادر وعشوائي، فإن العثور على مورثات فيروسية في موضعين متماثلين على صبغيين من نوعين مختلفين من المتعضيات يشير إلى سلف مشترك بينهما.[13] انظر أمثلة ذلك في الإنسان وفي القطط أدناه.
البروتينات
تدعم الأدلة البروتيومية شمولية أصل الحياة. البروتينات الحيوية كالريبوسوم، والدنا بولمريز والرنا بولمريز توجد في كل الكائنات بدءًا بأكثر البكتيريا بدائية وحتى أكثر الثدييات تعقيدًا. الجزء الرئيسي في البروتين محفوظ في كل سلالات الحياة ويؤدي وظائف متشابهة. طورت المتعضيات الأكثر تعقيدًا وحدات بروتين إضافية، مما يؤثر بقوة على تنظيمه وتآثرات البروتين-بروتين في مركزه. التشابه المترابط بين سلاسلات كل المتعضيات المعاصرة؛ مثل الدنا، والرنا، والأحماض الأمينية، وطبقة الدهن الثنائية تدعم كلها نظرية السلف المشترك. التحليل الوراثي للسلاسل البروتينية الموجودة في متعضيات متعددة ينتج أشجار علاقة وراثية متشابهة.[14] التماكب الضوئي للدنا، والرنا، والأحماض الأمينية محفوظ من الأصل المشترك في كل أشكال الحياة المعروفة. وبما أنه لا توجد أفضلية وظيفية لليدوانية التيأمنية أو التياسرية فإن أبسط فرضية هي أن الاختيار وقع عشوائيًا في المتعضيات القديمة ووَرِثت ذلك إلى كل أشكال الحياة المعاصرة من خلال سلفها المشترك. يأتي مزيد من الأدلة لاستبناء السلالات الوراثية من فضلة الدنا مثل المورثات الكاذبة، وهي مورثات "ميتة" تتراكم فيها الطفرات بانتظام.[15]
المورثات الكاذبة
المقال الرئيسي: مورثة كاذبة المورثات الكاذبة (المعروفة أيضًا باسم الدنا غير المشفِّر) هي سلاسل دنا في الجينوم ولا تُنسخ إلى رنا لتصنيع البروتينات. بعض هذا الدنا غير المشفر له وظائف معروفة، ولكن أكثره ليست له وظيفة معروفة ويسمى "فضلة الدنا". هذا مثال على الأثارية؛ لأن مضاعفة هذه المورثات تستهلك طاقة، مما يجعل الأمر إهدارًا tي كثير من الحالات. تشكل المورثات الكاذبة 99% من الجينوم البشري (1% دنا وظيفي).[16] يمكن أن تنتج مورثة كاذبة عندما تحدث طفرة لمورثة مشفِّرة وتمنع نسخه، مما يعطل وظيفته، لكن لأنه لا يُنسخ فيمكن أن يختفي دون أن يؤثر على صلاح المتعضية إلا إن أوجد ذلك وظيفة جديدة نافعة للدنا غير المشفر. يمكن أن تنتقل المورثات الكاذبة غير العاملة إلى الجيل التالي من النوع، وهذا يجعل من الواضح أن النوع التالي خلف للنوع الأول.
آليات أخرى
توجد أيضًا أدلة جزيئية كثيرة لعدد من الآليات المختلفة للتغييرات التطورية الكبيرة، منها: الجينوم وتضاعف المورثات، الذي يسهل التطور السريع عن طريق توفير كميات كبيرة من المادة الوراثية بقيود انتقائية ضعيفة أو بغيابها. كذلك نقل الجينات الأفقي، وهي عملية نقل المادة الوراثية إلى خلية أخرى ليست من ذرية المتعضية، مما يتيح للأنواع اكتساب مورثات نافعة من بعضها. إضافةً إلى التأشيب الذي يسمح بإعادة ترتيب أعداد كبيرة من الألّيلات المختلفة ويسمح بإقامة انعزال تكاثري. تشرح نظرية التعايش الداخلي أصل الميتوكوندريا والصانعات الخلوية (مثل الصانعات اليخضورية)، وهي عضيات في خلايا حقيقيات النوى، على أنه دمج لخلية عتيقة بدائية النواة في خلية عتيقة حقيقية النواة. تضع هذه النظرية آلية لقفزات تطورية مفاجئة بدمج المادة الوراثية والتركيب الحيوكيميائي لنوعين مختلفين بدلًا من تطور عضيات حقيقيات النوى ببطء. وُجدت ادلة تدعم هذه النظرية في الطلائعية Hatena arenicola؛ فهي تفترس خلية طحلب أخضر وتبتلعها، فتتصرف خلية الطحلب الأخضر كمتعايش داخلي يغذي Hatena التي تفقد جهازها الغذائي وتتصرف ككائن ذاتي التغذية.[17][18]
بما أن العلميات الأيضية لا تغادر الأحافير، فإن البحث في تطور العمليات الخليوية البسيطة يجري إلى حد بعيد بمقارنة المتعضيات المعاصرة. تفرعت سلالات عديدة مع ظهور عمليات أيضية جديدة، ويمكن -نظريًا- تحديد زمن ظهور عمليات أيضية محددة بمقارنة صفات ذرية السلف المشترك أو بملاحظة التظاهرات الفيزيائية لتلك الصفات. على سبيل المثال، ظهور الأكسجين في غلاف الأرض الجوي مرتبط بتطور التمثيل الضوئي.
أمثلة محددة
الصبغي 2 في الإنسان
- مقالة مفصلة: صبغي 2 (إنسان)
يمكن مشاهدة أحد أدلة تطور الإنسان من سلف مشترك مع الشمبانزي بالنظر إلى عدد صبغيات الإنسان مقارنةً ببقية القردة العليا؛ فكل القردة العليا باستثناء الإنسان تملك 24 زوجًا من الصبغيات، أما الإنسان فلديه 23 زوجًا فقط. الصبغي الإنساني رقم 2 هو نتيجة التحام طرفي بين صبغيين سلفيين.[19][20]
من أدلة ذلك ما يلي:
- توافق الصبغي الإنساني 2 مع صبغيين لدى بقية القردة. أقرب أقرباء الإنسان -الشمبانزي العام- لديه سلاسل دنا شبه مطابقة لتلك الموجودة في الصبغي 2 لدى الإنسان، لكنها موجودة لدى الشمبانزي في صبغيين منفصلين. الأمر ذاته ينطبق على الأقرباء الأبعد من ذلك كما في الغوريلا والأورانغوتان.[21][22]
- وجود مشطر أثاري. المعتاد أن يكون في كل صبغي مشطر واحد فقط، لكن في الصبغي 2 توجد بقايا صبغي آخر.[23]
- وجود تيلوميرين أثاريين. عادة ما تكون التيلوميرات موجودة في نهايتي الصبغي فقط، ولكن في الصبغي 2 توجد سلسلتا تيلومير إضافيتين في المنتصف.[24]
لذلك فإن الصبغي 2 يقدم أدلة قوية تدعم اشتراك السلف بين الإنسان والقردة الأخرى. يقول ي. ف. أيدو: "نستنتج أن الموقع المستنسخ في الكوزميدين c8.1 وc29B هو أثر التحام تيلوميري-تيلوميري عتيق ويشير إلى اللحظة التي التحم فيها صبغيان قرديان ليعطيا الصبغي الإنساني 2"."[24]
الأصل الأفريقي للإنسان الحديث
- مقالة مفصلة: فرضية المنشأ الحديث الموحد
- طالع أيضًا: هابلوغروب (متقدرة)
- هابلوغروب (صبغي Y)
النماذج الرياضية للتطور التي كان من روادها أمثال سيوال رايت، ورونالد فيشر، وجون هالدين، والتي وسّعتها نظرية الانتشار التي وضعها كيمورا موتو، تتيح تلك النماذج التنبؤ بالبنية الوراثية للتجمعات التي تخضع للتطور. كما أن الاختبار المباشر للبنية الوراثية في التجمعات الحديثة عن طريق سلسلة الدنا قد أتاح للعلماء التحقق من كثير من تلك التنبؤات. على سبيل المثال: نظرية المنشأ الأفريقي للإنسان، والتي تقول بأن الإنسان الحديث تطور في أفريقيا وهاجر تجمع صغير من أفراده إلى القارات الأخرى (في ما يمثل عنق زجاجة سكانية)، تعني هذه النظرية ضمنيًا أنه لابد أن نجد في التجمعات الحديثة علامات ذلك النمط من الهجرة. وتحديدًا، فإنه يفترض أن نجد في التجمعات التي تلت عنق الزجاج (الأوروبيون والآسويون) مستويات أقل في التنوع الوراثي عمومًا وأن نلاحظ توزيعًا أكثر انتظامًا في تكرارات الأليلات عند مقارنتهم بالتجمع السكاني الأفريقي. ثبتت صحة التنبؤين السابقين ببيانات حقيقية من مجموعة من الدراسات.[25]
الفيروسات القهقرية الداخلية في الإنسان
يحمل الإنسان الكثير من الفيروسات القهقرية التي تمثل ما يقارب 8% من جينومه.[26] يشترك الإنسان والشمبانزي في سبع مورثات فيروسية مختلفة، وتشترك الرئيسيات كلها في فيروسات قهقرية مشابهة تنسجم مع علم الوراثة العرقي.[27]
الفيروسات القهقرية الداخلية في القطط
تقدم السنوريات مثالًا آخر على سلسلة مورثة فيروسية وُجدت في سلف مشترك. تحتوي الشجرة التطورية للسنوريات قططًا صغيرة (مثل سنور الأدغال، والقط البري، والقط المنزلي) تفرعت عن أنواع القطط الكبيرة مثل تحت عائلة النمرية ولواحم أخرى. حقيقة وجود مورثة فيروسية داخلية لدى القطط الصغيرة حيث لا توجد في القطط الكبيرة تشير إلى أن تلك المورثة أُضيفت في جينوم سلف القطط الصغيرة بعد تفرع القطط الكبيرة عنها.[28]
أدلة من علم التشريح المقارن
تكشف الدراسة المقارنة لتشريح مجموعات من الحيوانات أو النباتات عن تشابه عام في خصائص بنيوية معينة. مثلًا: البنية الأساسية لكل الأزهار تتكون من سبلات، وبتلات، ومبيض وقلم وميسم؛ إلا أن أحجام الأزهار، وألوانها، وأعداد أجزائها، وبنياتها الخاصة متباينة بين الأنواع.
التأسل الرجعي
- مقالة مفصلة: تأسل رجعي
التأسل هو الرجعية التطورية، مثل ظهور صفات على متعضية مرة أخرى بعد أن انحسرت منذ أجيال.[29] يقع التأسل لأن مورثات الخصائص الظاهرية التي وجدت في السابق ما زالت محفوظة في الدنا، وإن كانت تلك المورثات لا يعبَّر عنها ظاهريًا في أغلب المتعضيات التي تحوزها.[30] من أمثلة ذلك ظهور الأرجل الخلفية في الثعابين[31] أو الحيتان[32] (انظر الأمثلة المحددة أدناه)، وأصابع الأقدام الزائدة لدى الحافريات والتي لا تصل إلى الأرض أصلًا،[33] ووجود أسنان للدجاج،[34] وعودة التكاثر الجنسي في ينم وCrotoniidae بعد اختفائه،[35] وظهور ذيل للإنسان،[29] وظهور حلمات زائدة،[31] وتضخم الأنياب.[31]
علم الأحياء التطوري التنموي والنمو الجنيني
- مقالة مفصلة: علم الأحياء التطوري التنموي
- طالع أيضًا: تخلق جنيني
علم الأحياء التطوري التنموي هو فرع علم الأحياء الذي يعنى بمقارنة عملية النمو في متعضيات مختلفة لتحديد العلاقات الوراثية بين أنواعها. تحتوي جينومات مجموعة متنوعة من المتعضيات جزءًا صغيرًا من المورثات التي تتحكم في نمو المتعضية. تعد مورثات العلبة المثلية من أمثلة تلك المورثات الشاملة في المتعضيات كلها تقريبًا والتي تشير إلى أصل السلف المشترك. يظهر الدليل الجنيني من دراسة نمو المتعضيات في طورها الجنيني ومقارنته بأجنة متعضيات أخرى وملاحظة التشابه بينها. عادة ما تظهر بقايا صفات سلفية وتختفي أثناء مراحل النمو الجنيني المختلفة. من أمثلة ذلك إنماء الشعر وفقدانه (زغب) أثناء نمو جنين الإنسان،[36] ووجود التحول الظاهري أثناء النمو الجنيني لدى الثدييات من مظهر الأسماك إلى البرمائيات إلى الزواحف وحتى الثدييات، ونمو الكيس المحّي ثم تنكّسه، وكون الضفادع والسمندلات البرية تمر بطور اليرقة -التي تتصف بصفات اليرقات المائية- داخل البيضة ولكنها تفقس جاهزة للحياة على اليابسة،[37] وظهور تركيبات خيشومية الشكل (الأقواس البلعومية) أثناء نمو الجنين الفقاري، والتي تستمر في النمو لدى الأسماك في صورة أقواس خيشومية حيث تشكل منشأ عدد من التركيبات في الرأس والعنق تختلف عن التي تنتجها الأقواس البلعومية في الإنسان مثلًا.
التنادد البنيوي والتطور التباعدي (التكيفي)
إن كانت مجموعتان متباعدتان من المتعضيات قد انحدرتا من أصل مشترك فمن المتوقع أن يكونا مشتركتين في خصائص أساسية معينة. يفترض أن تحدد درجة التشابه بين متعضيتين مدى قرابتهما تطوريًا:
- يُفترض أن المجموعات التي تتشارك صفات قليلة قد تفرعت عن أصل مشترك في زمن أقدم في التاريخ الجيولوجي عن تلك التي تتشابه كثيرًا.
- عند تقدير مدى القرابة التطورية بين حيوانين، يبحث عالم الأحياء المقارن عن البنى المتشابهة جوهريًا، حتى وإن كان لها وظائف مختلفة في الحيوان البالغ. تسمى هذه البنى بالمتناددة، وهي تشير إلى اشتراك الأصل.
- في حالات تشابه بنيتين واختلاف وظائفهما قد يكون من الضروري تتبع أصلهما ونموهما الجنيني. تشابه الأصل التنموي يشير إلى تطابق البنيتين، وعليه ترجيح أنهما مشتقتان من سلف مشترك.
عندما تتشارك مجموعة من المتعضيات في بنية متناددة تؤدي وظائف متعددة للتكيف مع ظروف بيئية وأنماط حياة مختلفة فإن ذلك يسمى التشعب التكيفي. ويسمى الانتشار التدريجي للمتعضيات ذات التشعب التكيفي بالتطور التباعدي.
التصنيف والمراتب المتداخلة
يعتمد التصنيف على حقيقة أن المتعضيات كلها مرتبطة وراثيًا في مراتب متداخلة مبنية على أساس الخصائص المشتركة. يمكن توزيع أغلب الأنواع المعاصرة في تصنيف مراتبي متداخل بسهولة. يتضح هذا من نظام التصنيف اللينيوسي. يمكن وضع الأنواع القريبة في مجموعة واحدة (كالجنس) اعتمادًا على الخصائص المشتركة بينها، ويمكن أن تُجمع عدة أجناس في فصيلة واحدة، وأن تجمع عدة فصائل معًا في رتبة واحدة...إلخ.[38] اكتشف كثير من العلماء وجود هذه المراتب المتداخلة قبل دارون، ولكنه وضح أن نظريته التطورية بسلفها المشترك ونمطها المتشعب تستطيع تفسير تلك المراتب.[38][39] وصف دارون كيف يمكن أن يكون اعتماد الأصل المشترك قاعدة منطقية للتصنيف:[40]
| " | إذا لم أكن في غاية الوهم، فإن كل ما سبق من قواعد وتسهيلات وصعوبات تصبح واضحة على أساس أن النظام الطبيعي مبني على التناسل مع التحوّر، وعلى أن الخصائص التي يرى الطبائعيون أنها تظهر قرابة حقيقية بين نوعين أو أكثر هي التي وُرثت عن سلف مشترك، وعلى ذلك فإن التصنيف الحقيقي تصنيف نَسَبي، وإن اشتراك السلف هو الرابطة الخفية التي يبحث عنها الطبائعيون لاشعوريًا... | " |
—تشارلز دارون، أصل الأنواع، صفحة 577 | ||
البنى الأثارية
- مقالة مفصلة: أثارية
- طالع أيضًا: الأثارية في الإنسان
تظهر أدلة قوية ومباشرة على السلف المشترك من دراسة البنى الأثارية.[41] تطلق تسمية العضو الأثاري على الأجزاء الجسدية الردمية التي تمتاز بصغر الحجم والضعف لدى مقارنتها بنظيرتها في نوع السلف. عادة ما تكون هذه الأعضاء ضعيفة ونموها متخلف. يمكن تفسير وجود الأعضاء الأثارية باعتبار التغيرات البيئية المحيطة بالنوع أو تغيرات نمط حياته. تكون تلك الأعضاء عاملة عادة في النوع السلفي ولكنها فقدت وظيفتها أو غيرتها. من أمثلة ذلك الحزام الحوضي لدى الحيتان، والأثقال (الأجنحة الخلفية) في الذباب والبعوض، وأجنحة الطيور غير الطائرة كالنعام، وأوراق بعض النباتات الصحراوية (مثل الصبار) والنباتات الطفيلية (مثل الحامول). إلا أن البنى الأثارية يمكن أن تستبدل وظائفها بوظائف جديدة. مثلًا: أثقال ذوات الجناحين تساعد في موازنة الحشرة أثناء طيرانها، وجناحا النعامة يستخدمان في طقوس التزاوج.
| " | أكثر الاستنتاجات منطقية هي أن هذه المخلوقات انحدرت من مخلوقات كانت فيها تلك الأجزاء عاملة، مما يشير إلى أن أغلب (بل جميع) المخلوقات تنحدر من أسلاف مشتركين. | " |
—ناتان سلفكن، "التحدي الخَلقي"، صفحة 262 | ||
أمثلة محددة

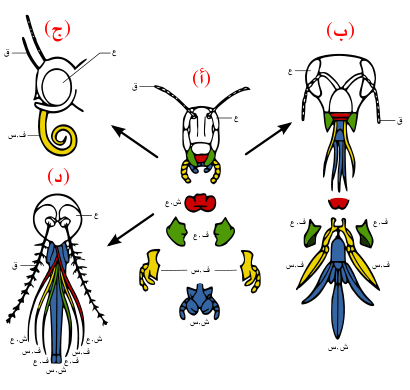
(ب) القرص والعض: مثل نحلة العسل؛ الشفة السفلى طويلة للعق الرحيق، والفكان العلويان يمضغان حبوب اللقاح ويشكّلان الشمع.
(ج) الامتصاص: مثل الفراشة؛ الشفة العيا مختزلة، والفك العلوي مفقود، والفك السفلي طويل ويكوّن أنبوبًا للامتصاص.
(د) الاختراق والامتصاص: مثل أنثى البعوض؛ الشفة العليا والفك السفلي يكونان أنبوبًا، والفكان العلويان يكوّنان إبرتين خازقتين، والشفة العليا عليها أخدود يسمح باحتواء أجزاء أخرى.



التركيبات الخلفية في الحيتان
للحيتان أجزاء خلفية داخلية مختزلة؛ كالحوض والأقدام الخلفية (شكل 3أ).[42][43] أحيانًا تكون الجينات المسؤولة عن نمو الأطراف الطويلة سببًا في نمو ساقين صغيرتين لدى حوت حديث. في يوم 28 أكتوبر 2006، عثر على دلفين بأربع زعانف ودُرس طرفاه الخلفيان الزائدان.[44] تعتبر هذه الحيتانيات ذات الأرجل مثالًا على أثارة يمكن توقعها بالنظر إلى سلفها المشترك.
تركيب فم الحشرات
تملك أنواع كثيرة من الحشرات أجزاء فم مشتقة من التركيبات الجنينية ذاتها، مما يشير إلى أن أجزاء الفم تلك هي تعديلات على الخصائص الأصلية لدى سلف مشترك. من هذه الأجزاء الشفة العليا، واللحى (الفكوك العلوية)، والبلعوم السفلي (اللسان أو قاع الفم)، والفكوك السفلية المساعدة، والشفة السفلى (شكل 3ب). أدى التطور إلى تضخم هذه التركيبات وتعديلها في بعض الأنواع، أو اختزالها وفقدانها في أنواع أخرى. تمكن تلك التعديلات الحشرات من الاستفادة من مواد غذائية متعددة ومختلفة.
أطراف مفصليات أخرى
تعتبر أجزاء أفواه الحشرات وقرون استشعارها متناظرة مع أرجلها. نلاحظ تطورًا متوازيًا في بعض العنكبيات: يمكن تعديل زوج الأرجل الأمامي ليصبح متناظرًا مع قرني الاستشعار، خاصة في العقارب السوطية التي تسير على ست أرجل. تدعم هذه التطورات نظرية أن التعديلات المعقدة تبدأ عادةً بتناسخ المكونات ثم تعديل كل نسخة على حدة في اتجاه مختلف.
الطرف خماسي الأصابع
- مقالة مفصلة: تطور الثدييات
إن نمط توزيع عظام الأطراف والمسمى الطرف خماسي الأصابع هو مثال على التركيبات المتناددة (شكل 3 ج). هذا النمط موجود في كل طوائف الفقاريات رباعية الأطراف (أي من البرمائيات وحتى الثدييات). كما يمكن تتبع أثره قديمًا في زعانف بعض الأسماك المتحجرة التي تطور منها أول البرمائيات، مثل تِكْتالِك. يوجد في الطرف عظمة واحدة دانية (عظمة العضد)، وعظمتان قاصيتان (الكعبرة والزند)، ومجموعة من عظام الرسغ (في المعصم)، وبعدها خمس عظام للمشط (عظام الكف) ثم السلاميات (عظام الأصابع). في كل رباعيات الأطراف نرى التركيب الأساسي للطرف خماسي الأصابع متطابقًا، مما يشير إلى أنها نشأت من سلف مشترك، لكن هذه التركيبات الأساسية قد عُدِّلت أثناء مسيرة التطور. أصبحت ظاهريًا تركيبات مختلفة وليست ذات علاقة ببعضها تقوم بوظائف مختلفة للتكيف مع البيئات وأنماط الحياة المتنوعة. نرى هذه الظاهرة بجلاء في الثدييات (شكل 3 ج)؛ مثلًا:
- في السعدان استطال الطرفان الأماميان كثيرًا ليكونا يدًا قابضة تساعد في تسلق الأشجار والتأرجح بينها.
- في الخنزير فُقد الإصبع الأول واختُزل الإصبعان الثاني والخامس. الإصبعان المتبقيان أطول وأكثر متانة من البقية ويحملان حافرًا لدعامة الجسد.
- في الحصان تكيفت الأطراف الأمامية للدعامة والجري عن طريق الاستطالة الشديدة للإصبع الثالث الذي يحمل حافرًا.
- في الخلد نجد طرفين أماميين قصيرين في صورة مجرفة للحفر والاختباء.
- آكلات النمل تستخدم إصبعها الثالث المتضخم لتحطيم تلال أوكار النمل وأعشاش الأرضة.
- في الحوت أصبح الطرفان الأماميان جدافتين (زعنفتين) لتوجيه السباحة والحفاظ على التوازن في الماء.
- في الخفاش تحول الطرفان الأماميان إلى جناحين للطيران باستطالة شديدة لأربعة أصابع، أما الإصبع الأول الذي يشبه الخطاف فبقي حرًا للتعلق والتدلي من الأشجار.
تركيب الحوض في الديناصورات
- طالع أيضًا: تطور الديناصورات
- تطور الطيور
كما في الطرف خماسي الأصابع في الثدييات، فإن الديناصورات الأولى انقسمت إلى رتبتين مختلفتين: سحليات الحوض وطيريات الحوض. تُصنف الديناصورات في إحدى الرتبتين حسب ما يظهر في أحافيرها. في الشكل 3 د نرى أن أولى سحليات الحوض كانت تشبه أولى طيريات الحوض. نمط تركيب الحوض في كل أنواع الديناصورات هو مثال على التركيبات المتناددة. كل من رتبتي الديناصورات لها عظام حوضية مختلفة قليلًا مما يقدم دليلًا على السلف المشترك لهما. كما أن أحواض الطيور الحديثة تظهر تشابهًا مع التراكيب الحوضية في سحليات الحوض مما يشير إلى تطور الطيور من الديناصورات. يمكن مشاهدة ذلك في الشكل 3 د حيث تنقسم الطيور عن التحترتبة ثيروبودا.
العصب الحنجري الراجع في الزرافات
العصب الحنجري الراجع هو الفرع الرابع من العصب المبهم الذي هو أحد الأعصاب القحفية. عادة ما يكون مساره طويلًا في الثدييات. يخرج العصب الحنجري الراجع ابتداءً من المخ جزءًا من العصب المبهم، ثم يمر في الرقبة حتى القلب، ويلتف حول الأبهر الظهري ثم يرجع صعودًا إلى الحنجرة من خلال الرقبة. (شكل 3 هـ)
ويدعي التطوريون أن هذا المسار ليس هو المسار الأقصر حتى في الإنسان، لكن في الزرافات يصبح أدنى من المستوى الأمثل بكثير. بسبب طول رقبة الزرافة فإنه يمكن أن يمتد العصب الحنجري الراجع لديها إلى 4 أمتار بالرغم من أن مساره الأقصر لا يتجاوز طوله عدة بوصات. ويعلل المسار غير المباشر لهذا العصب بأنه نتيجة تطور الثدييات من الأسماك التي لم تكن لها رقبة وكان عصبها قصير نسبيًا ويعصِّب فلعة خيشومية واحدة ويمر بالقرب من القوس الخيشومية. بعد ذلك أصبح الخيشوم الذي يصل إليه العصب حنجرةً، وأصبحت القوس الخيشومية الشريان الأبهر الظهري في الثدييات.[45][46]
أما وجهة النظر المعارضة لفكرة أن العصب الراجع دليل على التطور؛ أن هذا الاستدلال يقوم على أساس غير صحيح، وهو افتراض أن العصب يخص الحنجرة فقط وأن هذا الالتفاف لا فائدة منه، وبالتالي يعتبرونه خطأ تطوريًا، وقدم العلماء المعارضين لهذا الاستدلال شواهد تنقد الاستدلال البسيط بأنه خطأ تطوري:
- حالات عدم التفاف العصب شذوذ ضار: نشر عالم الأحياء ولف لونينغ Wolf-Ekkehard Lِnnig بحثًا بعنوان: "العصب الراجع في الزرافة: هل يثبت التطور؟! [47] قال فيه أنَّ ذلك الالتفاف لا يعني بالضرورة سوء تصميم، مستدلًا بذلك على حالات عدم التفاف العصب عند البشر (0.3 إلى 1% من البشر)، حيث يسبب عدم حصول ذلك الالتفاف عند البشر عيبًا خلقيًا من نتائجه أنه يسبب تضخمًا في الشريان تحت الترقوة اليمنى من قوس الأبهر[48].
- تفرعات العصب الحنجري ووظائفها: ادعاء التطوريون بني على أن وظيفة العصب هي فقط تعصيب الحنجرة وكأنه عصب فردي وليس أحد تفرعات العصب المبهم الكثيرة؛ فهذا الالتفاف للعصب الحنجري كما يقول مرجع التشريح غرايز أناتومي تصدر منه تفرعات لأعضاء مختلفة خلال رحلته، منها
- للضفيرة العصبية القلبية،
- وللأغشية المخاطية والطبقة العضلية للمري،
- وكذلك فهو يعصب الغشاء المخاطي والأليف العضلية للرغامى.
أي أنه لا يختص بتعصيب الرغامى فحسب [49]
- دور التفاف العصب الجنيني: أنَّ هذا الالتفاف ليس بلا فائدة كما يقول الأساس الذي بُني عليه الإشارة إلى أنه خطأ تطوري؛ إنما له دور هام في تكون القناة الشريانية في المرحلة الجنينية. فيوجه الدم المؤكسج الآتي للجنين ليوصله للقلب مباشرةً بدل وصوله للرئة [50]
مسار الأسهر
كما في عصب الحنجرة في الزرافة فإن الأسهر (جزء من التشريح الذكري لكثير من الفقاريات ينقل الحيوانات المنوية من البربخ تحسبًا للقذف) في الإنسان يصعد الأسهر من الخصية، ويلف حول الحالب، ثم ينزل إلى الإحليل والقضيب. يشير البعض إلى أن هذا بسبب هبوط الخصيتين أثناء مدة تطور الإنسان لأسباب متعلقة بدرجة الحرارة على الأرجح. ومع هبوط الخصيتين، ازداد طول الأسهر لمواءمة الانعقافة غير المقصودة حول الحالب.[46][51]
أدلة من علم الأحياء القديمة

عندما تموت المتعضيات فإنها عادة ما تحلل سريعًا أو تستهلك أجسادها قمامات، مما لا يدع أي أثر دائم لوجود تلك المتعضيات. إلا أنه في بعض الأحيان تحفظ المتعضيات. تسمى بقايا متعضيات الأزمنة الجيولوجية السابقة أو آثارها المطمورة في الصخور بآليات طبيعية، تسمى مستحاثات. المستحاثات غاية في الأهمية لفهم تاريخ تطور الحياة على الأرض، فهي تقدم أدلة مباشرة على التطور ومعلومات مفصّلة حول سلف المتعضيات. علم الأحياء القديمة يعنى بدراسة الحياة السابقة اعتمادًا على سجلات الأحافير وعلاقاتها بالأزمان الجيولوجية المختلفة.
لابد من أن تُدفن آثار المتعضيات وبقاياها سريعًا حتى لا تقع التجوية أو التحلل ومن ثم يمكن حدوث التحجّر. التركيبات الهيكلية أو الأجزاء الصلبة الأخرى هي أكثر أنواع البقايا المتحجرة شيوعًا. كما توجد بعض متحجرات الآثار في صورة قوالب أو طبعات لبعض الكائنات قديمة.
بعد موت الحيوان تبدأ المواد العضوية بالتحلل تدريجيًا حتى يصبح العظم مساميًا. إذا دُفن الحيوان بعد ذلك في الطين، فإن الأملاح المعدنية تتخلل العظام وتملأ المسام تدريجيًا. تتصلب العظام وتتحجر وتُحفظ في صورة أحافير. تعرف هذه العملية بالتحجر. إذا غمرت الحيوانات الميتة الرمال التي تحملها الرياح، ثم حولته الأمطار الغزيرة أو الفيضانات إلى طين، فإن عملية دخول المعادن نفسها قد تحدث. إلى جانب التحجر، فإن أجساد الحيوانات الميتة يمكن أن تُحفظ جيدًا في الجليد، أو في راتنج الأشجار المخروطية المتجمد (الكهرمان)، أو في القطران، أو في الخث الحمضي اللاهوائي. يمكن أن تكون الأحفورة لأثر مطبوع. ومن ذلك أوراق الأشجار وآثار خطوات الأقدام، التي تتكون أحافيرها طبقة طبقة ثم تتصلب.
سجل الأحافير

يمكن معرفة كيف تطورت مجموعة معينة من المتعضيات بواسطة ترتيب سجلها الأحفوري زمنيًا. يمكن تحديد هذا الترتيب لأن الأحافير توجد غالبًا في صخور رسوبية. يتكون الصخر الرسوبي من طبقات من الطمي أو الطين بعضها فوق بعض؛ ولذلك تحتوي الصخور الناتجة عن ذلك مجموعة من الطبقات الأفقية تسمى الطبقات الأرضية. كل طبقة تحتوي أحافير نموذجية للعصر المعين الذي تكونت فيه. أكثر الطبقات انخفاضًا تحتوي أقدم الصخور وأول الأحافير، بينما أعلى الطبقات تحتوي أحدث الصخور وأحافير أحدث.
يمكن أيضًا مشاهدة تتابع الحيوانات والنباتات في اكتشافات الأحافير. بعد دراسة أعداد الأحافير المختلفة ومدى تعقيدها في مستويات طبقية مختلفة، لوحظ أن الصخور الأقدم تحوي أحافير أقل، وكلها لكائنات لها تراكيب أبسط، في حين أن الصخور الأحدث فيها تنوع أكبر من الأحافير، مع زيادة متصاعدة في مستوى تعقيد تراكيبها.[52]
كان علماء الأرض قادرين على تقدير أعمار الطبقات والأحافير التي يجدونها بشكل تقريبي فقط لسنوات كثيرة. كانوا يفعلون ذلك بتقدير زمن تكون الصخر الرسوبي طبقة بطبقة مثلًا. أما اليوم، فيمكن للعلماء تحديد أعمار الأحافير بدقة أعلى مستخدمين نِسب العناصر المستقرة والمشعة في صخرة معينة. تُعرف هذه التقنية بالتأريخ الإشعاعي.
طوال السجل الأحفوري، فإن كثيرًا من الأنواع التي تظهر في مستوى طبقي أقدم تختفي في مستوى أحدث. يُفسَّر هذا بالتعبير التطوري على أنه محدد للأزمان التي نشأت فيها الأنواع أو انقرضت. تنوعت المناطق الجغرافية والأحوال الجوية على مدى تاريخ الأرض. وبما أن المتعضيات متكيفة لبيئات معينة، فأن الأحوال دائمة التغير تفضّل الأنواع التي تتكيف للبيئات الجديدة على غيرها، وذلك بواسطة آلية الاصطفاء الطبيعي.
امتداد السجل الأحفوري
- طالع أيضًا: أحفورة انتقالية
- قائمة الأحافير الانتقالية


بالرغم من ندرة توافر الأحوال المناسبة للتحجر، إلا أنه يُعرف ما يقارب 250 ألف نوع أحفوري.[54] عدد الأحافير يختلف كثيرًا من نوع لآخر، لكن ملايين الأحافير قد استُخرجت، مثلًا: استُخرج أكثر من ثلاثة ملايين أحفورة تعود إلى العصر الجليدي الأخير من بحيرة قطران لا بريا في لو أنجلس، الولايات المتحدة الأمريكية.[55] ما زال هنالك الكثير من الأحافير في الأرض في تشكيلات جيولوجية مختلفة معروف أنها ذات كثافة أحفورية عالية، مما يسمح بتقدير مجموع المحتوى الأحفوري في التشكيل. مثال على ذلك تشكل بوفورت في جنوب أفريقيا الغني بأحافير الفقاريات، ومنها الزواحف شبيهة الثدييات (وهي مجموعة أنواع انتقالية بين الزواحف والثدييات).[56] قُدر محتوى هذا التشكل بما يقارب 800 مليار أحفورة فقارية.[57]
قصور السجل الأحفوري
السجل الأحفوري مصدر مهم للعلماء عند تتبع التاريخ التطوري للمتعضيات، ولكن لوجود عجز متأصل في طبيعة السجل فإنه لا توجد تدرجات دقيقة من الأشكال الانتقالية بين المجموعات المتقاربة من الأنواع. يعد هذا الافتقار إلى سلسلة متتابعة من الأحافير في السجل أحد أكبر القيود التي تحول دون تتبع سلف المجموعات الحيوية من خلال السجل الأحفوري. تطلق تسمية "الحلقة المفقودة" عادةً في إطار غير علمي على الأحافير الانتقالية التي تُظهر هيئات متوسطة بين نوعين وكانت فجوة معرفية قبل العثور عليها.
توجد فجوة بحجم يقارب مئة مليون سنة بين بداية العصر الكامبري ونهاية العصر الأردفيشي. أوائل العصر الكامبري هي الفترة التي يوجد فيها الكثير من أحافير الإسفنجيات، واللاسعات (مثل قناديل البحر)، وشوكيات الجلد (مثل الزنبقيات الأولية، والرخويات (مثل الحلزونات)، ومفصليات الأرجل (مثل ثلاثيات الفصوص). قُدّر تاريخ وجود أول حيوان لديه الخصائص النموذجية للفقاريات (مدرعة أراندا) بأواخر العصر الأردفيشي. لذلك لم يعثر إلا على قليل من الأحافير المتوسطة بين اللافقاريات والفقاريات، إلا أن الأحافير المرشحة لمثل ذلك الدور تشمل حيوان تشكّل صخور برجس الطينية، والبيكايا[58] وأقرباؤه في صخور ماوتيانشان الطينية، وسمكة كونمينغ، وحيوان يونان، والهايكويلة السهمية،[59] وسمكة هايكو.[60]
من أسباب نقص سجلات الأحافير:
- احتمال تحجّر منعضية ما ضئيل جدًا بوجه عام.
- احتمال تحجّر بعض الأنواع أو المجموعات أقل من ذلك بسبب أجسادها اللينة والرخوة.
- احتمال تحجر بعض الأنواع أو المجموعات أقل من ذلك بسبب أنها تعيش (وتموت) في أحوال غير مؤاتية للتحجر.
- الكثير من المتحجرات قد دمرت بفعل عوامل التعرية وتكتونيات الصفائح.
- أغلب الأحافير متكسرة.
- بعض التغيرات التطورية تحدث في تجمعات تعيش في أقصى مدى جغرافي لتحمل النوع للعوامل البيئية؛ وبما أن تلك التجمعات غالبًا ما تكون صغيرة فإن احتمالية التحجّر تقل. (انظر التوازن المتقطع)
- كما أنه عند تغير الظروف البيئية، فإنه يرجح أن يقل كثيرًا حجم تجمع نوع ما من المتعضيات بحيث تقل احتمالية تحجر أي تغير تطوري سببته هذه الظروف البيئية الجديدة.
- تحمل معظم الأحافير معلومات عن الهيئة الخارجية، لكنها لا توضح الكثير عن طريقة عمل المتعضية.
- عند الاستناد إلى مدى التنوع الحيوي في الحاضر، فإن هذا يوحي بأن الأحافير المستخرجة لا تمثل سوى جزء ضئيل من العدد الكبير من أنواع المتعضيات التي كانت تعيش في الماضي.
أمثلة محددة
تطور الحصان
- مقالة مفصلة: تطور الحصان

يقدم الحصان أحد أفضل الأمثلة على التاريخ التطوري (علم الوراثة العرقي) بفضل السجل الأحفوري شبه الكامل والذي وجد في مستودعات رسوبية في أمريكا الشمالية تمتمد من أوائل عصر الإيوسين وحتى الحاضر.
تبدأ هذه السلسلة التطورية بحيوان صغير يسمى هايروكاثيريوم (أو الحصان الأولي، Eohippus)، الذي كان يعيش في أمريكا الشمالية منذ ما يقارب 54 مليون سنة، ثم انتشر إلى أوروبا وآسيا. تظهر البقايا الأحفورية للهايراكوثيريوم أنه يختلف مع الحصان الحديث في ثلاثة وجوه مهمة: كان حيوانًا صغيرًا (بحجم الثعلب)، بنيته خفيفة ومتكيف للركض؛ كانت أطرافه قصيرة ونحيلة، وأقدامه ممتدة حتى أن الأصابع كانت عمودية تقريبًا، وله أربعة أصابع في طرفيه الأماميين وثلاثة في طرفيه الخلفيين؛ وكانت قواطعه صغيرة، وأضراسه ذات تيجان منخفضة وشرفات مدورة تغطيها المينا.
المسار المرجح لتطور الحصان من هايراكوثيريوم إلى الحصان الحديث يشمل 12 جنسًا على الأقل وعدة مئات من الالأنواع. يمكن تلخيص الاتجاهات الرئيسية المشاهدة في تطور الحصان تبعًا للظروف البيئية المتغيرة كما يلي:
- زيادة في الحجم (من 0,4 م إلى 1,5 م)
- استطالة الأطراف والأقدام
- اختزال الأصابع الجانبية
- زيادة في طول الإصبع الثالث وسمكه
- زيادة في عرض القواطع
- استبدال الضواحك بالأضراس
- زيادة في طول الأسنان وارتفاع التيجان في الأضراس
تظهر النباتات المتحجرة في طبقات مختلفة أن الأرياف السبخية المشجرة التي كان يعيش فيها الهايراكوثيريوم أخذت في الجفاف تدريجيًا، وأصبح حينها البقاء يعتمد على كون الرأس في وضعية مرتفعة للتمتع بمنظر جيد للريف المحيط بالحيوان، وعلى سرعة جري عالية للفرار من المفترسات، ومن ثم الزيادة في الحجم واستبدال القدم المفرودة بالقدم ذات الحافر؛ فالأرض الجافة والصلبة تجعل القدم المنبسطة القديمة غير ضرورية للدعامة. يمكن تفسير التغيرات في الأسنان بافتراض أن النظام الغذائي قد تغير من كونه نباتات طرية إلى حشائش. اختير جنس سائد من كل فترة جيولوجية لإظهار التحور في سلالة الحصان من هيئته السلفية إلى هيئته الحديثة.
الانتقال من الأسماك إلى البرمائيات
قبل 2004 كان علماء الأحياء القديمة قد وجدوا مستحاثات لبرمائيات لها فقرات عنقية، وآذان، وأربع أرجل، في صخور لا يتجاوز عمرها 365 مليون سنة. ولم يستطيعوا أن يجدوا في الصخور الأقدم من 385 مليون سنة سوى الأسماك التي تفتقر إلى هذه الخصائص البرمائية. تنبأت نظرية التطور بأنه بما أن البرمائيات قد تطورت من الأسماك، فلابد من وجود هيئة انتقالية في الصخور التي تعود إلى الفترة الماضية ما بين 365 و385 مليون سنة. ويجب أن يمتاز هذا النوع الانتقالي بخصائص كثيرة تشبه الأسماك احتفظ بها من 385 مليون سنة أو أكثر، ولكن في الوقت ذاته يجب أن يمتاز بخصائص برمائية كذلك. في عام 2004 عثرت رحلة استكشافية انطلقت إلى جزر في القطب الشمالي الكندي بحثًا عن هذه الأحفورة تحديدًا، عثرت في صخور عمرها 375 سنة على أحافير لتيكتاليك.[61] لكن بعد ذلك ببضعة سنوات عثر علماء في بولندا على آثار كائن رباعي الأرجل متحجرة أقدم من تيكتاليك.[62]
أدلة من التوزيع الجغرافي
بيانات وجود الأنواع أو غيابها في القارات والجزر المختلفة (الجغرافيا الحيوية) يمكن أن تقدم أدلة على اشتراك السلف وأن تسلط الضوء على أنماط الانتواع.
التوزيع القارّي
كل المتعضيات متكيفة لبيئاتها بدرجات متفاوتة. إذا كانت العوامل الأحيائية وغير الأحيائية في موطن ما قادرة على دعم نوع معين في منطقة جغرافية ما، فقد يفترض المرء أن النوع ذاته سيوجد في بيئة مشابهة في منطقة جغرافية مشابهة، مثلًا: أفريقيا وأمريكا الجنوبية. الأمر ليس كذلك. توزيع أنواع النباتات والحيوانات متقطع حول العالم:
- في أفريقيا توجد قردة العالم القديم، والقردة، والفيلة، والنمور، والزرافات، وأبو قرن
- في أمريكا الجنوبية توجد قردة العالم الحديث، وأسود الجبال، واليغور، والكسلان، واللاما، والطوقان
- الصحارى في أمريكا الشمالية والجنوبية تحتوي على نباتات صبار متوطنة، أما الصحاري الأفريقية والآسيوية والأسترالية فتحتوي نباتات عصارية (ما عدا Rhipsalis baccifera)[63] وهي نباتات فربيونية تشبه الصبار شكلًا ولكنها مختلفة جدًا عنه.
يمكن ملاحظة اختلافات أكبر إن وضعنا أستراليا في الحسبان؛ فبالرغم من أنها تقع في دوائر العرض نفسها التي تقع عليها أجزاء كبيرة من أفريقيا وأمريكا الجنوبية، نجد أن الجرابيات كالكنغر والبندقوط والكوول تشكل قرابة نصف الأنواع الثديية أصلية الموطن في أستراليا.[64] يقابل هذا الغياب التام للجرابيات في أفريقيا وجزءًا صغيرًا من الحيوانات الثديية في أمريكا الجنوبية حيث يوجد الأبصوم والأبصوم الزبابي وقرد الجبل. الممثل الحي الوحيد للثدييات البدائية التي تضع البيض (أحاديات المسلك) هو خلد الماء وفصيلة النضناض. يعيش النضناض قصير الأنف بأنواعه في أستراليا، وتسمانيا، وغينيا الجديدة، وجزيرة الكنغر، أما النضناض طويل الأنف فيعيش فقط في غينيا الجديدة. يعيش خلد الماء في مياه شرق أستراليا، وأُدخل إلى تسمانيا، وجزيرة كنج، وجزيرة الكنغر. أحاديات المسلك هذه لا وجود لها في أي مكان آخر في العالم.[65] من ناحية أخرى تفتقد أستراليا إلى الكثير من مجموعات الثدييات المشيمية المنتشرة في القارات الأخرى (اللواحم، وشفعيات الأصابع، والزباب، والسنجابيات، وأرنبيات الشكل)، إلا أن بها خفافيش وفئران متوطنة. ووقد أدخل البشر أنواعًا أخرى كثيرة من المشيميات إلى أستراليا مثل الأرانب والثعالب.
من الأمثلة الأخرى توزيع الحيوانات الدب، حيث يوجد في جميع القارات باستثناء أفريقيا وأستراليا والقارة القطبية الجنوبية، والدب القطبي لا يوجد إلا في الدائرة القطبية الشمالية وما جاورها من يابسة.[66] البطريق يوجد حول القطب الجنوبي فقط بالرغم من تشابه ظروف الطقس في القطب الشمالي. عائلات الخيلانيات موزعة حول مياه الأرض بطريقة خاصة، حيث يتواجد خروف البحر في مياه غرب أفريقيا وشمال أمريكا الجنوبية والكاريبي، بينما قريبه الأطوم لا توجد إلا في مياه أوقيانوسيا شمال أستراليا وسواحل المحيط الهندي. كما أن بقرة بحر ستلر المنقرضة كانت تعيش في بحر بيرنغ.[67]
يمكن العثور على أحافير النوع الواحد في مناطق يُعرف أنها كانت متجاورة في الماضي ثم أصبحت الآن في مواقع جغرافية متباعدة بفعل الانجراف القاري. مثلًا: توجد أحافير للأنواع القديمة نفسها من البرمائيات، والمفصليات، والسراخس في أمريكا الجنوبية وأفريقيا والهند وأستراليا والقطب الجنوبي، وتعود للحقبة الأولية حين كانت هذه المناطق متحدة في كتلة يابسة واحدة تسمى غندوانا.[68] أحيانًا يمكن التعرف على السلالة الحية لهذه المتعضيات ويظهر التشابه بينها جليًا بالرغم من كونها تسكن الآن مناطق مختلفة جدًا جغرافيًا ومناخيًا.
الجغرافيا الحيوية للجزر

لعبت الأدلة المستمدة من علوم الجغرافيا الحيوية للجزر دورًا مهمًا وتاريخيًا في تطوير علم البيولوجيا التطورية. تنقسم الجزر بحسب الجغرافيا الحيوية إلى فئتين. الجزر القارية وهي جزر مثل بريطانيا العظمى واليابان التي كانت فيما مضى جزءًا من قارة. والجزر المحيطية مثل جزر هاواي وجزر غالاباغوس وسانت هيلينا وهي الجزر التي تشكلت في المحيط ولم تكن أبدًا جزءًا من أي قارة. تملك الجزر المحيطية توزيعًا غير متوازن للنباتات والحيوانات المحلية بطرق تجعل هذه الأنواع متميزة عن الكائنات الحية الموجودة في القارات أو الجزر القارية. لا تحتوي الجزر المحيطية على ثدييات أصلية (تحتوي في بعض الأحيان على الخفافيش والفقمات) أو برمائيات أو أسماك المياه العذبة. تملك الجزر المحيطية في بعض الحالات زواحف أرضية (مثل الإغوانة والسلاحف العملاقة في جزر غالاباغوس) ولكنها في اغلب الأحيان لا تحوي زواحف (مثل جزر هاواي). وذلك على الرغم من حقيقة أنه عندما يتم إدخال أنواع مثل الجرذان والماعز والخنازير والقطط والفئران وعلجوم القصب إلى مثل هذه الجزر من قبل البشر فهي تزدهر غالبا. أجرى العديد من العلماء بدءًا من تشارلز داروين تجارب وسجلوا ملاحظات أظهرت أن أنواع الحيوانات والنباتات التي لم يتم العثور عليها في هذه الجزر تتفق مع نظرية أنه تم استيطان هذه الجزر عن طريق الصدفة بواسطة النباتات والحيوانات التي كانت قادرة على الوصول إليها. يمكن أن يحدث هذا الاستيطان المفاجئ عن طريق الجو مثل البذور النباتية التي تحملها الطيور المهاجرة أو الخفافيش والحشرات التي يتم دفعها فوق البحر عن طريق الرياح أو عن طريق الطفو من قارة أو جزيرة أخرى فوق البحر (بعض أنواع بذور النباتات مثل جوز الهند التي يمكن أن تطفو في المياه المالحة)، والزواحف التي يمكن أن تعيش لفترات طويلة فوق نباتات طافية تحملها العواصف إلى البحر.[69]
التوطن
إن العديد من الأنواع الموجودة على الجزر النائية هي أنواع متوطنة في جزيرة معينة أو مجموعة من الجزر، وهذا يعني أنها لا توجد في أي مكان آخر على وجه الأرض. من الأمثلة على الأنواع المتوطنة في الجزر العديد من الطيور غير القادرة على الطيران في نيوزيلندا، والليمور في مدغشقر، وتنين كومودو في كومودو، [70] شجرة دم الأخوين في سقطرى،[71] والطراطرة في نيوزيلندا[72][73] وغيرها. ومع ذلك ترتبط العديد من هذه الأنواع المستوطنة بأنواع موجودة في جزر أو قارات أخرى قريبة، ومن الأمثلة المعروفة على ذلك علاقة الحيوانات الموجودة في جزر غالاباغوس بتلك الموجودة في أمريكا الجنوبية.[74] يمكن شرح كل هذه الدلائل (أنواع النباتات والحيوانات الموجودة في الجزر المحيطية، والعدد الكبير من الأنواع المستوطنة الموجودة في الجزر المحيطية وعلاقة هذه الأنواع بتلك التي تعيش في القارات القريبة) بسهولة من خلال فرضية استعمار الجزر من قبل بعض الأنواع الموجودة في القارات القريبة والتي تطورت فيما بعد إلى أنواع متوطنة موجودة الآن هناك.[74]
لا ينبغي أن تشمل انواع التوطن الأخرى (بالمعنى الحرفي للكلمة) الجزر. يمكن أن تدل الجزر على البحيرات المعزولة أو المناطق النائية. وتشمل بعض الأمثلة على ذلك المرتفعات في إثيوبيا وبحيرة بايكال وفنبوس في جنوب إفريقيا، وغابات كاليدونيا الجديدة وغيرها. بعض الأمثلة عن الكائنات الحية المستوطنة التي تعيش في مناطق معزولة: الكاغو في كاليدونيا الجديدة،[74] والجرذان في غابات الصنوبر الاستوائية في لوزون في الفلبين،[75] شجرة بوجوم في شبه جزيرة باخا في كاليفورنيا،[76][77] فقمة بايكال [78] وسمك أومول في بحيرة بايكال.
التشعبات التكيفية
غالبًا ما يسكن الجزر المحيطية مجموعات من الأنواع متصلة بشكل وثيق والتي تمثل مجموعة متنوعة من الأنماط الحياتية، غالبًا ما تكون هذه الأنماط الحياتية مملوءة بأنواع مختلفة تمامًا في القارات. وتسمى هذه المجموعات مثل عصافير غالاباغوس وطائر هاواي الباحث عن العسل وأفراد عائلة عباد الشمس في أرخبيل خوان فرنانديز وسوس الخشب في سانت هيلانا تشعبات تكيفية لأن أفضل تفسير لذلك هو أن نوع واحد يستعمر جزيرة (أو مجموعة من الجزر) ثم يتنوع لملء البيئات الحيوية المتاحة. يمكن أن تكون هذه التشعبات مذهلة: مثل كون 800 نوع من عائلة ذبابة الفاكهة (وهو ما يقارب نصف المجموع العالمي) مستوطنة في جزر هاواي. مثال توضيحي آخر من هاواي هو نبات السيوف الفضية وهي مجموعة من ثلاثين نوعًا من النباتات توجد فقط في تلك الجزر. يتراوح افرادها من (السيوف الفضية) التي تزهر بشكل خاص على المنحدرات البركانية العالية إلى الأشجار والشجيرات والكروم التي تزهر على ارتفاعات مختلفة من أعلى الجبل إلى مستوى سطح البحر وفي بيئات هاواي التي تختلف من الصحارى إلى الغابات المطيرة. أقرب الأنواع إليهم خارج هاواي (استنادا إلى الدراسات الجزيئية) هي من أعشاب القطران الموجودة على الساحل الغربي لأمريكا الشمالية. تملك هذه الاعشاب بذور لزجة يسهل نشرها بواسطة الطيور المهاجرة.[79] ويمكن بالإضافة إلى ذلك ان تتكاثر جميع الأنواع الموجودة في الجزيرة بشكل متبادل وغالبًا ما تكون الكائنات الهجينة خصبة،[80] وقد تم بالفعل تهجين بعض الانواع تجريبياً مثل نوعين من أنواع أعشاب القطران من الساحل الغربي أيضًا.[37] تحتوي الجزر القارية على الكائنات الحية قليلة الانعزال، ولكن تملك الكائنات التي انفصلت لفترة طويلة عن أي قارة أنواع مستوطنة وتشعبات تكيفية، مثل وجود 75 نوعًا من حيوان الليمور في مدغشقر، وأنواع طيور الموا الإحدى عشرة المنقرضة في نيوزيلندا.[74][81]
الأنواع الحلقية

إن الأنواع الحلقية عبارة عن سلسلة متصلة من مجموعات الأفراد يمكن لكل منها أن تتزاوج مع جيرانها، مع ما لا يقل عن جماعتين نهائيتين تختلفان بشكل كبير من ناحية التهجين على الرغم من إمكانية تبادل الجينات بين كل المجموعات. [150] تمثل الأنواع الحلقية نمطًا من الأنتواع وقد تم الاستشهاد بها كدليل على التطور. فهي توضح ما يحدث بمرور الوقت حين تتباعد المجموعات جينيا، خاصة لأنها تمثل ما يحدث عادة في المجموعات الحية مع مرور الزمن بين مجموعات الأسلاف المتوفين منذ فترة طويلة والأفراد الأحياء حيث انقرض الأفراد الوسطاء. يقول ريتشارد دوكينز إن الانواع الحلقية «تظهر لنا شيئًا في البعد المكاني فقط يجب أن يحدث دائمًا في البعد الزماني».[82]
أمثلة من التوزيع الجغرافي
توزع السرخس اللساني
يمكن في بعض الأحيان استخدام كل من الانجراف القاري والتطور للتنبؤ بما يمكن العثور عليه في السجل الأحفوري. السرخس اللساني هو نوع منقرض من نباتات سرخس البذور من العصر البرمي. يظهر السرخس اللساني في السجل الأحفوري في بداية العصر البرمي في قارة غندوانا القديمة.[83] يشرح الانجراف القاري الجغرافيا الحيوية الحالية للنبات. وجدت مستحاثات السرخس اللساني اليوم في طبقات العصر البرمي في جنوب شرق أمريكا وجنوب شرق إفريقيا وكامل جزيرة مدغشقر وشمال الهند وكامل أستراليا ونيوزلندا ووجدت منتشرة على الأطراف الجنوبية والشمالية للقارة القطبية الجنوبية. كانت هذه القارات متصلة خلال العصر البرمي وحملت اسم غندوانا (الشكل 4c) يتفق ذلك مع الشريط المغناطيسي وتوزيع المستحاثات الأخرى والشقوق الجليدية التي تشير إلى عدم اعتدال المناخ في القطب الجنوبي أثناء العصر البرمي.[84][85]
توزيع بعد الوحشيات
يقدم تاريخ بعد الوحشيات (القسم الذي يحتوي على الشقبانيات وأسلافها البدائيين المنقرضين) مثالًا على كيفية دمج النظرية التطورية وحركة القارات في صياغة تنبؤات تتعلق بطبقات المستحاثات والتوزع. توجد أقدم مستحاثات كائنات بعد الوحشيات في الصين.[86] انتشرت بعد الوحشيات غربًا إلى أمريكا الشمالية الحديثة (لا تزال مرتبطة بأوراسيا) ثم إلى أمريكا الجنوبية التي كانت مرتبطة بأمريكا الشمالية منذ حوالي 65 مليون عام. وصلت الجراثيم إلى أستراليا عبر القارة القطبية الجنوبية منذ 50 مليون عام بعد وقت قصير من انقسام أستراليا مما يشير إلى حدوث انفصال لنوع واحد فقط.[87] تشير النظرية التطورية إلى أن الجرابيات الأسترالية تنحدر من اسلافها في الأمريكتين. وتشير الدلائل الجيولوجية إلى أن أمريكا الجنوبية وأستراليا كانتا منذ ما بين 30 و 40 مليون عام لا تزالان جزءًا من قارة غندوانا العظمى في نصف الكرة الجنوبي وأنهما كانتا مرتبطان بأرض أصبحت الآن جزءًا من القارة القطبية الجنوبية. لذلك يمكن للعلماء عند الجمع بين النماذج أن يتنبؤوا بأن الجرابيات هاجرت مما يعرف الآن بأمريكا الجنوبية عبر القارة القطبية الجنوبية ومن ثم إلى أستراليا الحالية منذ ما بين 30 و 40 مليون عام. تم العثور على أول مستحاثة للجرابيات في جزيرة سيمور في شبه الجزيرة القطبية الجنوبية في عام 1982.[88] وتم العثور على مستحاثات أخرى في وقت لاحق تشمل افراد من فصيلة الأبسوم وقرود الجبل،[89] وكذلك الحافريات وأفراد النوع المنقرض الغريب غوندوانا تيريا، وربما ثدييات أمريكا الجنوبية.[90][91][92]]
هجرة وانعزال وتوزع الجمليات
يقدم تاريخ الجمليات مثالاً على كيفية استخدام الأدلة الأحفورية لإعادة تشكيل أحداث الهجرة والتطور اللاحق. يشير السجل الأحفوري إلى أن تطور الإبل بدأ في أمريكا الشمالية (انظر الشكل 4e) حيث هاجرت منها منذ 6 ملايين عام عبر مضيق بيرنغ إلى آسيا ثم إلى إفريقيا، وقبل 3.5 مليون سنة عبر مضيق بنما لتصل لأمريكا الجنوبية. وما إن تم عزلهم حتى تطوروا وفقًا لمصلحتهم الخاصة، مما أدى إلى ظهور الجمل ذو السنمين والجمل العربي في آسيا وإفريقيا واللامة في أمريكا الجنوبية. ثم انقرضت الإبل في أمريكا الشمالية في نهاية العصر الجليدي الأخير.[93]
أدلة من الاصطفاء
غالبًا ما تظهر أدلة التطور من الملاحظة المباشرة للاصطفاء الطبيعي في الحقل والمختبر. يعتبر هذا القسم فريد من نوعه لأنه يوفر سياقًا محددا يتعلق بعملية الاختيار. ذكرت جميع الأمثلة المقدمة لهذا الدليل على أن التطور قد حدث بالفعل، لكنها لم تقدم الآلية الأساسية الرئيسية: وهي الانتقاء الطبيعي. يقدم هذا القسم صراحةً دليلًا على حدوث الانتقاء الطبيعي، وقد تم تمثيله في الابحاث، ويمكن تمثيله في تجارب مخبرية.
وثق العلماء العديد من الأحداث التي كان فيها الاصطفاء الطبيعي قيد التنفيذ. الأمثلة الأكثر شهرة هي مقاومة المضادات الحيوية في المجال الطبي إلى جانب التجارب المخبرية المعروفة التي توثق حدوث التطور. يعتبر الانتقاء الطبيعي محاكاة للسلف المشترك على المدى الطويل ويمكن أن تؤدي ضغوط الاصطفاء إلى تنوع الحياة على الأرض كما هو موجود اليوم. جميع التكيفات (الموثقة وغير الموثقة) ناتجة عن الانتقاء الطبيعي بالإضافة لعدد قليل من العمليات الثانوية الأخرى. لقد ثبت أن «... الاصطفاء الطبيعي هو جزء اساسي في عملية الأنتواع »،[94] وهو المحرك الرئيسي له،[95] وبالتالي فإن الأمثلة التالية من الانتقاء الطبيعي والانتواع غالباً ما تعتمد على بعضها البعض أو تتوافق مع بعضها البعض. الأمثلة أدناه ليست سوى جزء صغير من التجارب والملاحظات الفعلية.
الاصطفاء الاصطناعي والتطور التجريبي

يوضح الاصطفاء الاصطناعي التنوع الذي يمكن أن يوجد بين الكائنات الحية التي تشترك في سلف حديث نسبياً. يتم في الاصطفاء الاصطناعي تهجين نوع واحد بشكل انتقائي في كل جيل مما يسمح بالتكاثر فقط للكائنات الحية التي تظهر الخصائص المرغوبة. تصبح هذه السمات متطورة بشكل جيد في الأجيال المتعاقبة. كان الاصطفاء الاصطناعي ناجحاً قبل أن يكتشف العلم الأساس الجيني بوقت طويل. تشمل الأمثلة على الانتقاء الاصطناعي تربية الكلاب والأغذية المعدلة وراثياً وتربية الزهور وزراعة الأطعمة مثل الملفوف البري[96] وغيرها.
يستخدم التطور التجريبي التجارب لاختبار فرضيات ونظريات التطور. قام وليام دالينجر في أحد الأمثلة الأولى بإجراء تجربة قبل عام 1880، حيث عرّض الميكروبات للحرارة بهدف فرض تغييرات تكيفية. استمرت تجربته نحو سبع سنوات وكانت نتائجه موضع إشادة، لكنه لم يكمل التجربة بعد تعطل الأدوات.[97]
يوجد مثال شامل للتطور التجريبي هو تجربة تطور الإشريكية القولونية طويلة الأمد للعالم ريتشارد لينسكي. لاحظ لينسكي أن بعض سلالات الاشريكيات القولونية طورت قدرة جديدة معقدة لاستقلاب حمض الليمون بعد عشرات الآلاف من الأجيال.[98][99] علق عالم الأحياء التطورية جيري كوين على ذلك بوصفه إبداع، وقال: «الشيء الذي يعجبني أكثر هو قول مؤيدو نظرية الخلق أنه يمكنك تطوير هذه الصفات المعقدة من خلال اجراء مجموعة من الأحداث العشوائية. وهذا ما لا يمكن أن يحدث».[98] وبالإضافة إلى التغيرات الاستقلابية فقد تم اكتشاف تجمعات بكتيرية مختلفة قد اختلفت فيما يتعلق بكل من التشكل (الحجم الكلي للخلية) والصلاحية (التي تم قياسها بمقارنتها مع الأسلاف).[100] لا تزال تجربة تطور الإشريكية القولونية طويلة الأمد التي بدأت في عام 1988 مستمرة، وقد أظهرت بعض التكيفات بما في ذلك تطور سلالة من الإشريكية القولونية كانت قادرة على النمو مع حمض الليمون في وسائط النمو، وهي سمة غائبة في جميع الأشكال الأخرى المعروفة للإشريكية القولونية بما في ذلك السلالة الأولية.
اللافقاريات
قدرة تحمل الرصاص لدى برغوث الماء
تنبأت دراسة أجريت على أنواع من برغوث الماء وتلوث الرصاص في القرن العشرين أن زيادة التلوث بالرصاص ستؤدي إلى اصطفاء قوي لسمات تحمل الرصاص. كان الباحثون قادرين على استخدام (علم الاحياء البيئي الإحيائي) لفقس بيض برغوث ماء حُفظت لعقود من الزمن عندما كانت البحيرات ملوثة بالرصاص بشكل كبير. وتمت مقارنة براغيث الماء الجديدة مع براغيث الماء الحالية مما أظهر: «فروقًا كبيرة في الصلاحية بين الأنماط القديمة والحديثة بسبب مواجهة ضغوط بيئية كبيرة واسعة النطاق».
لم تكن براغيث الماء المعاصرة قادرة على مقاومة أو تحمل مستويات عالية من الرصاص (وهذا بسبب الانخفاض الكبير في تلوث الرصاص في بحيرات القرن الحادي والعشرين). ومع ذلك فقد تمكنت براغيث الماء القديمة من تحمل نسب تلوث الرصاص المرتفع. واستنتج الباحثون أنه «تمكنا من خلال استخدام تقنيات (علم الأحياء البيئي الإحيائي) من إظهار تغيير واضح على مدى عقود».[101]
العث المفلفل
مثال على الاصطفاء الطبيعي هو التغير الظاهري والتكيف اللوني من الفاتح للداكن في العثة المفلفلة، بسبب التلوث الناتج عن الثورة الصناعية في إنجلترا.
الجراثيم
مقاومة المضادات الحيوية
إن تطور وانتشار الجراثيم المقاومة للمضادات الحيوية هو دليل على عملية تطور الأنواع. وبالتالي فإن ظهور المكورات العنقودية الذهبية المقاومة للفانكومايسين والخطر التي تشكله على المرضى هو نتيجة مباشرة للتطور من خلال الاصطفاء الطبيعي. يوضح ظهور سلالات شيغيلا المقاومة لفئة المضادات الحيوية الاصطناعية من السلفوناميدات أيضًا ظهور أجيال جديدة كعملية تطورية.[102] وبالمثل فإن ظهور مقاومة مركب (ثنائي كلورو ثنائي فينيل ثلاثي كلورو الإيثان) في أشكال مختلفة من حشرات بعوض الأنوفيل وظهور ميزات مقاومة مرض الرام المخاطي في مجموعات الأرانب في أستراليا هي دلائل على حدوث تطور بسبب ضغط الاصطفاء التطوري في الأنواع التي تتجدد فيها الأجيال بسرعة.
تطور جميع فئات الجراثيم مقاومتها: بما في ذلك الفطريات (مقاومة المضادات الفطرية)، والفيروسات (مقاومة مضادات الفيروسات)، والكائنات الأولية (مقاومة مضادات الأوالي)، والجراثيم (مقاومة المضادات الحيوية). وهذا أمر متوقع عند النظر إلى أن كل أشكال الحياة تملك رمزًا جينيًا وبالتالي فهي تخضع لعملية التطور ضمن آلياتها المختلفة.
البكتريا الآكلة للنايلون
مثال آخر للكائنات الحية التي تتكيف مع الظروف الناتجة عن ممارسات الإنسان هي البكتريا الآكلة للنايلون: هي سلالة من الجراثيم الصّيفريّة قادرة على هضم بعض المنتجات الثانوية لتصنيع النايلون 6. هناك إجماع علمي على أنّ القدرة على أكل النايلون قد تطورت على الأرجح كطفرة بخطوة واحدة واستقرت لأنها حسنت من صلاحية البكتيريا التي امتلكت هذه الطفرة. يعتبر ذلك مثالًا جيدًا للتطور من خلال التطفّر والاصطفاء الطبيعي الذي لوحظ عند حدوثه ومن غير الممكن له أن يتحقق حتى تم إنتاج النايلون من قبل البشر.[103][104][105][106]
النباتات والفطريات
تشعب ازهاد القرد
كلا النوعين الفرعيين ذات الازهار الحمراء والصفراء لأزهار القرد معزولان بسبب تفضيلات طيور الطنانة وعثث أبو الهول الملقحة لهذه الازهار. معظم أنواع سلالات ازهار القرد ذات لون أصفر. ومع ذلك يوجد نوعين من الازهار الحمراء. يقترح التحليل الوراثي وجود أصلين مستقلين من الزهور ذات اللون الأحمر التي نشأت بسبب الطفرات في جين (MaMyb2) الموجود في جميع أنواع أزهار القرد.
أشارت الأبحاث إلى أن عدم حدوث طفرتين مستقلتين، ولكن تم نقل أليل (MaMyb2) واحد عن طريق عملية التهجين.[107] تقدم هذه الدراسة مثالاً على تداخل البحوث في مختلف التخصصات. إنَّ انعزال الجينات والوظائف التنظيمية المتوافقة في الحمض النووي وتحليل شجرة تطور السلالات والموقع الجغرافي وتفضيل الكائنات المُلقحة وتهجين الأنواع والانتواع ليست سوى بعض المجالات التي يمكن الحصول على البيانات منها لتوثيق حدوث التطور.
=التغذية الاشعاعية للفطريات
يمكن للتلوث الناجم عن الإنسان أن يأتي بأشكال مختلفة. تعتبر التغذية الإشعاعية للفطريات مثالًا على الانتقاء الطبيعي الذي يحدث بعد حدوث تغير كيميائي. يبدو أن التغذية الاشعاعية للفطريات تستخدم اصبغة الميلانين لتحويل إشعاع جاما إلى طاقة كيميائية للنمو[108][109] واكتُشف لأول مرة في عام 2007 مع نمو العفن الاسود داخل وفي محيط محطة الطاقة النووية في تشيرنوبيل.[108] أظهرت الأبحاث في كلية ألبرت أينشتاين للطب أن ثلاثة فطريات تحتوي على الميلانين هي (الفطريات المبغثرة) و (فانجيلا الملهبة للجلد) و (مستخفية مورمة) قد زادت كتلتها الحيوية وراكمت الأسيتات (الخلات) بشكل أسرع في بيئة كان فيها مستوى الإشعاع أكبر بـ 500 ضعف من مستوى الإشعاع في البيئة الطبيعية.
الفقاريات
أسماك الجوبي

لاحظ عالم الأحياء جون إندلر أثناء دراسة أسماك الجوبي في جزيرة ترينيداد حدوث الاصطفاء ضمن مجموعات الأسماك. ومن اجل استبعاد الاحتمالات البديلة للاصطفاء أنشأ إندلر تجربة تحكّم بها بدقة لمحاكاة البيئات الطبيعية من خلال بناء عشرة أحواض مخبرية بجامعة برينستون. يحتوي كل حوض على حصى لتماثل تمامًا البيئات الطبيعية. بعد التقاط عينة عشوائية من أسماك الجوبي من الأحواض في ترينيداد، قام بتربيتها وخلطها لإنشاء تجمعات متنوعة وراثيا متماثلة فيما بينها، وقام بملاحظة كل سمكة (طول وارتفاع ومساحة البقعة، طول البقعة النسبي، ارتفاع البقعة النسبي، مساحة البقعة الكلية، وأطوال الجسد القياسية). أضاف العالم للتجربة اسماك (البلطي الرمحية) التي تعتبر المفترس الرئيسي لأسماك الجوبي في أربعة أحواض، وأضاف أسماك الريفولوس غير المفترسة في أربعة أحواض أخرى، وترك البركتين الباقيتين تحتويان فقط أسماك الجوبي. أجرى مقارنات بعد 10 أجيال بين أسماك الجوبي في كل بركة وأخذ القياسات مرة أخرى. وجد إندلر أن الأسماك قد طورت أنماط ألوان مختلفة اختلافًا جذريًا في الاحواض التي حوت أسماك الجوبي فقط أو الأسماك غير المفترسة وأنماط ألوان باهتة في أحواض الأسماك المفترسة. تسبب ضغط الافتراس في حدوث الاصطفاء لتتناسب مع الحصى في بيئتها.[110]
أجرى إندلر في موازاة التجربة السابقة تجربة ميدانية في ترينيداد حيث اخذ أسماك الجوبي من الأحواض التي تحوي اسماك مفترسة ونقلهم إلى أحواض أخرى أعلى حيث لا يوجد اسماك مفترسة. وجد اندلر بعد 15 جيلًا أن أسماك الجوبي المنقولة قد طورت أنماطًا مثيرة وملونة. أظهرت كلتا التجربتين تقاربا جوهريا بسبب ضغوط الاصطفاء المتماثلة (أي اختيار الأسماك التي تعرضت للافتراس لأنماط الألوان تتناسب مع بيئتها والانتقاء الجنسي لأنماط الألوان الاخرى في حال عدم تعرضها للافتراس).[110]
أجرى ديفيد ريزنيك دراسة لاحقة تم خلالها فحص الاسماك في بيئتها الاصلية بعد 11 عامًا منذ نقل إيندلر أسماك الجوبي إلى احواض عالية لا تحوي مفترسين. وجدت الدراسة أن الأسماك قد تطورت بعدد من الطرق المختلفة مثل: أنماط الألوان الزاهية، والنضج المتأخر، والأحجام الكبيرة، وصغر حجم الأسماك حديثة الولادة، وعدد نسل أكبر.[111] أشارت دراسات أخرى على أسماك الجوبي والحيوانات المفترسة في مياه ترينيداد إلى أن أنماط الاصطفاء المختلفة من خلال الافتراس لم تغير أنماط وأحجام وسلوكيات أسماك الجوبي فحسب ولكنها غيرت تواريخ حياتها وأنماط هذه الحياة.[112]
البشر
لوحظ الاصطفاء الطبيعي في المجموعات البشرية المعاصرة، حيث أظهرت النتائج الحديثة أن السكان المعرضين لخطر الإصابة بمرض كورو الخطير يملكون تمثيل مفرط لمتغير مناعي لجين البروتين (G127V) على حساب الأليلات غير المناعية. يفترض العلماء أن أحد أسباب الاختيار السريع لهذا المتغير الجيني هو خطر الموت من جراء المرض لدى الأشخاص الذين لا يتمتعون بالمناعة.[113][114] تشمل الاتجاهات التطورية الأخرى المكتشفة في السكان إطالة فترة التكاثر، وانخفاض مستويات الكوليسترول ونسبة الجلوكوز في الدم وضغط الدم.[115]
ومن الأمثلة المعروفة عن الاصطفاء في المجموعات البشرية هو عدم تحمل اللاكتوز. عدم تحمل اللاكتوز هو عدم القدرة على استقلاب اللاكتوز بسبب نقص انزيم اللاكتاز وهو الإنزيم المطلوب لهذه العملية في الجهاز الهضمي. الحالة الطبيعية للثدييات هي أن يعاني الصغار من انخفاض إنتاج اللاكتاز في نهاية فترة الفطام (وهي فترة زمنية محددة لكل نوع). بالنسبة البشر الذين يعيشون ضمن مجتمعات لا تستهلك الألبان ينخفض عادة إنتاج اللاكتاز نحو 90٪ خلال السنوات الأربع الأولى من الحياة، وتختلف قيمة الانخفاض الدقيقة مع مرور الوقت على نطاق واسع.[116] يرتبط استمرار نشاط افراز اللاكتاز لدى البالغين بتعدد الأشكال: (C / T 13910) و (G / A 22018) الموجودة في جين (MCM6).[117] يزيل هذا الاختلاف الجيني التوقف في إنتاج اللاكتاز مما يتيح لأفراد هذه المجموعات الاستمرار في استهلاك الحليب الخام ومنتجات الألبان الطازجة والمُخمرة طوال حياتهم دون مواجهة صعوبة هضمها. يبدو ذلك تكيف حديث تطوريًا لاستهلاك الألبان (منذ نحو 10000 عام، وقبل 7500 عام في أوروبا) [118] وحدث بشكل مستقل في كل من شمال أوروبا وشرق إفريقيا في السكان الذين لديهم نمط غذائي يعتمد على الألبان تاريخيًا.[119][120]
سحالي الجدار الإيطالية
تم نقل عشرة عينات بالغة من سحالي الجدار الايطالية في عام 1971 من جزيرة بود كوبيست الكرواتية إلى جزيرة بود ميركارو (تبعد عنها نحو 3.5 كم إلى الشرق). تقع كلتا الجزيرتنن في البحر الأدرياتيكي بالقرب من لاستوفو حيث أنشأت السحالي عددًا جديدًا من الافراد بعد ظاهرة عنق الزجاجة.[121][122] تتمتع الجزيرتان بنفس الحجم والارتفاع والمناخ المصغر وغياب عام للحيوانات المفترسة على الأرض[122] وعاشت السحالي لعقود دون تدخل بشري، حتى دون تدخل سحالي الجدار الدلماسية (أصبحوا حاليا منقرضين عن تلك الجزيرة). [123]
عاد العلماء إلى بود ميركارو في التسعينيات ووجدوا أن السحالي هناك تختلف اختلافًا كبيرًا عن تلك الموجودة في بود كوبيست. بينما أثبتت تحليلات الحمض النووي للميتوكوندريا أن سحالي الجدران الإيطالية الموجودة حاليًا على ميركارو تتشابه بشكل كبير من الناحية الوراثية مع الافراد الاصليين من بود كوبيست،[121] كان لدى مجموعة سحالي بود ميركارو الجديدة متوسط حجم أكبر وأطراف خلفية أقصر وكانت سرعتها القصوى أقل واختلفت استجابتها للهجمات المفترسة مقارنة بالسحالي الأصلية في بود كوبيست.[122] تُعزى هذه التغييرات إلى قلة أحداث الافتراس وزيادة الحماية من الغطاء النباتي في بود ميركارو.[122]
كشفت المزيد من الدراسات التحليلية في عام 2008 أن سحالي جزيرة ميركارو اختلفوا بشكل كبير في شكل الرأس (أطول وأوسع) وقوة عض أكبر مقارنةً بسحالي كوبيست الأصلية.[121] يتوافق هذا التغيير في شكل الرأس مع تغير في النظام الغذائي: تتغذى سحالي جزيرة كوبيست بشكل رئيسي على الحشرات، ولكن تتغذى السحالي الموجودة على ميركارو بشكل كبير على النباتات.[121] قد تكون التغييرات في أسلوب البحث عن الطعام قد ساهمت في زيادة عدد الأفراد وانخفاض سلوك السيطرة على الاراضي لدى سحالي جزيرة ميركارو.[121]
عثر على اختلاف آخر بين مجموعتي السحالي: هو اكتشاف وجود صمام لفائفي أعوري في سحالي ميركارو والتي تبطئ عملية مرور الغذاء وتوفر غرف تخمير، مما يسمح للكائنات الحية المجهرية المتعايشة معها بتحويل السليلوز إلى مغذيات قابلة للهضم بواسطة السحالي.[121] واكتشف الباحثون بالإضافة إلى ذلك أن الديدان الاسطوانة كانت شائعة في أحشاء سحالي ميركارو، لكنها غير موجودة في سحالي كوبيست والتي لا تملك على صمام لفائفي أعوري.[121] وصف الصمام اللفائفي الأعوري الموجود في أقل من 1% من جميع الأنواع المعروفة من الزواحف[121] بأنه «حدث تكيفي، ميزة جديدة غير موجودة في الأجداد وتطورت حديثًا في هذه السحالي».[124]
مقاومة مركبات الهيدروكربون العطري متعدد الحلقات لدى أسماك القوعاء
أجريت دراسة مماثلة فيما يتعلق بالهيدروكربونات العطرية متعددة الحلقات (PAHs) التي تلوث مياه نهر إليزابيث في بورتسموث في فرجينيا. تنتج هذه المادة الكيميائية عن الكريوزوت (نوع من القطران). طورت اسماك القوعاء الأطلسية مقاومة لمركبات الهيدروكربونات العطرية متعددة الحلقات من خلال جين AHR (نفس الجين الموجود في أسماك تومكودس وهي نوع من أسماك القد). وركزت هذه الدراسة على مقاومة السمية الحادة وتشويه القلب الناجم عن تلك المادة.
مقاومة ثنائي الفينيل متعدد الكلور لدى سمك القد
من الأمثلة على الملاحظة المباشرة لتغير الجينات بسبب ضغوط الانتقاء مقاومة مركبات ثنائي الفينيل متعدد الكلور لدى سمك القد. بعد أن طرحت شركة جنرال إلكتريك مركب ثنائي الفينيل متعدد الكلور (PCBs) في نهر هدسون بين عامي 1947 و 1976، استنتجوا ان سمك القد الذين يعيش في النهر قد طور مقاومة متزايدة لآثار المركب السامة.[125] سبب التأقلم مع السموم هو تغيير في ترميز جين معين. تم أخذ عينات جينية من أسماك القد من ثمانية أنهار مختلفة في منطقة نيو إنجلاند هي: نهر سانت لورانس ونهر ميراميتشي ونهر مارغري ونهر سكوامسكوت ونهر نيانتيك ونهر شينيكوك بيسك ونهر هدسونك ونهر هاكينساك. وجد التحليل الوراثي أن جين AHR2 الموجود في مجموعات أسماك القد في الأنهار الأربعة الواقعة في أقصى الجنوب كان يوجد كأليل مختلف حيث حذف منه حمضين امينيين.[126] منح هذا الحذف ميزة مقاومة مركب ثنائي الفينيل متعدد الكلور في أنواع الأسماك، ووجد في 99٪ من سمك التومكودس من نهر هدسون، 92٪ في نهر هاكنساك، 6٪ في نهر نيانتيك، و 5٪ في خليج شينيكوك.[126] يمثل هذا التغير في الأسماك المأخوذة من عينات المياه ارتباطًا مباشرًا بالضغوط الانتقائية التي أدت إلى تطور مقاومة ثنائي الفينيل متعدد الكلور في أسماك التومكود (القد) الأطلسية.[126]
الكائنات البرية في المدن

يعتبر وجود أشكال الحياة البرية في المدن حالة واسعة يمكن ملاحظتها بسهولة بسبب ضغط الانتقاء الناتج عن الإنسان ضد الحياة البرية. تكيفت حيوانات مختلفة بسبب نمو المناطق البشرية من أجل البقاء داخل هذه البيئات الحضرية. يمكن لهذه الأنواع من البيئات ممارسة ضغوط الانتقاء على الكائنات الحية، وغالبًا ما يؤدي ذلك إلى تكيفات جديدة. على سبيل المثال يحتوي نبات السراغة الموجود في فرنسا نوعين من البذور: ثقيلة وخفيفة. تسقط البذور الثقيلة بالقرب من النبات الأم في حين تطير البذور الخفيفة مع الريح. غالبًا ما تقع البذور الخفيفة التي تطير مع الريح في البيئات الحضرية على الخرسانة. لذلك تطورت هذه النباتات خلال نحو 5-12 جيلا لإنتاج بذور أثقل بكثير من مثيلاتها في المناطق الريفية.[127][128] ومن الأمثلة الأخرى على تكيف الحياة البرية في المناطق الحضرية الحمام الجبلي وأنواع الغربان التي تتكيف مع بيئات المدن في جميع أنحاء العالم، وطيور البطريق الأفريقية في بلدة سيمون، حيوانات الرباح في جنوب أفريقيا. ومجموعة متنوعة من الحشرات التي تعيش في المناطق البشرية. وجدت بعض الدراسات تغييرات مذهلة في سلوك الحيوانات (وتحديداً الثدييات) وحجم الدماغ الجسدي بسبب تفاعلها مع البيئات التي بناها الإنسان.[129][130]
سحالي الرمال البيضاء
تساعد الحيوانات التي تظهر تغيرات في البيئية الاجتماعية بإجراء البحوث المتعلقة بالآليات التي تحافظ من خلالها على تميز الأفراد. تم الحصول على ثروة من المعلومات حول الاصطفاء الطبيعي والتنوع الوراثي والظواهر الوراثية[131][132] والتكيف والتشكل البيئي[133] والإشارات الاجتماعية [134] من الدراسات التي أجريت على ثلاثة أنواع من السحالي الموجودة في صحراء الرمال البيضاء في نيو مكسيكو. أظهرت كل من سحالي هولبروكيا المرقطة والسحالي ذات الذيل الخيطي الطويل وسحالي السور الشرقية تجمعات بيئية تتشابه مع التربة الداكنة أو الرمال البيضاء في المنطقة. وقد وجدت الأبحاث التي أجريت على هذه الأنواع اختلافات شكلية ووراثية كبيرة بين السحالي الداكنة والسحالي الفاتحة بسبب ضغوط الانتقاء القوية. على سبيل المثال اظهرت سحالي هولبروكيا المرقطة اختلاف شكلي كبير (يتطابق بشكل أفضل مع لون بيئتها) مقارنة بالسكان ذوي اللون الفاتح الذي يبدون أقل نسبة تبادل للجينات بين المجموعات وأعلى اختلافات جينية عند مقارنتها بالأنواع الأخرى من السحالي.[131]
تعتبر الرمال البيضاء في نيو مكسيكو تكوين جيولوجي حديث (منذ نحو 6000 سنةref name=JMRobertson /> وربما 2000 سنة فقط[131]). يشير هذا التغير الحديث لكثبان الرمال الجبسية إلى أن الأنواع التي تظهر اختلافات ذات لون أفتح قد تطورت في إطار زمني قصير نسبيًا. تم ذكر الأنواع الثلاثة للسحالي السابقة لعرض تغير اللون من أجل التعايش مع المتغيرات البيئية.[134] لم تطور هذه الأنواع الثلاثة ألوانها فقط بسبب ضغوط الانتقاء البيئية بل طورت أيضًا سمات التشكل البيئي مثل: التشكل والسلوك (سلوك الهروب) والأداء (سرعة الركض) بشكل جماعي.[134] وجد عمل روش نتائج مذهلة في سلوك الهروب لدى سحالي هولبروكيا المرقطة وسحالي السور الشرقية. عندما وضعوا السحالي الداكنة على الرمال البيضاء تضاءلت استجابتها للمفاجئات بشكل كبير. قد يكون ذلك النتيجة عوامل مختلفة تتعلق بدرجة حرارة الرمل أو حدة البصر، وبغض النظر عن السبب «... إن فشل السحالي في الهرب ضمن البيئات غير المتطابقة يمكن أن يجعلها غير قادرة على التكيف عندما مواجهة المفترسات».[134]
أدلة من الانتواع
الانتواع هو العملية التطورية التي تنشأ من خلالها أنواع بيولوجية جديدة. يقوم علماء الأحياء بالبحث في الأنواع باستخدام أساليب نظرية مختلفة لاستخلاص ما يشكل نوعًا ما، وهناك نقاش قائم يتعلق بالتصنيفات.[135] ومع ذلك فإن الكثير من الأبحاث الحالية تشير إلى أن «... الانتواع هو عملية اختلاف السلالات الناشئة، وليس حدثا يؤثر على جميع الجينات في وقت واحد» [136]، وفي الانتواع متباين الموطن (الشكل الأكثر شيوعًا من الانتواع) تكون «العزلة الإنجابية نتيجة ثانوية للتغير التطوري في الأفراد المعزولين، وبالتالي يمكن اعتباره حادثة تطورية».[137] يحدث الانتواع نتيجة لهذا التباين في الموطن، ومع ذلك فقد تم توثيق مجموعة متنوعة من العوامل المختلفة وغالبًا ما يتم تعريفها وتصنيفها بأشكال مختلفة (مثل: الانتواع المحازي أو الانتواع التماثلي أو التهجين). وقد لوحظ حدوث حالات من الأنتواع في كل من الطبيعة الأم والمختبر. لاحظ كل من فلورين وأودين أن «الأدلة المخبرية الجازمة على حدوث الانتواع متباين الموطن غير موجودة ..»، ومع ذلك وعلى عكس الدراسات المخبرية (التي تركز بشكل خاص على نماذج الانتواع متباين الموطن) «يحدث الانتواع بشكل مؤكد، ويجعل الكم الهائل من الأدلة الطبيعية رفض ذلك أمرا غير معقول».[138] جمع كوين وَ أور قائمة من 19 تجربة مخبرية على ذبابة الفاكهة تقدم أمثلة على الانتواع متباين الموطن عن طريق الانتقاء التباعدي واستنتجوا أنه «يمكن ان تتطور العزلة الإنجابية في الانتواع متباين الموطن كنتيجة ثانوية للانتقاء التباعدي».[139]
إنَّ الابحاث التي توثق الأنتواع وفيرة. قام علماء الأحياء بتوثيق العديد من الأمثلة على الأنتواع في الطبيعة، حيث أنتج التطور أنواعًا أكثر بكثير مما يراه أي عالم ضروريًا. على سبيل المثال: هناك أكثر من 350000 نوع من الخنافس.[140] وتأتي أمثلة عن الأنتواع من ملاحظات علم الجغرافيا الحيوية للجزر وعملية التشعب التكيفي. كما يمكن العثور على أدلة للأصل المشترك من خلال دراسات المستحاثات القديمة داخل الطبقات الجيولوجية. تمثل الأمثلة الموضحة أدناه أنماطًا مختلفة من الأنتواع وتوفر دليلًا قويًا على الأصل المشترك. من المهم أن التنويه إلى أن أبحاث الأنتواع لا تراقب بشكل مباشر الاختلاف من البداية إلى النهاية. وتؤدي الأبحاث احيانا إلى إعادة بناء التاريخ. إن الأمثلة على ذلك كثيرة، والأمثلة التالية ليست شاملة فهي لا تضم سوى جزء صغير من الحالات التي تمت ملاحظتها. تذكر دائما حقيقة أن «... الاصطفاء الطبيعي هو جزء موجود في كل أنماط الأنتواع ...»،[94] وهو المحرك الرئيسي لهذه العملية،[95] لذلك غالباً ما تحتوي أمثلة الأنتواع على الاصطفاء.
المستحاثات
لا يمكن للسجل الأحفوري تقديم اي معلومات عمّا كان يشكل نوعًا ما. يعتمد علماء الحفريات إلى حد كبير على عملية مختلفة: مفهوم علم تشكل الأنواع.[141] يميز علماء الحفريات الأنواع باختلاف شكلها بسبب عدم وجود معلومات مثل السلوك التكاثري أو المادة الوراثية في الحفريات.[141] أدى البحث المكثف في السجل الأحفوري إلى طرح نظريات عديدة تتعلق بالانتواع (في سياق علم الحفريات)، مع وجود العديد من الدراسات التي تشير إلى شيوع أحداث التوازن والتوازن النقطي وتفرع النسب. نشر إروين وعلماء أخرون في عام 1995 عملًا مهما تحت عنوان (اساليب جديدة للانتواع في السجل الأحفوري) والذي جمع 58 دراسة حول الانتواع الأحفوري (بين عامي 1972 و 1995). لإيجاد معظم الأمثلة التي تشير إلى التوازن (بما في ذلك التخلق التجددي أو التوازن النقطي) و 16 دراسة تشير إلى احداث الانتواع.[141]على الرغم من أن التوازن يبدو التفسير السائد للوهلة الأولى، إلا أن هذه الدراسة استنتجت أنه «... لا يبدو أي نمط مهيمن ...» مع «... توضح الدراسات وجود كلا من التوازن والتدرج في تاريخ سلالة واحدة». تستخدم العديد من الدراسات رواسب قاع البحر التي يمكن أن توفر كمية كبيرة من البيانات المتعلقة بمستحاثات العوالق الدقيقة.[141] يمكن استخدام تعاقب الحفريات في الطبقات لتحديد الاتجاهات التطورية بين الكائنات الأحفورية. ويمكن بالإضافة إلى ذلك تفسير حالات حدوث انتواع من البيانات وقد أجريت العديد من الدراسات لتوثيق كل من التطور الشكلي والانتواع.
الغلوبوروتاليا

قدمت أبحاث واسعة عن عوالق الغلوبوروتاليا ترونكاتولينوديس رؤية هامة لدراسات الجيولوجيا الحيوية والبيئية القديمة إلى جانب العلاقة بين البيئة والتطور. وجد الباحثون من دراسة في الجغرافيا الحيوية القديمة لعوالق الغلوبوروتاليا ترونكاتولينوديس أدلة تشير إلى تكوين نوع جديد (عن طريق عملية الانتواع متماثل الموطن). تشير الطبقات الرسوبية المدروسة والتي تحتوي على الأنواع الثلاثة من العوالق أنه قبل 2.7 مليون عام كان يوجد فقط نوعي غلوبوروتاليا كراسافورميس وغلوبوروتاليا توزاينسيس. حدث انتواع في ذلك الوقت ووجدت بعض الأشكال الوسيطة لفترة من الزمن. اختفى نوع غلوبوروتاليا توزاينسيس من السجل فيما بعد (مما يشير إلى انقراضها) ولكنه موجود كوسيط بين نوعي غلوبوروتاليا كراسافورميس الموجود وغلوبوروتاليا ترونكاتولينوديس. يطابق هذا السجل الاحفوري المخطط الموجود والذي بني على أساس السمات الشكلية للأنواع الثلاثة.[142] انظر الشكل 6a.
الشعوعيات
في دراسة واسعة أجريت على خمسة أنواع من الشعوعيات وهي (كالوسيسليتا كابا، بتيروكانيوم بريسماتيوم، بسيدوكولوس فيما، يوكرتيديوم كالفيرتينس، يوكرتيديوم ماتوياماي)، وثق الباحثون تغيرا تطوريا كبيرا في كل سلالة. وبالإضافة إلى ذلك أظهرت توجهات الأنواع (يوكرتيديوم كالفيرتينس ويوكرتيديوم ماتوياماي) اللتان تملكان صلة وثيقة أنه منذ نحو 1.9 مليون عام غزت يوكرتيديوم كالفيرتينس منطقة جديدة من المحيط الهادئ وأصبحت هذه المجموعة معزولة عن افراد النوع الرئيسي. توضح طبقات هذا النوع بوضوح أن هذه المجموعة المعزولة تطورت لنوع يوكرتيديوم ماتوياماي. ثم عادت لتغزو المنطقة التي لا يزال تحوي نوع يوكرتيديوم كالفيرتينس حيث حدث انخفاض مفاجئ في حجم الجسم. اختفى في النهاية نوع يوكرتيديوم ماتوياماي من الطبقة (من المفترض أن يكون ذلك نتيجةً للانقراض) تزامنًا مع مقاومة نوع يوكرتيديوم كالفيرتينس لانخفاض حجم الجسد. ومن ذلك الوقت توقف تغير حجم الجسد. يقترح المؤلفون زوال هذه السمات بسبب المنافسة.[143][144]
ريزوزلينيا (الدياتوم)
أجرى الباحثون قياسات على 5000 عينة من عوالق الريزوزلينيا (الدياتوم) من ثمانية مستحاثات رسوبية في المحيط الهادئ. تمتد العينات الأساسية على زمن مليوني عام وتم ترتيبها باستخدام قياسات المجال المغناطيسي للطبقات الرسوبية. اظهرت جميع العينات الأساسية نفس نمط التشعب: مع وجود سلالة واحدة (ريزوزلينيا بيرجوني) حدثت قبل 3.1 مليون عام وظهرت سلالتين وليدتين مختلفتين شكليا (ريزوزلينيا برايبيرجوني) بعد ذلك. كانت الوحدات المستخدمة لقياس العينات متماثلة في كل عينة.[145] وجدت دراسة إضافية جرت على أنواع ريزوزلينيا بيرجوني أنها وبعد حدوث الاختلاف قد غزت المحيط الهندي.[141][146]
التوربوروتاليا
أجريت دراسة حديثة على عوالق المنخربات حيث استخرج العلماء «51 عينة مرتبة حسب المستوى الطبقي من موقع استوائي داخل شمال المحيط الهادئ». تم فحص مئتي نوع من الأنواع الفردية استنادا لعشر سمات شكلية محددة (الحجم، مؤشر الانضغاط، نسبة أبعاد الحجرة، تضخم الحجرة، نسبة أبعاد الفجوة، ارتفاع الاختبار، توسع الاختبار، زاوية الصرّة، جهة الالتفاف، وعدد الحجرات في التجمع). وجدت الدراسة باستخدام طرق التجميع الإحصائي متعدد المتغيرات أن الأنواع استمرت في التطور بشكل غير مباشر في العصر الإيوسيني منذ ما بين 45 و 36 مليون عام. وأظهرت الطبقات الأحفورية منذ ما بين 36 و 34 مليون عام مجموعتين مختلفتين لهما خصائص مختلفة تميز بين نوعين من الكائنات. استنتج الباحثون أنه لا بد من حدوث انتواع وأن النوعين الجديدين كانا من سلالة النوع السابق.[147] يمثل هذا المثال الطبيعة الواسعة ومجموعة البيانات اللازمة من مختلف المجالات العلمية (مثل علم المحيطات وعلم الآثار) واستخدام التحليل الرياضي (مثل علم الإحصاء الحيوي).
الفقاريات
توجد أدلة على وجود انتواع لدى الفقاريات بالرغم من قلة البيانات التي يوفرها السجل الأحفوري. أجريت دراسات لتوثيق الأنماط المتماثلة في اللافقاريات البحرية.[141] وساهمت الأبحاث المكثفة التي وثقت معدلات التغير الشكلي، والاتجاهات التطورية، وأنماط الانتواع في الثدييات الصغيرة[148] بشكل كبير في الانجازات العلمية، وفي النهاية: إن إثبات أن التطور (والانتواع) قد حدث فعلا في الماضي يدعم فرضية السلف المشترك.
وجدت دراسة أجريت على أربعة أجناس من الثدييات: الهايبسودوس، والبيليكودوس، والهابلومايلوس (الثلاثة من العصر الإيوسيني) وشبيهات الأدابيس (من العصر الباليوسيني) من خلال دراسة عدد كبير من الطبقات وأخذ العينات أن كل مجموعة قد أظهرت «تطور شكلي تدريجي، وزيادة الحجم العام وتطور الأنواع الصغيرة بشكل تكراري، واختلاف السمات في كل سلالة جديدة».[149] استنتج العلماء من خلال هذه الدراسة أن الانتواع كان واضحًا. وجدت دراسة أخرى عن توجهات التغير الشكلي ومعدلات التطور أن القوارض الأرفيكولاواتية تتشعب لـ 52 سلالة مختلفة على مدى فترة زمنية تبلغ 5 ملايين سنة، وقد وثقت أمثلة عن التطور الشكلي المتدرج والتوازن.[150]
اللافقاريات
ذبابة الفاكهة سوداء البطن

وجد ويليام رايس وجورج سالت دليلًا تجريبيًا على حدوث انتواع متماثل الموطن في ذبابة الفاكهة الشائعة. قاموا بجمع مجموعة من ذبابة الفاكهة من مدينة دافيس في كاليفورنيا ووضعوا الشرانق في متاهة بيئية. كان على الذباب الجديد البحث في المتاهة للعثور على الطعام. وكان للذباب ثلاثة خيارات من أجل العثور على الطعام، وهي اتباع الضوء والظلام (طريق ضوئي) أو الحركة للأعلى والأسفل (طريق جغرافي) أو اتباع رائحة الأسيتالديهيد ورائحة الإيثانول (طريق كيميائي). قسم ذلك في النهاية الذباب إلى 42 بيئة مكانية وزمانية.
ثم قاموا بأخذ سلالتين اختارتا بيئات معاكسة. تأقلمت واحدة من السلالات خلال وقت قصير وحلّقت على الفور إلى الأعلى في الظلام لتنجذب إلى الأسيتالديهيد. تأقلمت سلالة أخرى في وقت متأخر وحلقت على الفور إلى أسفل منجذبة إلى الضوء والإيثانول. ثم تم وضع شرانق السلالتين معًا في المتاهة وسمح لها بالتزاوج في مكان وجود الطعام. ثم جمعوها. وفرضت عقوبة انتقائية على الذباب الإناث التي غيرت بيئتها. يحتم ذلك عدم انتقال أعراسها إلى الجيل التالي. ظهر انعزال تكاثري بين السلالتين بعد مضي 25 جيلًا من اختبار التزاوج. كرروا التجربة مرة أخرى دون تطبيق عقوبة انتقائية على تغيير البيئة وكانت النتيجة نفسها وهي إنتاج العزلة التناسلية.[151][152][153]
الدبابير الآبرات
وجدت دراسة عن أنواع دبور الآبرات أن الافراد التي تتغذى على النباتات المضيفة المختلفة (سنديان مزدوج وسنديان فرجيني) أظهرت حجم جسم وشكل عفصة مختلفًا إلى جانب تعبير قوي عن العزلة الجنسية. افترضت الدراسة أن الافراد الذين يعيشون ضمن نباتات مضيفة مختلفة سيظهرون دليلًا على التطور التباعدي الذي يدعم حدوث الأنتواع. قام الباحثون بأخذ عينات من أنواع الدبابير الآبرة في أشجار البلوط وقاموا بقياس حجم الجسم (قصبة الساق اليمنى لكل دبور)، واحصاء عدد حجرات العفص. بالإضافة إلى ذلك أجروا فحوصات التزاوج والتحليلات الإحصائية. وتم إجراء التحليل الوراثي أيضًا على موقعين من الدنا المتقدرة (416 زوجًا قاعديا من السيتوكروم C و 593 زوجًا قاعديا من السيتوكروم أوكسيديز) من أجل «التحكم في التأثيرات الخارجية غير المتوقعة للوقت عند الاختلاف بين الأنواع التي خضعت للانتواع متباين الموطن».[154]
قام الباحثون بدراسة نوعين من الدبابير الآبرة هي (بيلونوسنيما تريتي) و(ديشولكاسبيس كيركوسفيرنس) ووجدوا تباينًا شكليا وسلوكيًا كبيرا بين المجموعات المرتبطة بنبات مضيف. حققت هذه الدراسة الشروط المسبقة للانتواع.[155]
ذبابة الزعرور
أحد الأمثلة على حدوث التطور هي ذبابة الزعرور (والمعروفة أيضًا باسم ذبابة التفاح) والتي يبدو أنها خضعت للانتواع متماثل الموطن.[156] تتغذى مجموعات مختلفة من ذبابة الزعرور على ثمار مختلفة. ظهرت مجموعة مختلفة في أمريكا الشمالية في القرن التاسع عشر بعد ظهور التفاح وهي ليست وليدة البيئة ذاتها. تتغذى مجموعات ذبابة الزعرور الجديدة عادةً على التفاح فقط وليس على ثمار الزعرور التي يفضلها هذا النوع بشكل عام، لا تتغذى مجموعة ذبابة الزعرور الأصلية عادة على التفاح. توجد بعض الأدلة (مثل حقيقة أن ستة من أصل ثلاثة عشر من الإنزيمات المتغايرة جينيا مختلفة) تقول أنّ ذباب الزعرور ينضج في وقت متأخر من الموسم ويستغرق وقتًا أطول لينضج مقارنة بذباب التفاح، وتوجد القليل من الأدلة على حدوث التهجين (قام الباحثون بتوثيق حدوث التهجين بنسبة بلغت 4 - 6٪) والتي تشير إلى حدوث الانتواع.[157][158][159][160][161]
بعوضة لندن تحت الأرض
إن بعوضة لندن تحت الأرض هي نوع من أنواع البعوض الموجود في مترو أنفاق لندن. تطورت من الأنواع فوق الأرضية. هذا البعوض وعلى الرغم من اكتشافه لأول مرة في نظام مترو أنفاق لندن، فقد عثر عليه في عدة أنظمة تحت الأرض حول العالم. يُقترح أن هذا البعوض ربما قد تكيف مع الأنظمة تحت الأرضية التي يبنيها الإنسان منذ القرن الماضي وهو من سلالة البعوض المحلي فوق سطح الأرض، على الرغم من أن الأدلة الحديثة تشير إلى أنه صنف من البعوض الجنوبي الذي تكيف مع المساحات الدافئة تحت الأرض في المدن الشمالية.[162]
يملك النوعان سلوكيات مختلفة بشكل كبير [236] مما يجعل من الصعب جدًا تزاوجها،[163] وتملك معدل تكرار أليلي مختلف يدل على الانحراف الوراثي الذي حدث خلال نشأتها.[164] تتكاثر هذه البعوضة على مدار العام ولا تتحمل الاجواء الباردة، وتلدغ القوارض والفئران والبشر، على عكس الأنواع الموضحة أعلاه التي تتحمل البرودة، وتقوم بالسبات في فصل الشتاء وتلدغ الطيور فقط. عندما تم تهجين الصنفين كانت البويضات تعاني من العقم مما يشير إلى وجود عزلة تكاثرية.[165]
تشير البيانات الوراثية إلى أن بعوضة لندن تحت الأرضية تملك أصلًا مشتركًا ولا ترتبط كل مجموعة في كل محطة بأقرب مجموعة بعوض فوق سطح الأرض. افترضت ابحاث بيرن ونيكولز أن التكيف مع البيئة تحت الأرضية حدث محليًا في لندن مرة واحدة فقط. تتميز هذه المجموعات المنفصلة والمنتشرة بشكل واسع باختلافات جينية طفيفة للغاية مما يوحي بأن شكل البعوض قد تطور: يوجد اختلاف واحد جيني مشترك بين مجموعات البعوض تحت الأرضية في عشر مدن روسية،[166] ويوجد اختلاف واحد ثابت في التكرارات المترادفة القصيرة في المجموعات المنتشرة في أوروبا واليابان وأستراليا والشرق الأوسط وجزر الأطلسي.[162]
القريدس العضاض وبرزخ بنما
يوجد نقاش لتحديد متى تم إغلاق برزخ بنما. تدعم الكثير من الأدلة إغلاقه منذ ما يقارب من 2.7 إلى 3.5 مليون عام وهي «... أدلة متعددة ودراسات تحليلية مستقلة».[167] ومع ذلك تشير دراسة حديثة إلى وجود جسر عابر بشكل مسبق منذ ما بين 13 إلى 15 مليون عام.[168] بغض النظر عن توقيت تشكل واغلاق البرزخ يمكن لعلماء الأحياء دراسة الأنواع على جانبي المحيط الهادئ ومنطقة البحر الكاريبي فيما يسمى «واحدة من أعظم التجارب الطبيعية للتطور.» [167] قدمت دراسات عن القريدس العضاض أدلة مباشرة على حدوث الانتواع متباين الموطن،[169] وساهمت في اغناء المؤلفات المتعلقة بمعدلات التطور الجزيئي.[167] تدعم عمليات إعادة التركيب الشكلي باستخدام «مجموعات بيانات متعددة وأساليب تحليلية مترابطة» العلاقات بين الأنواع في المجموعة[170] وتدعم تقنيات الساعة الجزيئية حدوث فصل بين 15 زوجًا من أنواع القريدس منذ ما بين 3 و 15 مليون عام.[167]
النباتات
كان عالم النبات فيرني غرانت رائداً في مجال انتواع النباتات من خلال أبحاثه ومنشوراته الهامة حول هذا الموضوع.[171] وكما قال سابقا: يعتمد العديد من علماء الأحياء على مفهوم الأنواع البيولوجية، ويوجد بعض الباحثين الحديثين اللذين يستخدمون مفهوم الأنواع التطورية. يوجد نقاش حول إطار العمل الذي يجب تطبيقه في البحث.[171] وبغض النظر عن ذلك تعتبر العزلة الإنجابية هي العامل الرئيسي في عملية الانتواع، وقد درسها علماء الأحياء على نطاق واسع.
اكتشف ان كل من التهجين وتعدد الصيغ الصبغية قد لعبا دورا هاما كمساهمين رئيسيين في انتواع النباتات.[172] وكان «حدوث التهجين أكثر تكرارا بكثير مما كان يعتقد سابقا».[171] وبالإضافة إلى هذين الحدثين اللذين يؤديان إلى حدوث الانتواع أصبحت التفضيلات والانعزال وإعادة ترتيب الصبغيات والانتقاء الطبيعي من الأمور الحاسمة في انتواع النباتات. وبالإضافة إلى ذلك تشير الأبحاث الحديثة إلى أن الاصطفاء الجنسي وعوامل التخلق وإنشاء تراكيب أليلية غير متوافقة والتي تسببت في موازنة الانتقاء ساهمت أيضًا في تكوين أنواع جديدة. [172] تم اجراء دراسة على ذلك في كل من المختبر والطبيعة. وتشير الدراسات أيضًا إلى أنه بسبب «الطبيعة الثابتة للنباتات (لا تغير مكانها) ... تزيد أهمية الانتواع البيئي ...»[172]
يؤدي التهجين بين نوعين مختلفين في بعض الأحيان إلى ظهور نمط الظاهري مختلف. يمكن أن يكون هذا النمط الظاهري أكثر تجانسًا من السلالات الأبوية وبالتالي فمن الممكن ان يفضل الانتقاء الطبيعي هؤلاء الأفراد. وفي النهاية: إذا تم تحقيق العزلة الإنجابية سيسبب ذلك حدوث نوع منفصل. ومع ذلك يصعب تحقيق العزلة الإنجابية بين الانواع الهجينة والآباء بشكل خاص، وبالتالي تعد الأنواع الهجينة حدثًا نادرًا. ومع ذلك يُعتبر التهجين الذي يسبب العزلة التناسلية وسيلة هامة لحدوث الانتواع في النباتات[173] نظرًا لأن النباتات يمكن ان تحتوي على تعدد الصيغ الصبغية (امتلاك أكثر من نسختين من كل كروموسوم) بسهولة أكبر من الحيوانات.[174]
يعتبر تعدد الصيغ الصبغية مهما في الكائنات الهجينة لأنها تسمح لها بالتكاثر، حيث توجد مجموعتين مختلفتين من الصبغيات التي تكون كل منهما قادرة على الاقتران مع شريك مماثل لها خلال الانقسام المنصف.[175][176] يشتمل تعدد الصيغ الصبغية أيضًا على المزيد من التنوع الجيني مما يسمح للكائنات بتجنب انحدار زواج الأقارب في المجتمعات الصغيرة. يطلق على التهجين بدون تغيير في عدد الصبغيات الانتواع الهجين المثلي. يعتبر هذا الانتواع نادرًا جدًا ولكنه يظهر في فراشات هلكونيوس وزهور عباد الشمس. يعتبر الانتواع بسبب تعدد الصيغ الصبغية الذي يحوي تغييرات في عدد الكروموسومات ظاهرة أكثر شيوعًا خاصة في الأنواع النباتية.
إن تعدد الصيغ الصبغية هي آلية تسببت في العديد من أحداث الانتواع السريع بشكل محلي لأن النسل الناتج عن زواج كائن رباعي الصيغ الصبغية مع كائن ثنائي الصيغ الصبغية غالباً ما يكون ذرية عقيمة ثلاثية الصيغ الصبغية.[177] لا يكون كل نسل الكائنات متعددة الصيغ الصبغية معزول تكاثريا عن النباتات الأبوية وقد يستمر تبادل الجينات بينها من خلال - على سبيل المثال - زواج كائنات ثلاثية الصيغ الصبغية مع كائنات ثنائية الصيغ الصبغية والذي ينتج كائنات رباعية الصيغ الصبغية، أو التزاوج بين الاعراس ذات الانقسام المنصف ثلاثية الصيغ الصبغية مع رباعية الصيغ الصبغية. تم طرح فكرة كون العديد من الأنواع الموجودة في معظم النباتات والحيوانات قد خضعت لحدث تعدد الصيغ الصبغية خلال تاريخها التطوري.[178] إن تكاثر الأنواع متعددة الصيغ الصبغية بشكل ناجح يكون أحيانًا غير جنسي بل عن طريق التوالد البكري أو التكاثر اللا اخصابي، وتوجد كائنات متعددة الصيغ الصبغية لأسباب غير معروفة. هناك حالات نادرة لوجود ثدييات متعددة الصيغ الصبغية، ولكن غالبًا ما تؤدي إلى الوفاة قبل الولادة.
يعتبر الباحثون أن العزلة الإنجابية هي مفتاح الانتواع.[179] يتمثل أحد الأقسام الرئيسية لأبحاث الأنتواع في تحديد طبيعة الحواجز التي تحول دون التكاثر. غالبًا ما يعتبر علماء النبات أن التصنيفات الحيوانية للانعزال قبل التزاوج أو بعد التزاوج غير كافية.[179] تعطي الأمثلة التالية شرحًا هامًا لعملية الانتواع.
ميمولوس بيريجرينوس
لوحظ وجود نوع جديد متعدد الصيغ الصبغية من ازهار القردة يسمى (ميمولوس بيريجرينوس) على ضفاف نهر في ليدهيلز جنوب لاناركشاير في اسكتلندا. كان الأبوين هما ازهار القردة الصفراء الشائعة (تحتوي على 14 زوجًا من الصبغيات) وازهار القردة الصفراء المرقطة (تحتوي على 30-31 زوجًا من الصبغيات)، يملك النوع الناتج ست نسخ من الصبغيات (ناتجة عن تكرار الصيغ الصبغية الثلاثية الهجينة والعقيمة). وبسبب طبيعة هذه الأنواع فهي تملك القدرة على الإخصاب الذاتي. ونظرًا للعدد الذي تملكه من الصبغيات فإنه لا يمكنها التزاوج مع ازهار القردة الصفراء الشائعة أو ازهار القردة الصفراء المرقطة أو ذريتهما ثلاثية الصيغ الصبغية العقيمة. إما سيموت نوع ميمولوس بيريجرينوس الناتج، أو لن ينتج ذرية، أو سيتكاثر مع نفسه بشكل فعال مؤديا إلى نشوء نوع جديد.[180][181]
رافانوبراسيكا
تشمل رافانوبراسيكا كل انواع النسل الهجين بين أجيال الفجل والبراسيكا (الملفوف).[182][183] ان الرافانوبراسيكا هو تهجين متعدد الصيغ الصبغية بين الفجل والملفوف. ويعرف شكلين اخرين من أشكال الرافانوبراسيكا الخصبة وهما الراباراديش وهو نبات هجين متعدد الصيغ الصبغية ناتج عن تهجين فجل رافانوس ساتيفوس مع فجل براسيكا رابا ويزرع كمحصول علفي. رافانوفورتي هو نبات هجين متعدد الصيغ الصبغية ناتج عن تهجين براسيكا تورنفور ورافانوس كاوداتوس. يعد نبات رافانوبراسيكا نباتًا مميزا لأنه وعلى الرغم من طبيعته الهجينة فهو ليس عقيمًا. وقد دفع ذلك بعض علماء النبات إلى اقتراح أن آلية تهجين زهرة من خلال حبوب طلع تعود لنوع آخر في الطبيعة يمكن أن يكون آلية الأنتواع الشائعة في أنواع النباتات العليا.
نبات السينيسيو (الشيخة)

إن نباتات الشيخة الويلزية هي نباتات متعددة الصيغ الصبغية، وتحتوي على مجموعات من الصبغيات تنشأ من نوعين مختلفين. كانت النبات الأبوة (سينيسيو باكستيري) هجينا عقيمة يمكن أن ينشأ تلقائيًا عندما ينمو نبات الشيخة (سينيسيو فولغاريس) ونبات سينيسيو سكوالديوس جنبًا إلى جنب. أدى تضاعف مفاجئ في عدد الصبغيات في نبات سينيسيو باكستيري في بداية القرن العشرين إلى تكوين نوع جديد خصب.[184][185]
سينيسيو إيبوراسينسيس هو نوع نبات هجين من سينيسيو سكواليدوس الذي لا يتكاثر ذاتيا وسينيسيو فولغاريس الذي يتكاثر ذاتيًا (المعروف باسم نبات الشيخة الشائع). تمتع نبات سينيسيو إيبوراسينسيس بميزة التكاثر بشكل ذاتي مثل سينيسيو فولغاريس. ومع ذلك فهو يظهر تشابه قليل أو غير موجود مع الأنواع الأم وبالتالي فهو معزول تكاثريًا، مما يشير إلى وجود حواجز تكاثر قوية بين هذا النوع الهجين الجديد والابوين. وقد نتج عن التهجين الأبوي بين النوع الهجين مع النباتات الأبوية نوع سينيسيو فولغاريس. إن موطن سينيسيو فولغاريس الأصلي هو بريطانيا، بينما اكتشفت نباتات سينيسيو سكواليدوس في صقلية في أوائل القرن الثامن عشر، وظهر نوع سينيسيو إيبوراسينسيس نتيجة لتهجين هذين النوعين خلال الـ 300 عام الماضية.
تم اجراء دراسة على الانواع الهجينة الأخرى المنحدرة من نفس الوالدين، وكان بعضها مصاب بالعقم مثل سينيسيو باكستيري. ويوجد أيضا أنواع هجينة خصبة أخرى مثل نباتات الشيخة الشائعة الآن في بريطانيا، وسينيسيو كامبرينسيس متعدد الصيغ الصبغية والذي ربما نشأ (وفقًا للأدلة الجزيئية) بشكل مستقل في ثلاث مواقع مختلفة على الأقل. تدعم الأدلة الشكلية والوراثية اعتبار سينيسيو إيبوراسينسيس نوعا منفصلا عن الأنواع الهجينة الأخرى المعروفة.[186]
نبات رشاد أذن الفأر
اكتشفت كريستين بومبليز وعلماء اخرون من معهد ماكس بلانك لعلوم الأحياء التطورية اثنين من الجينات الموجودة في نبات رشاد أذن الفأر. عندما يرث النسل كلا الجينين فإنه يسبب رد فعل في النبات الهجين يحول نظام المناعة الخاص به ضده. لم تكن الجينات ضارة في الوالدين لكنها تطورت بشكل منفصل لتتفاعل بشكل سيء عندما تكون مجتمعة.[187] ولاختبار ذلك قام العلماء بتهجين 280 سلالة مختلفة وراثيا من النبات من خلال 861 طريقة مختلفة ووجدوا أن 2% من السلالة الهجينة الناتجة كانت تحول نظام المناعة الخاص بها ضدها. وتشاركت النباتات الـ 20 أيضًا في مجموعة جينية متماثلة في مجموعة من 1080 جينًا. اكتشفت بومبليز انه كان مطلوبا في جميع الحالات تقريبًا وجود اثنين من الجينات ليتسببوا في حدوث استجابة المناعة ضد النبات ذاته. قامت بومبليز بدراسة نبات هجين واحد بدقة ووجدت أن أحد الجينين ينتمي إلى فئة (NB-LRR) وهي مجموعة شائعة من جينات مقاومة الأمراض التي تساهم في التعرف على الإصابات الجديدة. عندما أزالت بومبليز الجين المسبب للمشاكل نما النبات الهجين بشكل طبيعي. يمكن أن يؤدي عدم التوافق على مدى الأجيال المتعاقبة إلى حدوث انقسامات بين سلالات النباتات المختلفة، مما يقلل من فرص نجاح التزاوج ويحول السلالات المختلفة إلى أنواع منفصلة.[188]
نبات لحية التيس
يمثل نبات لحية التيس مثالا على حدوث الأنتواع الهجين. أدخل البشر ثلاثة أنواع من نبات لحية التيس إلى أمريكا الشمالية في أوائل القرن العشرين. هذه الأنواع هي نبات لحية التيس الغربي، ونبات لحية التيس في المروج، ونبات المحار وهي الآن نباتات شائعة في الاراضي ضمن المدن. وجد علماء النبات في خمسينيات القرن العشرين نوعين جديدين في ولايتي أيداهو وواشنطن، حيث كانت توجد الأنواع الثلاثة المعروفة مسبقا. أحد الأنواع الجديدة هو نبات سلسفي موسكو وهو هجين رباعي الصيغة الصبغية ناتج عن نبات لحية التيس الغربي ونبات لحية التيس في المروج. من الأنواع الجديدة الأخرى نبات لحية التيس ميروس هو أيضًا متعدد الصيغ الصبغية، لكن أسلافه كانوا نبات لحية التيس الغربي ونبات المحار. يشار عادة إلى هذه الأنواع الجديدة باسم (أنواع أوينباي الهجينة) نسبة لعالم النبات الذي اكتشفها أول مرة. ينمو نبات لحية التيس ميروس بشكل رئيسي عن طريق إعادة إنتاج أفراده مجددا، ولكن تستمر احداث التهجين الإضافية في زيادة أعداده.[189]
تزاوج نبات لحية التيس الغربي مع نبات لحية التيس في المروج في أوروبا لكنهما لم يكونا قادرين على التهجين. وجدت دراسة نشرت في آذار / مارس عام 2011 أنه عندما تم إدخال هذين النباتين إلى أمريكا الشمالية في العشرينيات من القرن الماضي قاما بالتزاوج ومضاعفة عدد الصبغيات الموجودة في نبات لحية التيس موسكو الهجين الناتج مما سمح بإعادة ضبط جيناتها والذي أدى بدوره إلى حدوث تباين وراثي أكبر. قال البروفيسور دوج سولتيس من جامعة فلوريدا «لقد رأينا عملية التطور أثناء حدوثها ... قد تساعد أنماط جديدة ومتنوعة من الجينيات الأنواع الجديدة على التكيف السريع في بيئات جديدة».[190][191] تعزز هذه الملاحظات (حدوث الأنتواع من خلال التهجين) الأدلة على السلف المشترك للكائنات الحية والإطار الزمني الذي نشأت فيه الأنواع الجديدة في بيئتها الجديدة. تتم إعادة عملية التهجين بشكل مصطنع في المختبرات منذ عام 2004 إلى يومنا هذا.
الفقاريات
طائر أبو قلنسوة
تعيش أنواع طيور أبو قلنسوة في ألمانيا وتطير نحو الجنوب الغربي إلى إسبانيا بينما تطير مجموعة صغيرة منها إلى الشمال الغربي نحو بريطانيا العظمى خلال الشتاء. وجد جريجور رولسهاوزن من جامعة فرايبرغ أن الفصل الجيني بين المجموعتين يتم حاليا بالفعل. ظهرت الاختلافات الموجودة خلال نحو 30 جيلًا. يمكن تحديد المجموعة التي ينتمي اليها كل فرد بدقة 85٪ باستخدام تسلسل الحمض النووي. ذكر العالم ستيوارت بيروب من جامعة إكزتر أن الطيور التي تسافر في فصل الشتاء نحو إنجلترا تميل إلى التزاوج فيما بينها فقط ولا تتزاوج عادة مع التي تسافر في فصل الشتاء نحو مناطق البحر الأبيض المتوسط.[192] لا يزال من غير المنطقي أن نقول أنّ المجموعتين ستصبحان نوعين مختلفين، لكن الباحثين يتوقعون ذلك بسبب استمرار الفصل الوراثي والجغرافي. [193]
سمك المولي
سمكة المولي هي سمكة صغيرة تعيش في كهوف الكبريت في المكسيك. وجدت الدراسات التي أجريت على هذا النوع أن مجموعتين مختلفتين من اسماك المولي (الأسماك الداكنة التي توجد في الاعماق وأسماك المياه السطحية فاتحة اللون) تصبح أكثر اختلافًا وراثياً.[194] لا تملك المجموعتين حاجز واضح يفصل بين الاثنين، ومع ذلك فقد وجد أن أسماك المولي يتم اصطيادها بواسطة بقيات الماء الكبيرة. قام توبلر بجمع بقيات الماء الكبيرة وكلا النوعين من اسماك المولي ووضعهما في زجاجات بلاستيكية كبيرة ثم وضعهما في الكهف. تبين بعد مضي يوم واحد أن الأسماك المتكيفة مع الكهف والتي وضعت في الضوء تعرضت لأكبر قدر من الضرر، حيث أصابت نسبة أربع من كل خمس هجمات قامت بها بقيات الماء الكبيرة اجزاء من فم الاسماك. سَرَت الأحداث في الظلام عكس ذلك. تستطيع حواس أسماك المولي اكتشاف تهديد المفترس في بيئتها وليس في المناطق الأخرى. إنَّ الانتقال من بيئة إلى أخرى يزيد بشكل كبير من خطر الموت. يخطط توبلر لإجراء مزيد من التجارب، وهو يعتقد أن هذه التجربة مثال جيد على ظهور نوع جديد.[195]
الدب القطبي
إنَّ الانتقاء الطبيعي والعزلة الجغرافية وعملية الانتواع تكون واضحة في العلاقة بين الدب القطبي والدب البني. حيث تعتبر أنواعًا منفصلة في مناطقها،[196] ومع ذلك تم توثيق أنها تملك القدرة على التزاوج فيما بينها وإنتاج ذرية خصبة. حدث هذا التهجين الأبوي المتكرر في البرية وفي الدبب المأسورة لدى الإنسان على حد سواء وتم توثيقه[197] والتحقق منه من خلال اختبار الحمض النووي.[198] يرجع أقدم دليل أحفوري معروف للدببة القطبية إلى نحو 130000 إلى 110000 عام،[199] ومع ذلك قدرت البيانات الجزيئية أوقات مختلفة لزمن حدوث الانتواع. قدر تحليل الحمض النووي للمتقدرات حدوث الانتواع قبل 150000 عام[199] في حين أظهر تحليل الشريط الوراثي النووي حدوث اختلاف تقريبي قبل 603000 عام.[200] أثبتت الأبحاث الحديثة باستخدام شريط وراثي كامل (بدلاً من الدنا المتقدرة أو الشريط الوراثي النووي الجزئي) اختلاف الدببة القطبية والبنية منذ ما بين 343 - 479 ألف عام.[201] وعلى الرغم من الاختلافات على تحديد اوقات التباين تشير الأبحاث الجزيئية إلى أن الأنواع الشقيقة خضعت لعمليات انتواع وتزاوج معقدة للغاية.[202]
اكتسبت الدببة القطبية اختلافات تشريحية ووظيفية كبيرة عن الدببة البنية مما سمح لها بالبقاء على قيد الحياة بشكل مريح في ظروف لا تستطيع الدببة البنية تحملها. من الأمثلة البارزة: القدرة على السباحة لمدة ستين ميلًا أو أكثر بشكل متواصل في المياه المتجمدة، وامتزاج الفراء الابيض مع الثلج، بقاء الدب دافئًا في البيئة القطبية الشمالية، رقبة طويلة تجعل من السهل على الدببة إبقاء رؤوسها فوق الماء أثناء السباحة، وامتلاك أرجل مكشوفة كبيرة متصلة تعمل كمجاديف عند السباحة. كما طورت حليمات صغيرة وفجوات شفط على باطن القدمين لجعلها أقل عرضة للانزلاق على الجليد، إلى جانب امتلاك اذنين أصغر للحد من فقدان الحرارة، واجفان تعمل مثل النظارات الشمسية، والتأقلم مع نظام غذائي يتكون من اللحوم فقط، سعة معدة كبيرة لتستطيع الدببة الاكل عندما تسمح لها الفرصة، والقدرة على الصيام لمدة تصل إلى تسعة أشهر أثناء إعادة تدوير مادة البولة (اليوريا).[203][204] يقدم هذا المثال تغييراً تطوريًا كبيرًا يحتوي عدة مجالات في علم الأحياء التطورين على سبيل المثال: التكيف من خلال الانتقاء الطبيعي والعزلة الجغرافية والانتواع والتهجين.
أدلة من التلون

قدم تلون الحيوانات دليلًا مبكرًا مهمًا على التطور عن طريق الاصطفاء الطبيعي، في الوقت الذي لم يتوفر فيه أي دليل مباشر. تم اكتشاف ثلاث وظائف رئيسية للتلون في النصف الثاني من القرن التاسع عشر وتم استخدامها لاحقًا كدليل على حدوث الاصطفاء: التمويه (التلون الوقائي)، والتنكر مثل: الباتيسيان والمولريان، والتحذير اللوني. سعى علماء الطبيعة بما في ذلك المعاصرين لداروين مثل هنري والتر بيتس وفريتز مولر بعد الأدلة المفصلة التي قدمها داروين في كتابه (أصل الأنواع) ونظراً لعدم وجود آليات لتحليل التغير الوراثي أو الجينات في ذلك الوقت إلى الحصول على أدلة مما يمكنهم ملاحظته في الطبيعة.[206]
التنكر والتحذير اللوني
وصف بيتس ومولر أشكال التنكر (والتي حملت الآن أسماءهم) بناءً على ملاحظاتهم عن الفراشات الاستوائية.
تم شرح هذه الأنماط الدقيقة من التلون بسهولة عن طريق الاصطفاء الطبيعي لأن الحيوانات المفترسة مثل الطيور التي تصطاد من خلال البصر سوف تصطاد وتقتل في أغلب الأحيان الحشرات التي تملك نماذج تنكر أقل كفاءة من تلك التي تتنكر بكفاءة عالية، لكن يصعب تفسير هذه الأنماط. سعى العلماء الذين اتبعوا نهج داروين مثل ألفريد راسل والاس وإدوارد باغنال بولتون، وهيو كوت وبرنارد كيتولويل في القرن العشرين إلى إثبات حدوث الاصطفاء الطبيعي.[207][208]
التمويه
لاحظ والاس في عام 1889 أن التمويه الثلجي وخاصة الريش والفرو التي تغيرت مع المواسم، واقترح تفسيرًا واضحًا لها هو التكيف للاختفاء.[209][205] استخدم كتاب بولتون لعام 1890 الذي حمل عنوان (ألوان الحيوانات) جميع أشكال التلون للدفاع عن فكرة الاصطفاء الطبيعي.[210] وصف كوت أنواعًا كثيرة من التمويه والتنكر والتلون التحذيري في كتابه الذي نشر عام 1940 تحت عنوان (التلون التكيفي في الحيوانات) ولا سيما رسوماته الخاصة بالتلون المتزامن في الضفادع، وأقنع علماء الأحياء الآخرين بأن هذه السمات المخادعة كانت نتاجًا للاصطفاء الطبيعي.[207] لاحظت العالمة كيتلويل تطور العث المفلفل، ووضحت أن هذا النوع قد تكيف مع تغير التلوث في البيئة، قدم ذلك دليلًا دامغًا على التطور الدارويني.[208]
أدلة من النمذجة الرياضية
تسمح علوم الحاسوب بدراسة تكرار الأنظمة المعقدة المتغيرة ذاتياً، مما يقدم شرح رياضي لطبيعة العمليات التي تقف وراء عملية التطور، وتقدم أدلة على الأسباب الخفية للأحداث التطورية المعروفة. يمكن دراسة تطور الآليات الخلوية المحددة مثل جسيمات التضفير التي يمكن أن تحول الشريط الوراثي في الخلية إلى ورشة عمل واسعة تمتلك مليارات الأجزاء القابلة للتبديل والتي يمكن أن تشكل الأدوات التي تخلق الانسان بطريقة دقيقة.
«استغرقت الدراسة أكثر من خمسة عقود، لكن أصبح الحاسوب الإلكتروني الآن قويًا بدرجة كافية لمحاكاة التطور»،[211] وهو يساعد في مجال المعلوماتية الحيوية محاولا حل المشكلات البيولوجية.
أتاحت البيولوجيا التطورية الحاسوبية للباحثين تتبع تطور عدد كبير من الكائنات الحية من خلال قياس التغيرات في الحمض النووي الخاص بهم، وليس من خلال التصنيف الفيزيائي أو الملاحظات الجسدية وحدها. حيث تمت مقارنة الأشرطة الوراثية بأكملها مما سمح بدراسة الأحداث التطورية المعقدة مثل تكرار الجينات ونقل الجينات الأفقي والتنبؤ بالعوامل المهمة في عملية الانتواع. وقد ساعد أيضًا في بناء نماذج حسابية معقدة من المجموعات الحيوية من أجل التنبؤ بنتيجة النظام الطبيعي مع مرور الوقت وتتبع وتبادل المعلومات حول عدد كبير متزايد من الأنواع والكائنات الحية.
يسعى العلماء في المستقبل لإعادة بناء شجرة الحياة المعقدة.
أوضح كريستوف أدامي الأستاذ بمعهد كيك للخريجين هذه النقطة في (تطور التعقيد الحيوي):
- لإثبات أو دحض توجه ما في تطور التعقيد الحيوي يجب تحديد التعقيد بدقة وقياسه. يحدد تعريف نظري حديث للمعلومات التسلسل الجيني مع كمية المعلومات التي يخزنها حول بيئته. نحن نبحث في تطور التعقيد الحيوي في مجموعات الكائنات الممثلة حاسوبيا ونراقب بالتفصيل التحولات التطورية التي تزيد من التعقيد. وتبين أنه نظرًا لأن الانتقاء الطبيعي يجبر السلاسل الوراثية على التصرف كتجربة (عفريت ماكسويل) طبيعية داخل بيئة ثابتة، يؤدي ذلك إلى زيادة التعقيد الحيوي. .[212]
أوضح كل من ديفيد يارل وميشيل ديم (أساتذة في جامعة رايس) هذه النقطة في كتاباتهم تحت عنوان (قابلية التطور هي سمة قابلة للاختيار):
- لم تتطور الحياة فقط، ولكنها تطورت لتتطور. وتطورت بذلك الارتباطات داخل بنية البروتين، وتطورت آليات لتغيير هذه الارتباطات أيضا. إن المعدّلات التي تحدث فيها الأحداث المختلفة داخل التسلسل الهرمي للحركات التطورية ليست عشوائية أو اعتباطية ولكن يتم اختيارها بواسطة التطور الدارويني. بشكل منطقي: يؤدي التغير البيئي السريع أو الشديد إلى حدوث الاصطفاء وبالتالي حدوث المزيد من التطوير. لا تمنع السببية حدوث هذا الاصطفاء، وهو الأقوى في مقياس التغيرات الكبرى ضمن التسلسل الهرمي للطفرات. يتم تفسير العديد من النتائج داخل البيولوجيا التطورية والتي تعتبر حتى الآن من الحوادث التطورية عن طريق الاصطفاء لقابلية التطور. على سبيل المثال: يوضح الجهاز المناعي للفقاريات أن البيئة المتغيرة لمولدات الضد قد سببت ضغوطا انتقائية لاستخدام الكودونات القابلة للتكيف والبوليميرات منخفضة الدقة أثناء فرط التطفر. لوحظ وجود قوة دافعة مماثلة لاستخدام الكودون نتيجة لارتفاع معدلات الطفرات المنتجة في بروتين الهيماغلوتينين لفيروس إنفلونزا أ.[213]
«أظهرت المحاكاة الحاسوبية لتطور التسلسلات الخطية أهمية إعادة تجميع بنيات السلاسل بدلاً من الطفرات النقطية وحدها. يجب أن تسمح الدورات المتكررة للطفرات النقطية وإعادة التركيب والاصطفاء بالتطور الجزيئي للسلاسل المعقدة في المختبر مثل سلاسل البروتينات».[214] تشتمل الهندسة الجزيئية التطورية (تُسمى أيضًا التطور الموجه أو التطور الجزيئي في المختبر) على دورات متكرّرة للطفرات والتضاعف وإعادة التركيب واختيار أصلح الجزيئات الفردية (البروتينات والحمض النووي الريبوزي منقوص الأكسجين والحمض النووي الريبوزي). يمكن إعادة إحياء التطور الطبيعي لنرى المسارات المحتملة من الدورات التحفيزية القائمة على البروتينات وعلى الحمض النووي الريبوزي وعلى الحمض النووي الريبوزي منقوص الأكسجين.[214][215][216][217]
المصادر
- Mount DM. (2004). Bioinformatics: Sequence and Genome Analysis (الطبعة الثانية). Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY. .
- Douglas J. Futuyma (1998). Evolutionary Biology (الطبعة الثالثة). Sinauer Associates Inc. صفحات 108–110. .
- Haszprunar (1995). "The mollusca: Coelomate turbellarians or mesenchymate annelids?". In Taylor (المحرر). Origin and evolutionary radiation of the Mollusca : centenary symposium of the Malacological Society of London. Oxford: Oxford Univ. Press. .
- Kozmik, Z; Daube, M; Frei, E; Norman, B; Kos, L; Dishaw, LJ; Noll, M; Piatigorsky, J (2003). "Role of Pax genes in eye evolution: A cnidarian PaxB gene uniting Pax2 and Pax6 functions" ( كتاب إلكتروني PDF ). Developmental cell. 5 (5): 773–85. doi:10.1016/S1534-5807(03)00325-3. PMID 14602077. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 24 سبتمبر 2015.
- Kozmik, Z; Daube, Michael; Frei, Erich; Norman, Barbara; Kos, Lidia; Dishaw, Larry J.; Noll, Markus; Piatigorsky, Joram (2003). "Role of Pax Genes in Eye Evolution A Cnidarian PaxB Gene Uniting Pax2 and Pax6 Functions". Developmental Cell. 5 (5): 773–785. doi:10.1016/S1534-5807(03)00325-3. PMID 14602077.
- Land, M.F. and Nilsson, D.-E., Animal Eyes, Oxford University Press, Oxford (2002) .
- Chen FC, Li WH (2001). "Genomic Divergences between Humans and Other Hominoids and the Effective Population Size of the Common Ancestor of Humans and Chimpanzees". Am J Hum Genet. 68 (2): 444–56. doi:10.1086/318206. PMC . PMID 11170892.
- Cooper GM, Brudno M, Green ED, Batzoglou S, Sidow A (2003). "Quantitative Estimates of Sequence Divergence for Comparative Analyses of Mammalian Genomes". Genome Res. 13 (5): 813–20. doi:10.1101/gr.1064503. PMC . PMID 12727901.
- The picture labeled "Human Chromosome 2 and its analogs in the apes" in the article Comparison of the Human and Great Ape Chromosomes as Evidence for Common Ancestry is literally a picture of a link in humans that links two separate chromosomes in the nonhuman apes creating a single chromosome in humans. Also, while the term originally referred to fossil evidence, this too is a trace from the past corresponding to some living beings which when alive were the physical embodiment of this link. نسخة محفوظة 06 سبتمبر 2017 على موقع واي باك مشين.
- The The New York Times report Still Evolving, Human Genes Tell New Story, based on A Map of Recent Positive Selection in the Human Genome, states the International HapMap Project is "providing the strongest evidence yet that humans are still evolving" and details some of that evidence. نسخة محفوظة 30 يناير 2018 على موقع واي باك مشين.
- Woese C, Fox G (1977). "Phylogenetic structure of the prokaryotic domain: the primary kingdoms". Proc Natl Acad Sci USA. 74 (11): 5088–90. Bibcode:1977PNAS...74.5088W. doi:10.1073/pnas.74.11.5088. PMC . PMID 270744.
- Woese C, Kandler O, Wheelis M (1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". Proc Natl Acad Sci USA. 87 (12): 4576–9. Bibcode:1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC . PMID 2112744. مؤرشف من الأصل في 27 يونيو 200811 فبراير 2010.
- "29+ Evidences for Macroevolution: The Scientific Case for Common Descent". Theobald, Douglas. مؤرشف من الأصل في 15 يونيو 201910 مارس 2011.
- "Converging Evidence for Evolution." Phylointelligence: Evolution for Everyone. Web. 26 Nov. 2010. نسخة محفوظة 27 يوليو 2011 على موقع واي باك مشين.
- Petrov DA, Hartl DL (2000). "Pseudogene evolution and natural selection for a compact genome". J Hered. 91 (3): 221–7. doi:10.1093/jhered/91.3.221. PMID 10833048.
- Junk DNA: Science Videos – Science News. ScienCentral (2004-05-06). Retrieved on 2011-12-06.نسخة محفوظة 08 مارس 2012 على موقع واي باك مشين.
- Okamoto N, Inouye I (2005). "A secondary symbiosis in progress". Science. 310 (5746): 287. doi:10.1126/science.1116125. PMID 16224014.
- Okamoto N, Inouye I (2006). "Hatena arenicola gen. et sp. nov., a katablepharid undergoing probable plastid acquisition". Protist. 157 (4): 401–19. doi:10.1016/j.protis.2006.05.011. PMID 16891155.
- Human Chromosome 2 is a fusion of two ancestral chromosomes by Alec MacAndrew; accessed 18 May 2006. نسخة محفوظة 26 ديسمبر 2017 على موقع واي باك مشين.
- Evidence of Common Ancestry: Human Chromosome 2 على يوتيوب(video) 2007
- Yunis and Prakash; Prakash, O (1982). "The origin of man: a chromosomal pictorial legacy". Science. 215 (4539): 1525–1530. doi:10.1126/science.7063861. PMID 7063861.
- Human and Ape Chromosomes; accessed 8 September 2007. نسخة محفوظة 06 سبتمبر 2017 على موقع واي باك مشين.
- Avarello, Rosamaria; Pedicini, A; Caiulo, A; Zuffardi, O; Fraccaro, M (1992). "Evidence for an ancestral alphoid domain on the long arm of human chromosome 2". Human Genetics. 89 (2): 247–9. doi:10.1007/BF00217134. PMID 1587535.
- Ijdo, J. W.; Baldini, A; Ward, DC; Reeders, ST; Wells, RA (1991). "Origin of human chromosome 2: an ancestral telomere-telomere fusion". Proceedings of the National Academy of Sciences. 88 (20): 9051–5. doi:10.1073/pnas.88.20.9051. PMC . PMID 1924367.
- Pallen, Mark (2009). Rough Guide to Evolution. Rough Guides. صفحات 200–206. .
- Belshaw, R ; Pereira V; Katzourakis A; Talbot G; Paces J; Burt A; Tristem M. (2004). "Long-term reinfection of the human genome by endogenous retroviruses". Proc Natl Acad Sci USA. 101 (14): 4894–99. doi:10.1073/pnas.0307800101. PMC . PMID 15044706.
- Bonner TI; et al. (1982). "Cloned endogenous retroviral sequences from human DNA". Proceedings of the National Academy of Sciences. 79 (15): 4709–13. doi:10.1073/pnas.79.15.4709. PMC . PMID 6181510.
- Van Der Kuyl, AC; Dekker, JT; Goudsmit, J (1999). "Discovery of a New Endogenous Type C Retrovirus (FcEV) in Cats: Evidence for RD-114 Being an FcEVGag-Pol/Baboon Endogenous Virus BaEVEnv Recombinant". Journal of Virology. 73 (10): 7994–8002. PMC . PMID 10482547.
- TalkOrigins Archive. "29+ Evidences for Macroevolution: Part 2". مؤرشف من الأصل في 28 سبتمبر 201808 نوفمبر 2006.
- Lambert, Katie. (2007-10-29) HowStuffWorks "How Atavisms Work". Animals.howstuffworks.com. Retrieved on 2011-12-06. نسخة محفوظة 17 سبتمبر 2011 على موقع واي باك مشين.
- JPG image - تصفح: نسخة محفوظة 24 مارس 2016 على موقع واي باك مشين.
- Evolutionary Atavisms. Edwardtbabinski.us. Retrieved on 2011-12-06. نسخة محفوظة 10 يناير 2017 على موقع واي باك مشين.
- Tyson, Reid; Graham, John P.; Colahan, Patrick T.; Berry, Clifford R. (يوليو 2004). "Skeletal Atavism in a Miniature Horse". Veterinary Radiology & Ultrasound. 45 (4): 315–317.
- Biello, David (2006-02-22). "Mutant Chicken Grows Alligatorlike Teeth". ساينتفك أمريكان. مؤرشف من الأصل في 10 أبريل 202008 مارس 2009. نسخة محفوظة 19 نوفمبر 2007 على موقع واي باك مشين.
- Domes, Katja; Norton, Roy A.; Maraun, Mark; Scheu, Stefan (2007-04-24). "Reevolution of sexuality breaks Dollo's law". PNAS. 104 (17): 7139–7144. doi:10.1073/pnas.0700034104. PMC . PMID 17438282. مؤرشف من الأصل في 4 أبريل 201908 أبريل 2009.
- Held, Lewis I. (2010). "The Evo-Devo Puzzle of Human Hair Patterning". Evolutionary Biology. 37 (2–3): 113. doi:10.1007/s11692-010-9085-4.
- Douglas J. Futuyma (1998). Evolutionary Biology (الطبعة الثالثة). Sinauer Associates Inc. صفحة 122. .
- 29+ Evidences for Macroevolution: Part 1. Talkorigins.org. Retrieved on 2011-12-06. نسخة محفوظة 11 ديسمبر 2017 على موقع واي باك مشين.
- Coyne, Jerry A. (2009). Why Evolution is True. Viking. صفحات 8–11. .
- Charles Darwin (1859). On the Origin of Species. John Murray. صفحة 420.
- Natan Slifkin (2006). The Challenge of Creation... Zoo Torah. صفحات 258–9. .
- Coyne, Jerry A. (2009). Why Evolution Is True. Viking. صفحات 69–70. .
- Mary Jane West-Eberhard (2003). Developmental plasticity and evolution. Oxford University Press. صفحة 232. .
- "Example 1: Living whales and dolphins found with hindlimbs". Douglas Theobald. مؤرشف من الأصل في 28 سبتمبر 201820 مارس 2011.
- Mark Ridley (2004). Evolution (الطبعة الثالثة). Blackwell Publishing. صفحة 282. . مؤرشف من الأصل في 10 أبريل 2020.
- Dawkins, Richard (2009). The Greatest Show on Earth: The Evidence for Evolution. Bantam Press. صفحات 364–365. .
- ورقة ولف لونينغ، العصب الراجع في الزرافة - تصفح: نسخة محفوظة 4 أبريل 2016 على موقع واي باك مشين.
- دراسة حالة عصب حنجري غير ملتف - تصفح: نسخة محفوظة 31 مايو 2018 على موقع واي باك مشين.
- grays anatomy، الطبعة 40، ص. ،459، 588، 589.
- grays anatomy، الطبعة 39، ص. 1053
- Williams, G.C. (1992). Natural selection: domains, levels, and challenges. Oxford Press. .
- Coyne, Jerry A. (2009). Why Evolution is True. Viking. صفحات 26–28. .
- "Confessions of a Darwinist". Niles Eldredge. مؤرشف من الأصل في 24 ديسمبر 201322 يونيو 2010.
- Laboratory 11 – Fossil Preservation, by Pamela J. W. Gore, Georgia Perimeter College نسخة محفوظة 1 أبريل 2012 على موقع واي باك مشين.
- "Frequently Asked Questions". The Natural History Museum of Los Angeles County Foundation. مؤرشف من الأصل في 7 ديسمبر 201121 فبراير 2011.
- William Richard John Dean and Suzanne Jane Milton (1999). The Karoo: ecological patterns and processes. Cambridge University Press. صفحة 31. [[Special:BookSources/0-521-55430-0|0-521-55430-0 [[تصنيف:مقالات ذات أرقام كتب دولية غير صالحة]]]].
- Robert J. Schadewald (1982). "Six "Flood" Arguments Creationists Can't Answer". Creation Evolution Journal. 3: 12–17. مؤرشف من الأصل في 19 سبتمبر 2015.
- "من الواضح أنه من اللازم وجود أسلاف للفقاريات عاشت في العصر الكامبري، ولكن افتُرض أنها أسلاف لافقارية للفقاريات الحقيقية — أي حبليات أولية. وقد رُوج كثيرًا للبيكايا أنه أقدم أحفورة للحبليات الأولية." رتشرد دوكنز 2004 {{نسخ:مترجمة|:en:The Ancestor's Tale|حكاية السلف (كتاب)|حكاية السلف}}Page 289,
- Chen, J. Y.; Huang, D. Y.; Li, C. W. (1999). "An early Cambrian craniate-like chordate". Nature. 402 (6761): 518. Bibcode:1999Natur.402..518C. doi:10.1038/990080.
- Shu, D. G.; Morris, S. C.; Han, J.; Zhang, Z. F.; Yasui, K.; Janvier, P.; Chen, L.; Zhang, X. L.; Liu, J. N.; Li, Y.; Liu, H. -Q. (Jan 2003), "Head and backbone of the Early Cambrian vertebrate Haikouichthys", Nature, 421, صفحات 526–529, Bibcode:2003Natur.421..526S, doi:10.1038/nature01264, ISSN 0028-0836, PMID 12556891
- "Shubin, Neil. (2008). Your Inner Fish. Pantheon. .
- Niedzwiedzki, G.; Szrek, P.; Narkiewicz, K.; Narkiewicz, M.; Ahlberg, P. (2010). "Tetrapod trackways from the early Middle Devonian period of Poland". Nature. 463 (7227): 43–48. Bibcode:2010Natur.463...43N. doi:10.1038/nature08623. PMID 20054388.
- Cota-Sánchez, J. Hugo & Bomfim-Patrício, Márcia C. (2010). "Seed morphology, polyploidy and the evolutionary history of the epiphytic cactus Rhipsalis baccifera (Cactaceae)" ( كتاب إلكتروني PDF ). Polibotanica. 29: 107–129. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 6 أكتوبر 201828 فبراير 2013
- Menkhorst, Peter; Knight, Frank (2001). A Field Guide to the Mammals of Australia. Oxford University Press. صفحة 14. .
- Michael Augee, Brett Gooden, and Anne Musser (2006). Echidna: Extraordinary egg-laying mammal. CSIRO Publishing.
- "Polar Bears/Habitat & Distribution". SeaWorld Parks & Entertainment. مؤرشف من الأصل في 25 أكتوبر 201321 فبراير 2011.
- "Sirenians of the World". Save the Manatee Club. مؤرشف من الأصل في 26 أكتوبر 201721 فبراير 2011.
- Continental Drift and Evolution. Biology.clc.uc.edu (2001-03-25). Retrieved on 2011-12-06. نسخة محفوظة 28 سبتمبر 2015 على موقع واي باك مشين.
- Augee, Michael; Gooden, Brett; Musser, Anne (2006). Echidna: Extraordinary egg-laying mammal. CSIRO Publishing. . مؤرشف من الأصل في 7 فبراير 2020.
- Murphy, James B.; Ciofi, Claudio; de la Panouse, Colomba; Walsh, Trooper, المحررون (2002). Komodo Dragons: Biology and Conservation (Zoo and Aquarium Biology and Conservation Series). واشنطن العاصمة: Smithsonian Books. .
- Burdick, Alan (2007-03-25). "The Wonder Land of Socotra, Yemen". ALAN BURDICK. مؤرشف من الأصل في 10 مايو 201208 يوليو 2010.
- "Tuatara". New Zealand Ecology: Living Fossils. TerraNature Trust. 2004. مؤرشف من الأصل في 8 أكتوبر 201810 نوفمبر 2006.
- "Facts about tuatara". Conservation: Native Species. Threatened Species Unit, Department of Conservation, Government of New Zealand. مؤرشف من الأصل في 30 سبتمبر 200710 فبراير 2007.
- Coyne, Jerry A. (2009). Why Evolution is True. Viking. صفحات 99–110. . مؤرشف من الأصل في 16 ديسمبر 2019.
- "New Caledonia's most wanted". مؤرشف من الأصل في 29 سبتمبر 201808 يوليو 2010.
- "Giant bushy-tailed cloud rat (Crateromys schadenbergi)". مؤرشف من الأصل في 30 يونيو 20108 يوليو 2010.
- Rabor, D.S. (1986). Guide to Philippine Flora and Fauna. Natural Resources Management Centre, Ministry of Natural Resources and University of the Philippines.
- Humphrey, Robert R. (1974). The Boojum and its Home. University of Arizona Press. .
- Schofield, James (27 July 2001). "Lake Baikal's Vanishing Nerpa Seal". موسكو تايمز. مؤرشف من الأصل في 1 مارس 200727 سبتمبر 2007.
- Baldwin, B. G.; Robichaux, R. H. (1995). "Historical biogeography and ecology of the Hawaiian silversword alliance (Asteraceae). New molecular phylogenetic perspectives". In Wagner, W. L.; Funk, V. A. (المحررون). Hawaiian biogeography: evolution on a hotspot archipelago. Washington: Smithsonian Institution Press. صفحات 259–287.
- Pallen, Mark (2009). Rough Guide to Evolution. Rough Guides. صفحة 87. .
- Irwin, D.E.; Irwin, J.H.; Price, T.D. (2001). "Ring species as bridges between microevolution and speciation" ( كتاب إلكتروني PDF ). Genetica. 112-113: 223–43. doi:10.1023/A:1013319217703. PMID 11838767. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 10 أبريل 2020.
- Davis, Paul and Kenrick, Paul. 2004. Fossil Plants. Smithsonian Books (in association with the Natural History Museum of London), Washington, D.C. (ردمك )
- Coyne, Jerry A. (2009). Why Evolution is True. Viking. صفحة 103. . مؤرشف من الأصل في 16 ديسمبر 2019.
- Pioneer Productions (2010-01-19). "Episode Guide". How The Earth Was Made. History channel. مؤرشف من الأصل في 29 سبتمبر 2015.
- Luo, Zhe-Xi; Ji, Qiang; Wible, John R.; Yuan, Chong-Xi (2003-12-12). "An early Cretaceous tribosphenic mammal and metatherian evolution". ساينس. 302 (5652): 1934–1940. Bibcode:2003Sci...302.1934L. doi:10.1126/science.1090718. PMID 14671295. مؤرشف من الأصل في 24 سبتمبر 201527 ديسمبر 2010.
- Nilsson, M. A.; Churakov, G.; Sommer, M.; Van Tran, N.; Zemann, A.; Brosius, J.; Schmitz, J. (2010-07-27). Penny, David (المحرر). "Tracking Marsupial Evolution Using Archaic Genomic Retroposon Insertions". PLoS Biology. 8 (7): e1000436. doi:10.1371/journal.pbio.1000436. PMC . PMID 20668664.
- Woodburne, Michael O.; Zinsmeister, William J. (Oct 1982). "Fossil Land Mammal from Antarctica". Science. 218 (4569): 284–286. Bibcode:1982Sci...218..284W. doi:10.1126/science.218.4569.284. PMID 17838631. مؤرشف من الأصل في 7 أبريل 200817 يناير 2009.
- Goin, Francisco J.; et al. (Dec 1999). "New Discoveries of "Opposum-Like" Marsupials from Antarctica (Seymour Island, Medial Eocene)". Journal of Mammalian Evolution. 6 (4): 335–365. doi:10.1023/A:1027357927460. مؤرشف من الأصل في 10 أبريل 202017 يناير 2009.
- Reguero, Marcelo A.; Marenssi, Sergio A.; Santillana, Sergio N. (May 2002). "Antarctic Peninsula and South America (Patagonia) Paleogene terrestrial faunas and environments: biogeographic relationships". Palaeogeography, Palaeoclimatology, Palaeoecology. 179 (3–4): 189–210. Bibcode:2002PPP...179..189R. doi:10.1016/S0031-0182(01)00417-5.
- Mills, William James. Exploring Polar Frontiers: A Historical Encyclopedia, ABC-CLIO, 2003. (ردمك ), (ردمك )
- Goin, F.J.; Reguero, M.A.; Pascual, R.; von Koenigswald, W.; Woodburne, M.O.; Case, J.A.; Marenssi, S.A.; Vieytes, C.; Vizcaíno, S.F. (2006). "First gondwanatherian mammal from Antarctica". Geological Society, London, Special Publications. 258 (1): 135–144. Bibcode:2006GSLSP.258..135G. doi:10.1144/GSL.SP.2006.258.01.10.
- Prothero, Donald R.; Schoch, Robert M. (2002). Horns, tusks, and flippers: the evolution of hoofed mammals. JHU press. صفحة 45. .
- James M. Sobel; et al. (2009), "The Biology of Speciation", Evolution, 64, صفحات 295–315, doi:10.1111/j.1558-5646.2009.00877.x, PMID 19891628
- Jerry A. Coyne; H. Allen Orr (2004), Speciation, Sinauer Associates, صفحة 5,
- Raven, Peter H. (2005). Biology of Plants (الطبعة 7th rev.). New York: W.H. Freeman. . OCLC 183148564.
- Haas, J. W. Jr. (June 2000). "The Rev. Dr. William H. Dallinger F.R.S.: Early Advocate of Theistic Evolution and Foe of Spontaneous Generation". Perspectives on Science and Christian Faith. 52: 107–117. مؤرشف من الأصل في 10 أبريل 202015 يونيو 2010. نسخة محفوظة 23 أكتوبر 2016 على موقع واي باك مشين.
- Le Page, Michael (16 April 2008). "NS:bacteria make major evolutionary shift in the lab". New Scientist. مؤرشف من الأصل في 17 سبتمبر 200809 يوليو 2012.
- Blount Z.D.; Borland C.Z.; Lenski, R.E. (June 2008). "Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli". Proceedings of the National Academy of Sciences of the United States of America. 105 (23): 7899–906. Bibcode:2008PNAS..105.7899B. doi:10.1073/pnas.0803151105. PMC . PMID 18524956.
- Lenski, Richard E.; Travisano, Michael (1994). "Dynamics of adaptation and diversification: A 10,000-generation experiment with bacterial populations". PNAS. 91 (15): 6808–6814. Bibcode:1994PNAS...91.6808L. doi:10.1073/pnas.91.15.6808. PMC . PMID 8041701.
- Turko, Patrick; et al. (2016), "Rapid Evolutionary Loss of Metal Resistance Revealed by Hatching Decades-Old Eggs", Evolution, 70, صفحات 398–407, doi:10.1111/evo.12859, PMID 26768308
- Tanaka, T.; Hashimoto, H. (1989). "Drug-resistance and its transferability of Shigella strains isolated in 1986 in Japan". Kansenshogaku Zasshi. 63 (1): 15–26. doi:10.11150/kansenshogakuzasshi1970.63.15. PMID 2501419.
- Thwaites, W.M. (Summer 1985). "New Proteins Without God's Help". Creation Evolution Journal. 5 (2): 1–3. مؤرشف من الأصل في 7 سبتمبر 2018.
- Evolution and Information: The Nylon Bug. Nmsr.org. Retrieved on 2011-12-06. نسخة محفوظة 15 فبراير 2019 على موقع واي باك مشين.
- Than, Ker (23 September 2005). "Why scientists dismiss 'intelligent design". إم إس إن بي سي. مؤرشف من الأصل في 23 أكتوبر 2012.
- Miller, Kenneth R. Only a Theory: Evolution and the Battle for America's Soul (2008) pp. 80–82
- Sean Stankowski; Matthew A. Streisfeld (2015), "Introgressive hybridization facilitates adaptive divergence in a recent radiation of monkeyflowers", Proc. R. Soc. B, 282, صفحة 20151666, doi:10.1098/rspb.2015.1666, PMC , PMID 26311673
- Science News, Dark Power: Pigment seems to put radiation to good use, Week of 26 May 2007; Vol. 171, No. 21, p. 325 by Davide Castelvecchi نسخة محفوظة 24 أبريل 2008 على موقع واي باك مشين.
- Dadachova, E.; Bryan, R.A.; Huang, X.; Moadel, T.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. (2007). Rutherford, Julian (المحرر). "Ionizing Radiation Changes the Electronic Properties of Melanin and Enhances the Growth of Melanized Fungi". PLOS ONE. 2 (5): e457. Bibcode:2007PLoSO...2..457D. doi:10.1371/journal.pone.0000457. PMC . PMID 17520016.
- John A. Endler (1980). "Natural Selection on Color Patterns in Poecilia reticulata". Evolution. 34 (1): 76–91. doi:10.2307/2408316. JSTOR 2408316. PMID 28563214.
- David N. Reznick; Frank H. Shaw; F. Helen Rodd; Ruth G. Shaw (1997). "Evaluation of the Rate of Evolution in Natural Populations of Guppies (Poecilia reticulata)". Science. 275 (5308): 1934–1937. doi:10.1126/science.275.5308.1934. PMID 9072971.
- David Reznick; John A. Endler (1982). "The Impact of Predation on Life History Evolution in Trinidadian Guppies (Poecilia reticulata)". Evolution. 36 (1): 160–177. doi:10.2307/2407978. JSTOR 2407978. PMID 28581096.
- Medical Research Council (UK) (21 November 2009). "Brain Disease 'Resistance Gene' Evolves in Papua New Guinea Community; Could Offer Insights Into CJD". Science Daily (online). Science News. مؤرشف من الأصل في 8 يونيو 201922 نوفمبر 2009.
- Mead, S.; Whitfield, J.; Poulter, M.; Shah, P.; Uphill, J.; Campbell, T.; Al-Dujaily, H.; Hummerich, H.; Beck, J.; Mein, C. A.; Verzilli, C.; Whittaker, J.; Alpers, M. P.; Collinge, J. (2009). "A Novel Protective Prion Protein Variant that Colocalizes with Kuru Exposure". The New England Journal of Medicine. 361 (21): 2056–2065. doi:10.1056/NEJMoa0809716. PMID 19923577.
- Byars, S. G.; Ewbank, D.; Govindaraju, D. R.; Stearns, S. C. (2009). "Natural selection in a contemporary human population". Proceedings of the National Academy of Sciences. 107 (suppl_1): 1787–1792. Bibcode:2010PNAS..107.1787B. doi:10.1073/pnas.0906199106. PMC . PMID 19858476.
- Soy and Lactose Intolerance Wayback: Soy Nutrition نسخة محفوظة 04 يوليو 2017 على موقع واي باك مشين.
- Enattah, N.S.; Sahi, T.; Savilahti, E.; Terwilliger, J.D.; Peltonen, L.; Järvelä, I. (2002). "Identification of a variant associated with adult-type hypolactasia". Nature Genetics. 30 (2): 233–7. doi:10.1038/ng826. PMID 11788828.
- Curry, Andrew (31 July 2013). "Archaeology: The milk revolution". Nature. مؤرشف من الأصل في 11 سبتمبر 2019.
- Harriet, Coles (2007-01-20). "The lactase gene in Africa: Do you take milk?". The Human Genome, Wellcome Trust. مؤرشف من الأصل في 29 سبتمبر 200818 يوليو 2008.
- Tishkoff, S.A.; Reed, F.A.; Ranciaro, A.; et al. (January 2007). "Convergent adaptation of human lactase persistence in Africa and Europe". Nature Genetics. 39 (1): 31–40. doi:10.1038/ng1946. PMC . PMID 17159977.
- Herrel, A.; Huyghe, K.; Vanhooydonck, B.; et al. (March 2008). "Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource". Proc. Natl. Acad. Sci. U.S.A. 105 (12): 4792–5. Bibcode:2008PNAS..105.4792H. doi:10.1073/pnas.0711998105. PMC . PMID 18344323.
- Bart Vervust; Irena Grbac; Raoul Van Damme (August 2007). "Differences in morphology, performance and behaviour between recently diverged populations of Podarcis sicula mirror differences in predation pressure". Oikos. 116 (8): 1343–1352. doi:10.1111/j.0030-1299.2007.15989.x.
- "Lizards Rapidly Evolve After Introduction to Island". National Geographic. مؤرشف من الأصل في 18 يونيو 2018.
- Myers, PZ (23 April 2008). "Still just a lizard". ScienceBlogs. مؤرشف من الأصل في 18 نوفمبر 2018.
- Welsh, Jennifer (17 February 2011). "Fish Evolved to Survive GE Toxins in Hudson River". لايف ساينس (موقع). مؤرشف من الأصل في 3 أكتوبر 201819 فبراير 2011.
- Isaac Wirgin; et al. (2011). "Mechanistic Basis of Resistance to PCBs in Atlantic Tomcod from the Hudson River". Science. 331 (6022): 1322–1325. Bibcode:2011Sci...331.1322W. doi:10.1126/science.1197296. PMC . PMID 21330491.
- Cheptou, P.; Carrue, O.; Rouifed, S.; Cantarel, A. (2008). "Rapid evolution of seed dispersal in an urban environment in the weed Crepis sancta". Proceedings of the National Academy of Sciences. 105 (10): 3796–9. Bibcode:2008PNAS..105.3796C. doi:10.1073/pnas.0708446105. PMC . PMID 18316722.
- "Evolution in the urban jungle". مؤرشف من الأصل في 08 نوفمبر 201208 يوليو 2010.
- "Human Activity Boosts Brain Size in Animals". Yale Scientific. مؤرشف من الأصل في 18 فبراير 201816 مارس 2015.
- Snell-Rood, Emilie C.; Wick, Naomi (2013). "Anthropogenic environments exert variable selection on cranial capacity in mammals" ( كتاب إلكتروني PDF ). R Soc B. 280 (1769): 20131384. doi:10.1098/rspb.2013.1384. PMC . PMID 23966638. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 9 أكتوبر 201816 مارس 2015.
- Rosenblum, Erica Bree (2006), "Convergent Evolution and Divergent Selection: Lizards at the White Sands Ecotone", The American Naturalist, 167, صفحات 1–15, doi:10.1086/498397, PMID 16475095
- Rosenblum, Erica Bree (2007), "A multilocus perspective on colonization accompanied by selection and gene flow", Evolution, 61, صفحات 2971–2985, doi:10.1111/j.1558-5646.2007.00251.x, PMID 17976180
- Des Roches, Simone (2014), "Beyond black and white: divergent behaviour and performance in three rapidly evolving lizard species at White Sands", Biological Journal of the Linnean Society, 111, صفحات 169–182, doi:10.1111/bij.12165
- Robertson, Jeanne Marie (2009), "Rapid divergence of social signal coloration across the White Sands ecotone for three lizard species under strong natural selection", Biological Journal of the Linnean Society, 98, صفحات 243–255, doi:10.1111/j.1095-8312.2009.01291.x
- Schemske, Douglas W. (2000), "Understanding the Origin of Species", Evolution, 54, صفحات 1069–1073, doi:10.1554/0014-3820(2000)054[1069:utoos]2.3.co;2
- Mallet, James (2001), "The Speciation Revolution", Journal of Evolutionary Biology, 14, صفحات 887–888, doi:10.1046/j.1420-9101.2001.00342.x
- Coyne, Jerry A.; Orr, H. Allen (2004), Speciation, Sinauer Associates, صفحة 37,
- Florin, A.-B.; A. Ödeen (2002), "Laboratory environments are not conducive for allopatric speciation", Journal of Evolutionary Biology, 15, صفحات 10–19, doi:10.1046/j.1420-9101.2002.00356.x
- Coyne, Jerry A.; Orr, H. Allen (2004), Speciation, Sinauer Associates, صفحات 88–89,
- Liebherr, James K.; McHugh, Joseph V. in Resh, V. H.; Cardé, R. T. (Editors) 2003. Encyclopedia of Insects. Academic Press.
- Benton, Michael J.; Pearson, Paul N. (2001), "Speciation in the Fossil Record", Trends in Ecology and Evolution, 16, صفحات 405–411, doi:10.1016/S0169-5347(01)02149-8
- Lazarus, David; et al. (1995), "Sympatric Speciation and Phyletic Change in Globorotalia truncatulinoides", Paleobiology, 21, صفحات 28–51, doi:10.1017/s0094837300013063
- Kellogg, Davida E.; Hays, James D. (1975), "Microevolutionary Patterns in Late Cenozoic Radiolaria", Paleobiology, 1, صفحات 150–160, doi:10.1017/s0094837300002347
- Hays, James D. (1970), "Stratigraphy and Evolutionary Trends of Radiolaria in North Pacific Deep-Sea Sediments", Geological Society of America Memoirs, 126, صفحات 185–218, doi:10.1130/MEM126-p185,
- Ulf Sörhannus; et al. (1998), "Cladogenetic and anagenetic changes in the morphology of Rhizosolenia praebergonii Mukhina", Historical Biology, 1, صفحات 185–205, doi:10.1080/08912968809386474
- Sörhannus, Ulf; et al. (1991), "Iterative evolution in the diatom genus Rhizosolenia Ehrenberg", Lethaia, 24, صفحات 39–44, doi:10.1111/j.1502-3931.1991.tb01178.x
- Pearson, Paul N.; Ezard, Thomas H. G. (2014). "Evolution and speciation in the Eocene planktonic foraminifer Turborotalia". Paleobiology. 40: 130–143. doi:10.1666/13004.
- Gingerich, P. D. (1985), "Species in the Fossil Record: Concepts, Trends, and Transitions", Paleobiology, 11, صفحات 27–41, doi:10.1017/s0094837300011374
- Gingerich, P. D. (1976), "Paleontology and Phylogeny: Patterns of Evolution at the Species Level in Early Tertiary Mammals", American Journal of Science, 276, صفحات 1–28, Bibcode:1976AmJS..276....1G, doi:10.2475/ajs.276.1.1
- Chaline, J.; et al. (1993), "Morphological Trends and Rates of Evolution in Arvicolids (Arvicolidae, Rodentia): Towards a Punctuated Equilibria/Disequilibria Model", Quaternary International, 19, صفحات 27–39, Bibcode:1993QuInt..19...27C, doi:10.1016/1040-6182(93)90019-c
- Rice, William R.; Salt, George W. (1990). "The Evolution of Reproductive Isolation as a Correlated Character Under Sympatric Conditions: Experimental Evidence". Evolution, Society for the Study of Evolution. 44.
- "The Evolution of Reproductive Isolation as a Correlated Character Under Sympatric Conditions: Experimental Evidence" ( كتاب إلكتروني PDF ). William R. Rice, George W. Salt. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 13 مايو 201223 مايو 2010.
- "Observed Instances of Speciation, 5.3.5 Sympatric Speciation in Drosophila melanogaster". Joseph Boxhorn. مؤرشف من الأصل في 16 يونيو 201923 مايو 2010.
- Scott P. Egan; et al. (2012), "Divergent host-plant use promotes reproductive isolation among cynipid gall wasp populations", Biology Letters, 8, صفحات 605–608, doi:10.1098/rsbl.2011.1205, PMC , PMID 22337505
- Egan, Scott P.; et al. (2013), "Parallel Patterns of Morphological and Behavioral Variation among Host-Associated Populations of Two Gall Wasp Species", PLOS ONE, 8, صفحات e54690, Bibcode:2013PLoSO...854690E, doi:10.1371/journal.pone.0054690, PMC , PMID 23349952
- Feder, J.L.; Roethele, J.B.; Filchak, K.; Niedbalski, J.; Romero-Severson, J. (1 March 2003). "Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella". Genetics. 163 (3): 939–53. PMC . PMID 12663534.
- Berlocher, S.H.; Bush, G.L. (1982). "An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny". Systematic Zoology. 31 (2): 136–55. doi:10.2307/2413033. JSTOR 2413033.
- Berlocher, S.H.; Feder, J.L. (2002). "Sympatric speciation in phytophagous insects: moving beyond controversy?". Annu Rev Entomol. 47: 773–815. doi:10.1146/annurev.ento.47.091201.145312. PMID 11729091.
- Bush, G.L. (1969). "Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera: Tephritidae)". Evolution. 23 (2): 237–51. doi:10.2307/2406788. JSTOR 2406788.
- Prokopy, R.J.; Diehl, S.R.; Cooley, S.S. (1988). "Behavioral evidence for host races in Rhagoletis pomonella flies". Oecologia. 76 (1): 138–47. Bibcode:1988Oecol..76..138P. doi:10.1007/BF00379612. JSTOR 4218647. PMID 28312391. مؤرشف من الأصل في 10 أبريل 2020.
- Feder, J.L.; Roethele, J.B.; Wlazlo, B.; Berlocher, S.H. (1997). "Selective maintenance of allozyme differences among sympatric host races of the apple maggot fly". Proc Natl Acad Sci USA. 94 (21): 11417–21. Bibcode:1997PNAS...9411417F. doi:10.1073/pnas.94.21.11417. PMC . PMID 11038585.
- Fonseca, D. M.; Keyghobadi, N; Malcolm, CA; Mehmet, C; Schaffner, F; Mogi, M; Fleischer, RC; Wilkerson, RC (2004). "Emerging vectors in the Culex pipiens complex" ( كتاب إلكتروني PDF ). Science. 303 (5663): 1535–8. Bibcode:2004Sci...303.1535F. doi:10.1126/science.1094247. PMID 15001783. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 23 يوليو 2011.
- Burdick, Alan (2001). "Insect From the Underground — London, England Underground home to different species of mosquitos". Natural History. مؤرشف من الأصل في 10 أبريل 2020. نسخة محفوظة 6 سبتمبر 2015 على موقع واي باك مشين.
- "London underground source of new insect forms". The Times. 1998-08-26. مؤرشف من الأصل في 3 يناير 2019.
- Byrne, K.; Nichols, R.A. (1999). "Culex pipiens in London Underground tunnels: differentiation between surface and subterranean populations". Heredity. 82 (1): 7–15. doi:10.1038/sj.hdy.6884120. PMID 10200079.
- Vinogradova, E.B.; Shaikevich, E.V. (2007). "Morphometric, physiological and molecular characteristics of underground populations of the urban mosquito Culex pipiens Linnaeus f. molestus Forskål (Diptera: Culicidae) from several areas of Russia" ( كتاب إلكتروني PDF ). European Mosquito Bulletin. 22: 17–24. مؤرشف من الأصل ( كتاب إلكتروني PDF ) في 22 أبريل 2016.
- Hurt, Carla; et al. (2008), "A Multilocus Test of Simultaneous Divergence Across the Isthmus of Panama Using Snapping Shrimp in the Genus Alpheus", Evolution, 63, صفحات 514–530, doi:10.1111/j.1558-5646.2008.00566.x, PMID 19154357
- C. Montes; et al. (2015), "Middle Miocene closure of the Central American Seaway", Science, 348, صفحات 226–229, Bibcode:2015Sci...348..226M, doi:10.1126/science.aaa2815, PMID 25859042
- Knowlton, Nancy (1993), "Divergence in Proteins, Mitochondrial DNA, and Reproductive Compatibility Across the Isthmus of Panama", Science, 260, صفحات 1629–1632, Bibcode:1993Sci...260.1629K, doi:10.1126/science.8503007
- Knowlton, Nancy; Weigt, Lee A. (1998), "New dates and new rates for divergence across the Isthmus of Panama", Proc. R. Soc. Lond. B, 265, صفحات 2257–2263, doi:10.1098/rspb.1998.0568, PMC
- Rieseberg, Loren H.; Wendel, Jonathan (2004), "Plant Speciation - Rise of the Poor Cousins", New Phytologist, 161, صفحات 1–21, doi:10.1111/j.1469-8137.2004.00957.x
- Lafon-Placette, Clément; et al. (2016), "Current plant speciation research: unravelling the processes and mechanisms behind the evolution of reproductive isolation barriers", New Phytologist, 209, صفحات 29–33, doi:10.1111/nph.13756, PMID 26625345
- Anacker, Brian L.; Strauss, Sharon Y. (2014), "The Geography and Ecology of Plant Speciation: Range Overlap and Niche Divergence in Sister Species", Proc. R. Soc. B, 281, صفحة 20132980, doi:10.1098/rspb.2013.2980, PMC , PMID 24452025
- Hegarty, Matthew J.; Hiscock, Simon J. (2004), "Hybrid speciation in plants: new insights from molecular studies", New Phytologist, 165, صفحات 411–423, doi:10.1111/j.1469-8137.2004.01253.x, PMID 15720652
- Wendel, Jonathan F. (January 2000). "Genome evolution in polyploids". Plant Molecular Biology. 42 (1): 225–249. doi:10.1023/A:1006392424384. ISSN 0167-4412. PMID 10688139.
- Sémon, Marie; Wolfe, Kenneth H. (December 2007). "Consequences of genome duplication". Current Opinion in Genetics & Development. 17 (6): 505–512. doi:10.1016/j.gde.2007.09.007. ISSN 0959-437X. PMID 18006297.
- Comai, Luca (November 2005). "The advantages and disadvantages of being polyploid". Nature Reviews Genetics. 6 (11): 836–846. doi:10.1038/nrg1711. ISSN 1471-0056. PMID 16304599.
- Mavarez, Jesús; Salazar, Camilo A.; Bermingham, Eldredge; et al. (15 June 2006). "Speciation by hybridization in Heliconius butterflies". نيتشر (مجلة). 441 (7095): 868–871. Bibcode:2006Natur.441..868M. doi:10.1038/nature04738. ISSN 0028-0836. PMID 16778888.
- Eric Baack; et al. (2015), "The Origins of Reproductive Isolation in Plants", New Phytologist, 207, صفحات 968–984, doi:10.1111/nph.13424, PMID 25944305
- Vallejo-Marín, Mario (2012). "Mimulus peregrinus (Phrymaceae): A new British allopolyploid species". PhytoKeys. 14 (14): 1–14. doi:10.3897/phytokeys.14.3305. PMC . PMID 23170069. مؤرشف من الأصل في 2 فبراير 201427 يناير 2014.
- Vallejo-Marín, Mario; Buggs, Richard J.; Cooley, Arielle M.; Puzey, Joshua R. (2015). "Speciation by genome duplication: Repeated origins and genomic composition of the recently formed allopolyploid species Mimulus peregrinus". Evolution. 69 (6): 1487–1500. doi:10.1111/evo.12678. PMC . PMID 25929999.
- Karpechenko, G.D. (1927). "Polyploid hybrids of Raphanus sativus X Brassica oleracea L". Bull. Appl. Bot. 17: 305–408.
- Terasawa, Y (1933). "Crossing between Brassico-raphanus and B. chinensis and Raphanus sativus". Japanese Journal of Genetics. 8 (4): 229–230. doi:10.1266/jjg.8.229.
- Lowe, Andrew J.; Abbott, Richard J. (1996). "Origins of the New Allopolyploid Species Senecio camrensis (asteracea) and its Relationship to the Canary Islands Endemic Senecio tenerifae". American Journal of Botany. 83 (10): 1365–1372. doi:10.2307/2446125. JSTOR 2446125.
- Coyne, Jerry A. (2009). Why Evolution is True. Penguin Group. صفحات 187–189. . مؤرشف من الأصل في 16 ديسمبر 2019.
- Missouri Botanical Garden. "TROPICOS Web display Senecio vulgaris L". Nomenclatural and Specimen Data Base. Missouri State Library. مؤرشف من الأصل في 10 أبريل 202001 فبراير 2008.
- Bomblies, Kirsten; Lempe, Janne; Epple, Petra; Warthmann, Norman; Lanz, Christa; Dangl, Jeffery L.; Weigel, Detlef (2007). "Autoimmune Response as a Mechanism for a Dobzhansky-Muller-Type Incompatibility Syndrome in Plants". PLoS Biol. 5 (9): e236. doi:10.1371/journal.pbio.0050236. PMC . PMID 17803357.
- "New plant species arise from conflicts between immune system genes". Ed Yong. مؤرشف من الأصل في 03 فبراير 201022 مايو 2010.
- Purves, William Kirkwood; Sadava, David E.; Orians, Gordon H.; Heller, H. Craig (2006). Life, the science of biology (الطبعة 7). Sinaur Associates, Inc. صفحة 487. .
- Soltis, Pam (2011-03-17). "UF researcher: Flowering plant study 'catches evolution in the act". EurekAlert, American Association for the Advancement of Science. مؤرشف من الأصل في 25 يناير 201828 مارس 2011.
- Buggs, Richard J.A.; Zhang, Linjing; Miles, Nicholas; Tate, Jennifer A.; Gao, Lu; Wei, Wu; Schnable, Patrick S.; Barbazuk, W. Brad; Soltis, Pamela S. (2011). "Transcriptomic Shock Generates Evolutionary Novelty in a Newly Formed, Natural Allopolyploid Plant". Current Biology. 21 (7): 551–6. doi:10.1016/j.cub.2011.02.016. PMID 21419627.
- Bearhop, S.; Fiedler, W.; Furness, R.W.; Votier, S.C.; Waldron, S.; Newton, J.; Bowen, G.J.; Berthold, P.; Farnsworth, K. (2005). "Assortative mating as a mechanism for rapid evolution of a migratory divide". Science. 310 (5747): 502–504. Bibcode:2005Sci...310..502B. doi:10.1126/science.1115661. PMID 16239479. Supporting Online Material - تصفح: نسخة محفوظة 13 مارس 2010 على موقع واي باك مشين.
- Yong, Ed (3 December 2009). "British birdfeeders split blackcaps into two genetically distinct groups". ScienceBlogs. مؤرشف من الأصل في 12 سبتمبر 201821 مايو 2010.
- Tobler, Michael (2009). "Does a predatory insect contribute to the divergence between cave- and surface-adapted fish populations?". Biology Letters. 5 (4): 506–509. doi:10.1098/rsbl.2009.0272. PMC . PMID 19443506.
- "Giant insect splits cavefish into distinct populations". إد جونغ. مؤرشف من الأصل في 01 فبراير 201022 مايو 2010.
- J. P. Doupé; J. H. England; M. Furze; D. Paetkau (2007), "Most Northerly Observation of a Grizzly Bear (Ursus arctos) in Canada: Photographic and DNA Evidence from Melville Island, Northwest Territories", Artic, 60, صفحات 271–276
- Gray, A. P. (1972), Mammalian hybrids. A check-list with bibliography (الطبعة الثانية), Farnham Royal, Slough SL2 3BN, England: Commonwealth Agricultural Bureaux
- Kutschera, Verena E; Bidon, Tobias; Hailer, Frank; Rodi, Julia L.; Fain, Steven R.; Janke, Axel (2014). "Bears in a Forest of Gene Trees: Phylogenetic Inference Is Complicated by Incomplete Lineage Sorting and Gene Flow". Molecular Biology and Evolution. 31 (8): 2004–2017. doi:10.1093/molbev/msu186. PMC . PMID 24903145.
- Lindqvist, C.; Schuster, S. C.; Sun, Y.; Talbot, S. L.; Qi, J.; Ratan, A.; Tomsho, L. P.; Kasson, L.; Zeyl, E.; Aars, J.; Miller, W.; Ingolfsson, O.; Bachmann, L.; Wiig, O. (2010). "Complete mitochondrial genome of a Pleistocene jawbone unveils the origin of polar bear". Proceedings of the National Academy of Sciences. 107 (11): 5053–7. Bibcode:2010PNAS..107.5053L. doi:10.1073/pnas.0914266107. PMC . PMID 20194737.
- Hailer, F.; Kutschera, V. E.; Hallstrom, B. M.; Klassert, D.; Fain, S. R.; Leonard, J. A.; Arnason, U.; Janke, A. (2012). "Nuclear Genomic Sequences Reveal that Polar Bears Are an Old and Distinct Bear Lineage". Science. 336 (6079): 344–7. Bibcode:2012Sci...336..344H. doi:10.1126/science.1216424. hdl:10261/58578. PMID 22517859.
- Liu S.; et al. (2014), "Population genomics reveal recent speciation and rapid evolutionary adaptation in polar bears", Cell, 157, صفحات 785–794, doi:10.1016/j.cell.2014.03.054, PMC , PMID 24813606
- Miller, W; Schuster, SC; Welch, AJ; Ratan, A; Bedoya-Reina, OC; Zhao, F; Kim, HL; Burhans, RC; Drautz, DI; Wittekindt, NE; Tomsho, LP; Ibarra-Laclette, E; Herrera-Estrella, L; Peacock, E; Farley, S; Sage, GK; Rode, K; Obbard, M; Montiel, R; Bachmann, L; Ingólfsson, O; Aars, J; Mailund, T; Wiig, O; Talbot, SL; Lindqvist, C (2012). "Polar and brown bear genomes reveal ancient admixture and demographic footprints of past climate change". Proceedings of the National Academy of Sciences of the United States of America. 109 (36): E2382–E2390. Bibcode:2012PNAS..109E2382M. doi:10.1073/pnas.1210506109. PMC . PMID 22826254.
- "Evolution". Polar Bears International. مؤرشف من الأصل في 30 نوفمبر 201630 يناير 2014.
- Hansman, Jared (6 August 2008). "Adaptive Traits of the Polar Bear (Ursus maritimus)". Helium, Inc. مؤرشف من الأصل في 25 يناير 201830 يناير 2014.
- Tickell, W. L. N. (March 2003). "White Plumage". Waterbirds: The International Journal of Waterbird Biology. 26 (1): 1–12. doi:10.1675/1524-4695(2003)026[0001:wp]2.0.co;2. JSTOR 1522461.
- Larson, Edward J. (2004). Evolution: The Remarkable History of a Scientific Theory. New York: Modern Library. صفحات 121–123, 152–157. . مؤرشف من الأصل في 16 ديسمبر 2019.
- Cott, Hugh B. (1940). Adaptive Coloration in Animals. Methuen. صفحات 68–72. مؤرشف من الأصل في 11 مايو 2017.
- Cook, L. M.; Grant, B. S.; Saccheri, I. J.; Mallet, James (2012). "Selective bird predation on the peppered moth: the last experiment of Michael Majerus". Biology Letters. 8 (4): 609–612. doi:10.1098/rsbl.2011.1136. PMC . PMID 22319093. مؤرشف من الأصل في 2 يونيو 2018.
- Wallace, Alfred Russel (2015) [1889]. Darwinism - An Exposition Of The Theory Of Natural Selection - With Some Of Its Applications. Read Books. صفحة 180. . مؤرشف من الأصل في 10 أبريل 2020.
- Poulton, Edward Bagnall (1890). The Colours of Animals. صفحة Foldout after page 339, and throughout.
- Simulated Evolution Gets Complex. Trnmag.com (2003-05-08). Retrieved on 2011-12-06. نسخة محفوظة 15 مارس 2019 على موقع واي باك مشين.
- Adami, C.; Ofria, C.; Collier, T.C. (2000). "Evolution of biological complexity". Proc Natl Acad Sci USA. 97 (9): 4463–8. arXiv:. Bibcode:2000PNAS...97.4463A. doi:10.1073/pnas.97.9.4463. PMC . PMID 10781045.
- Earl, D.J.; Deem, M.W. (2004). "Evolvability is a selectable trait". Proc Natl Acad Sci USA. 101 (32): 11531–6. arXiv:. Bibcode:2004PNAS..10111531E. doi:10.1073/pnas.0404656101. PMC . PMID 15289608.
- Stemmer, W.P. (1994). "DNA shuffling by random fragmentation and reassembly: in vitro recombination for molecular evolution". Proc Natl Acad Sci USA. 91 (22): 10747–51. Bibcode:1994PNAS...9110747S. doi:10.1073/pnas.91.22.10747. PMC . PMID 7938023.
- Sauter, E. (27 March 2006). "Accelerated Evolution" Converts RNA Enzyme to DNA Enzyme In Vitro". TSRI – News & Views. 6 (11). مؤرشف من الأصل في 21 يونيو 2017.
- Molecular evolution. kaist.ac.kr نسخة محفوظة 06 يوليو 2017 على موقع واي باك مشين.
- In Vitro Molecular Evolution. Isgec.org (1975-08-04). Retrieved on 2011-12-06. نسخة محفوظة 26 سبتمبر 2018 على موقع واي باك مشين.
{kind=link}
{kind=link}