L'origine de la vie sur Terre demeure incertaine. Les plus anciens micro-organismes fossiles observés sont datés d'au moins 3,5 Ga (milliards d'années) durant le Paléoarchéen. Une étude publiée en 2015 sur des inclusions de carbone considérées comme d'origine organique leur assigne un âge de 4,1 milliards d'années[1] et plus récemment des précipités hydrofuges sous-marins trouvés dans des roches sédimentaires ferrugineuses de la ceinture de roches vertes de Nuvvuagittuq (Québec) ont été interprétés comme de possibles traces de vie (organismes filamenteux) il y a au moins 3,77 Ga et peut-être 4,28 Ga[2]. Si leur origine biologique est confirmée, la vie pourrait donc être apparue dès l'Hadéen.
Cette question a donné naissance à une branche des sciences du vivant et de la cosmochimie, l'abiogenèse.


Apparition de la vie sur Terre
Histoire conceptuelle

Les premières interrogations sur l'origine des espèces sont bien antérieures à Darwin. Les religions traditionnelles font typiquement intervenir une divinité créatrice, responsable de cette création ex nihilo.
Fondant les sciences naturelles, Aristote et la philosophie grecque antique posent la théorie de la génération spontanée, qui a continué à être envisagée, y compris dans les milieux universitaires, jusqu'au XIXe siècle. D'après cette théorie, des animaux « inférieurs » qualifiés de « vermines » se forment spontanément à partir de matières en décomposition, comme les pucerons sur la rosée des plantes, les larves de mouches sur la viande, ou les souris dans du foin sale. Une théorie connexe et complémentaire était l'hétérogenèse, pour laquelle certaines formes de vie pouvaient émerger d'autres, comme les papillons des vers, ou les abeilles des fleurs. Avec la découverte des premiers micro-organismes en 1665 par Robert Hooke, beaucoup ont estimé que leur existence était une preuve à l'appui de la génération spontanée, car ils semblaient trop simplistes pour la reproduction sexuelle, et la reproduction asexuée par division cellulaire n'avait pas encore été observée.
Les travaux d'Antoni van Leeuwenhoek lui ont permis de contester ces idées courantes à l'époque, grâce à l'invention du microscope et à des expériences comme celles de l'incubation de viande scellée et ouverte par Redi, ou l'étude des bouillons de culture par Pasteur. Il a été progressivement démontré que, du moins dans le cas de tous les organismes supérieurs et facilement visibles, le sentiment antérieur concernant la génération spontanée était faux. Dans les années 1680, van Leeuwenhoek s'affirmait ainsi convaincu que la génération spontanée était incorrecte. L'hypothèse alternative était la biogenèse, selon laquelle chaque chose vivante provenait d'une chose vivante préexistante (« omne vivum ex ovo », « chaque chose vivante provient d'un œuf »).

Aujourd'hui, les scientifiques s'accordent à dire que toute la vie actuelle découle d'une vie antérieure, qui est devenue progressivement plus complexe et s'est diversifiée grâce au mécanisme d'évolution par sélection naturelle décrit par Charles Darwin. L'une des premières spéculations à ce sujet a été faite dans la lettre de Darwin à Joseph Dalton Hooker du [5], dans laquelle il suggère que « l'étincelle de vie d'origine peut avoir commencé dans une petite mare chaude, où étaient présents toutes sortes de produits de base, ammoniac et sels phosphoriques, la lumière, la chaleur, l'électricité, etc., et c'est là qu'un composé protéique s'est formé chimiquement, prêt à subir des changements encore plus complexes. ». Il poursuivait en expliquant que « à l'heure actuelle, une telle matière serait immédiatement dévorée ou absorbée, ce qui n'aurait pas été le cas avant la formation de créatures vivantes. ». Mais il avait lui-même conscience du caractère très conjectural, voire évanescent, de ces propositions, puisqu'il avait déjà écrit à Hooker en 1863 cette déclaration "définitive" : « Toutes ces spéculations sur l'origine de la vie sont bonnes à jeter au panier ; on pourrait tout autant spéculer sur l'origine de la matière. », ce qui ne l'empêchait manifestement pas de continuer à s'interroger (et cette interrogation sera reprise par de nombreux chercheurs jusqu'à nos jours, comme d'ailleurs celle sur l'origine de la matière). Toujours est-il que dans L'Origine des espèces, Darwin se contenta de dire que la vie avait été « créée [qu'elle était apparue] par un processus totalement inconnu », mais il avait rapidement regretté d'utiliser le terme « création », issu de l'Ancien Testament.
Les premières cellules

Qualitativement, un certain milieu disposant d'un gradient chimique a permis la création de composés organiques et de réactions variées. Certaines réactions conduisent à des polymérisations (polynucléotides), et certains composés formés agissent comme catalyseurs à d'autres. Dans un tel système complexe, dès qu'une chaîne de réactions s'auto-catalyse, elle tend à dominer et épuiser les ressources disponibles, et peut alors être à la base de nouvelles complexités.
Dans cette complexification croissante, il n'y a pas de limite franche entre un système autocatalytique et « la vie » ; dans les deux cas la compétition conduit à une sélection naturelle favorisant différentiellement ce qui est le plus efficace en matière de reproduction et d'utilisation efficiente de ressources. Dès que le système devient assez stable et robuste pour assurer une reproduction suffisamment fidèle, dans des conditions environnementales plus variées, il peut être qualifié d'autopoïétique : « la vie » commence et se répand dans tous les environnements connexes, en quelques centaines ou milliers d'années[6].
La voie exacte suivie par les origines de la vie, qui remonteraient à environ 3,5 à 3,8 milliards d'années, demeure incertaine, et la date d'apparition exacte de la première cellule n'est pas connue par les scientifiques. Mais d'une manière ou d'une autre, la vie a finalement pris la forme d'une « cellule », capable de maintenir l'intégrité d'un « individu » par rapport à l'environnement, capable de maintenir un métabolisme cohérent par des échanges chimiques avec le milieu environnant, et capable de se répliquer en produisant d'autres « individus » semblables.
Ces trois fonctions de base sont essentielles à la vie. La délimitation d'un « individu » par rapport à un environnement « extérieur » renvoie à ce qu'est une membrane plasmique, capable d'assurer une telle séparation. La réplication à l'identique d'un système formel renvoie au codage génétique de l'information nécessaire à cette cellule, probablement d'abord sous forme d'un monde à ARN, puis sous forme stabilisée par l'ADN. Mais c'est le métabolisme de ces cellules qui va s'avérer essentiel pour décrire leur évolution et leur influence sur l'histoire de la Terre, à travers l'impact que cette biochimie aura sur l'évolution de la planète.
Le registre fossile

Le premier intervalle de temps de l’histoire de la Terre est l’éon de l’Hadéen, il s’étend de la formation de la planète jusqu’au début de l’éon de l’Archéen, c’est-à-dire entre environ −4,568 et −4 Ga (milliards d'années).
Les fossiles connus les plus anciens sont des micro-organismes marins de type bactérien. Il s’agit essentiellement de stromatolithes constitués de cyanobactéries, qui forment des empilements de lamines que l’on retrouve ensuite sous forme de reliefs ou de dômes fossilisés dans les sédiments. Les plus anciens qui ne soient pas controversés ont été découverts en Australie et datent d'environ 3,5 Ga[7],[2],[1],[8]. On trouve aussi des fossiles bactériens bien conservés dans les chailles de la formation Strelley Pool (en) (Australie-Occidentale), qui datent de 3,4 Ga[9]. D'autres, âgés de 3,7 Ga, ont été décrits au Groenland[10] mais leur interprétation a été contestée en 2018, au profit d'une explication abiotique[11],[12].
D’autres micro-organismes (des tubes et des filaments micrométriques épigénisés en hématite) semblent s’être développés très tôt et avoir été fossilisés par des dépôts minéraux à proximité de sources hydrothermales sous-marines. En 2017, E. Bella et son équipe décrivent des fossiles de « micro-organismes putatifs » découverts dans des sédiments ferrugineux affleurant dans la ceinture de roches vertes de Nuvvuagittuq, au Québec[13],[2]. Ils sont datés d'au moins 3,77 Ga, peut-être même de 4,28 Ga[2].
Parallèlement à la recherche de microfossiles dans les sédiments les plus anciens de la Terre, l’analyse isotopique, en 2015, d’inclusions de carbone, sous forme de graphite, dans des zircons hadéens détritiques de la région de Jack Hills en Australie-Occidentale, a fourni un âge de 4,1 Ga[1].
Le même travail, fait à l'ouest du Groënland (dans les roches métasédimentaires d'Isua, qui datent d’il y a plus de 3,7 milliards d'années), a mis en évidence des composés carbonés pauvres en 13C présentant des rapports isotopiques compatibles avec une origine biogène[14],[15],[16]. Ces composés sont notamment trouvés en inclusion dans des cristaux de grenat métamorphique, dans un environnement de litage sédimentaire riche en carbone entièrement graphitisé. Jusqu'à mi-2017, les études n’avaient pas mis en évidence d'autres éléments permettant la vie, tels que de l'hydrogène, de l'oxygène, de l'azote et du phosphore, structurellement associés à cette matière carbonée[17],[18], puis une étude des inclusions trouvées dans les porphyroblastes de grenat a conclu que les spectres d'absorption de l’infrarouge de ces matériaux sont compatibles avec la présence de carbone lié à l'azote et à l'oxygène, et probablement aussi au phosphore, avec des niveaux de liaisons C-H ou O-H faibles[19]. Ceci plaide en faveur d’une matière organique biogénique restée isolée durant des milliards d'années, et métamorphisée à environ 500 °C, ce qui en ferait les plus anciennes traces de carbone biogénique connues en 2017 sur la planète, hypothèse soutenue dès 2014 par Ohtomo et al.[15] (confortée par la description de structures interprétées en 2016 comme des microorganismes fossiles[10] mais contestées en 2018[11],[12]).
Maturation des connaissances en biochimie

Le biochimiste Robert Shapiro a résumé les théories d'Oparine et Haldane sur la « soupe primordiale » de la manière suivante[20] :
- La Terre avait primitivement une atmosphère réductrice ;
- Cette atmosphère a produit des composés organiques simples (monomères) sous l'action de diverses sources d'énergie ;
- Ces composés se sont accumulés au fil du temps dans une sorte de soupe chimique, qui a pu se trouver concentrée en divers endroits (côte et lagune, mont hydrothermal, etc.) ;
- Par la suite, des polymères et composés plus complexes se sont développés dans le mélange, conduisant finalement à la vie.
Les trois premières étapes ont fait l'objet de reconstitutions et d'études en laboratoire ; mais la quatrième « étape » de cette théorie a été qualifiée de très simpliste - rappelant le coup de baguette magique des contes de fées.
Plusieurs modèles font l'hypothèse de l'apparition d'un métabolisme primitif qui aurait précédé l'émergence de la réplication de l'ARN. Une des premières versions de cette hypothèse fut présentée en 1924 par Alexandre Oparine avec son idée de vésicules primitives capables de se répliquer (coacervats), à une époque où on ne connaissait pas encore la structure de l'ADN.
L’école de pensée privilégiant le rôle du métabolisme[21],[22], comme le concept d’autopoïèse[23], s’inspire du concept du système dissipatif qui se maintient loin de l'équilibre thermodynamique, grâce au flux de matière et d'énergie qui le traverse[24].
Le problème de cette approche est de ne pouvoir rendre compte de l'évolution de ces systèmes métaboliques[25], car ils se répliquent sans transmettre d’information, c’est-à-dire qu’ils n'ont pas d'hérédité[26].
Les modèles « gènes d'abord » ont été formulés après la découverte de l'ADN et de son rôle dans l'hérédité. Dans ces modèles, l'apparition du génome a précédé l'apparition du métabolisme. Des molécules d'ADN auraient ainsi existé seules, s'autorépliquant à partir des molécules présentes dans leur environnement. Les « individus » sont donc représentés par les molécules d'acides nucléiques elles-mêmes.
S'il est logique de penser que des polymères non homogènes aient pu faire l'objet d'une pression de sélection, la nature exacte de ce polymère est cependant une question ouverte quand on se réfère à la biochimie actuelle. En effet, la biochimie actuelle sépare la fonction de catalyseur, tenue par les polypeptides (polymères d'acide aminé), et celle de support et de réplication de l'information, tenue par l'ADN. Ces deux aspects sont inter-dépendants, mais actuellement mutuellement exclusifs. L’origine de la vie soulève alors une difficulté logique, celle de l’antériorité du métabolisme ou du code génétique. En effet, un métabolisme complexe ne peut pas exister sans protéines, donc sans code génétique permettant de les reproduire ; mais inversement le code génétique n’a pas de sens sans le milieu métabolique capable de l’interpréter.
La logique d'une approche de type abiogenèse suppose l'existence d'un polymère à la fois auto-répliquant et ayant des fonctions de catalyseur. L’école de pensée privilégiant les acides nucléiques se subdivise en deux classes d’hypothèses[27] :
- la première est que les premières formes de vie sur Terre ont utilisé l’ARN comme seul constituant génétique capable d’encoder l’information nécessaire aux catalyses biologiques ;
- la deuxième est encore plus restrictive puisqu’elle considère l’ARN comme la première forme de vie sur Terre, c’est-à-dire comme le premier système chimique à l’origine de l’évolution biologique, l'hypothèse du monde à ARN.
Il y a ensuite un gouffre à franchir entre les modèles décrivant la complexification croissante d'une chimie pré-biotique pour aboutir à la formation de quelques polynucléotides dominants, et ceux modélisant la cellule primitive moderne à ADN, qui semble nécessiter au bas mot quelques centaines de gènes pour produire les constituants d'un métabolisme capable de se répliquer[28]. Seules les protéines-enzymes y paraissent capables d'une activité catalytique, mais ces protéines sont trop complexes et variées pour être dupliquées.
Matériaux et méthodes

Si la théorie de l'évolution a pu reposer sur l'étude des restes fossiles, elle ne permet pas par cette méthode de remonter dans ce qui s'est passé pendant le précambrien. Les analyses isotopiques modernes réalisées sur des roches plus anciennes ont permis de retracer les grandes évolutions précambriennes dans l'Histoire de la Terre, et en particulier l'oxygénation de la Terre sous l'influence de la biosphère, mais sans pouvoir rentrer dans le détail de l'évolution qui a permis de passer de réactions chimiques élémentaires à l'émergence de la première cellule.
La biochimie a pu donner un premier éclairage sur les différents systèmes métaboliques, montrant en particulier la différence de nature des membranes cellulaires des archées par rapport à celles des bactéries, et les isolant comme un règne à part entière issu d'un dernier ancêtre commun universel (LUCA en anglais) primitif.
C'est la découverte du séquençage de l'ADN, puis à partir de 2005 des méthodes de séquençage haut débit, qui a permis d'analyser de manière pratique le génome d'organismes variés, et d'en décrire les variations par cladistique, reconstituant ainsi un arbre supposé représenter la classification phylogénétique de ces espèces. En parallèle, le degré de divergence de ces séquences permet d'en estimer la date de divergence par des méthodes d'horloge moléculaire. En pratique ces méthodes restent d'interprétation délicate, du fait de la présence de transfert horizontal de gènes venant emmêler les branches de l'arbre.
Au-delà de la comparaison des séquences génétiques, la comparaison structurale des domaines des protéines ou de l'ARN du ribosome permet de retrouver des similitudes entre organismes là où la dérive génétique ne permet plus de comparaison au niveau du code génétique lui-même.
Penser l’émergence de la complexité
Métabolisme et génome

Le nom de Manfred Eigen est lié à la théorie des hypercycles (en), la liaison cyclique de cycles de réactions comme explication de l'auto-organisation des systèmes prébiotiques, qu'il a décrit dans les années 1970.
Eigen souligna le problème que représente l'accumulation explosive d'erreurs si la recopie n'est pas fidèle : pour qu'une molécule de longueur L puisse s'imposer par rapport à ses allèles, il faut que le taux d'erreur par nucléotide soit inversement proportionnel à la longueur L[29]. Cette contrainte conduit au paradoxe d'Eigen (en) : sans système de correction d'erreur, la longueur maximale d'une molécule répliquante ne peut guère dépasser une centaine de bases ; mais pour qu'une molécule répliquante code une enzyme correctrice, sa longueur doit être significativement plus grande qu'une centaine de bases : comment une molécule répliquante capable d'engendrer un métabolisme complexe comprenant de nombreuses enzymes catalysantes a-t-il pu émerger ?
La solution proposée par Eigen est que le métabolisme initial a pu être un hyper-cycle ; chaque catalyseur élémentaire étant à la fois capable d'être répliqué dans l'environnement formé par les autres, et étant de longueur suffisamment faible pour rester sous le seuil catastrophique. L'ensemble peut alors déterminer un métabolisme complexe, sans que l'information nécessaire à sa définition soit concentrée sur un polymère unique[29]. Par ailleurs, ce système est stable si la reproduction d'un catalyseur n'est pas autonome, mais dépend de la production des autres[29].
Kauffman et les réseaux autocatalytiques
Que des formes complexes ne puissent pas avoir une origine aléatoire était un principe très généralement accepté. En 2002, Stuart Kauffman est le premier à l’avoir mis en doute à la suite d’expériences numériques, point de vue qu'il défendit dans l'introduction de son ouvrage Self Organization in Biological Systems. Il étudiait la dynamique de réseaux d’automates cellulaires engendrés d’une façon aléatoire. Il a alors observé que des formes stables animées d’un mouvement périodique apparaissaient spontanément, quelles que soient les conditions initiales. Il voyait ainsi des formes complexes et durables engendrées d’une façon purement aléatoire. Il a vite compris que son observation renouvelle d’une façon très originale nos façons d’expliquer les phénomènes et qu’il pouvait l’appliquer à la question de l’origine de la vie. Il a mis quelques années avant de le faire accepter par d’autres scientifiques.
Lorsque de nombreuses molécules différentes sont mises en présence, on peut décrire la dynamique chimique par un réseau. Chaque espèce de molécule est reliée aux autres espèces avec lesquelles elle réagit. Un tel réseau peut avoir des boucles autocatalytiques, c’est-à-dire qu’une espèce moléculaire, appelons-la A, favorise, ou catalyse, des réactions chimiques qui vont conduire à la production de molécules du type A. Une telle boucle est potentiellement explosive, parce qu’il y a un effet boule de neige, mais si les espèces à partir desquelles A est produite sont en quantité limitée, l’autoproduction de A est elle-même limitée.
Les êtres vivants sont toujours des réseaux autocatalytiques. Toutes leurs molécules (ADN, protéines et autres) réagissent avec les molécules ingérées (les aliments) pour produire des molécules semblables à elles-mêmes. Les plantes sont même capables d’utiliser la lumière du soleil dans ce but.
En étudiant des modèles, Kauffman a établi que sous des conditions assez générales un réseau de réactions chimiques contient nécessairement des réseaux autocatalytiques. Il suffit que le réseau soit suffisamment touffu, que les espèces moléculaires soient suffisamment nombreuses et réactives les unes vis-à-vis des autres.
Les réseaux autocatalytiques expliquent la reproduction des molécules mais à eux seuls ils ne suffisent pas pour expliquer la reproduction des unicellulaires.
Catalyse et métabolisme

La « soupe primordiale » est initialement formée par des molécules relativement simples, mais sa composition se complexifie progressivement à la suite des réactions chimiques portant sur les molécules de première génération. D'une part, l'action de la source d'énergie (rayonnement ultraviolet ou autre) sur ces premières molécules formées crée des radicaux libres plus complexes, capables d'interagir avec les molécules de premières générations pour conduire par générations successives à des molécules de plus en plus lourdes. D'autre part, les molécules ainsi formées peuvent directement réagir entre elles, conduisant à de nouvelles synthèses.
Les expériences visant à produire des composés dans des conditions prébiotiques conduisent généralement à des mélanges de très nombreuses molécules ; mais l'apparition de cycles catalytiques peut conduire à réduire cette diversité[30].
Un site métabolique peut être comparé à un récipient contenant un mélange de quelques milliers de composés chimiques[29]. Potentiellement, chaque composé A peut réagir avec n'importe quel autre B, conduisant potentiellement à des millions de réactions possibles A+B→C. Dans cette « soupe chimique », un composé A donné peut réagir avec différents composés B1, B2, etc., conduisant aux composés C1, C2, etc. Cependant ces réactions peuvent être plus ou moins rapides, suivant des ordres de grandeurs très différents. En pratique, ce qui consommera effectivement le composé A sera celle de ces réactions qui est la plus rapide, et pour laquelle le composant Bi est suffisamment présent.
Dans un premier temps, l'évolution de la composition ne dépend que de la constante d'équilibre et de la concentration molaire des paires de molécules susceptibles d'interagir. Cependant, dans un deuxième temps, certaines des molécules formées peuvent intervenir comme catalyseur sur telle ou telle réaction.
Un catalyseur se présente comme une espèce chimique intermédiaire K, qui peut fixer faiblement deux molécules particulières A et B (successivement, par exemple A+K→KA puis KA+B→KAB, ou dans l'ordre inverse). Une fois les deux composés A et B fixés, leur maintien dans leur position relative par le support K est tel que la vitesse de transition A+B→C peut être multipliée par un facteur allant jusqu'au million ou au milliard[29]. La réaction étant faite, son produit C se sépare du complexe KC→K+C, et le catalyseur K, qui n'a pas été altéré par ce cycle, peut capturer deux nouvelles molécules A et B et démarrer un nouveau cycle.
L'effet du catalyseur est tel, que les seules réactions observées en pratique sont celles pour lesquelles un catalyseur est présent, dès lors que cette possibilité existe. La présence dans la « soupe chimique » d'un catalyseur K catalysant la réaction A+B→C élimine pratiquement les alternatives, comme dans un procédé chimique industriel un réacteur K recevant deux tuyaux de A et de B pour produire le composé C. L'ensemble des catalyseurs présents dans cette « soupe chimique » va donc structurer les réactions qui s'y déroulent, suivant ce qui constitue un métabolisme déterminé.
À partir du moment où l'effet d'un catalyseur porte sur des réactions chimiques qui conduisent (directement ou non) à sa propre formation, une réaction en chaîne est susceptible de se mettre en place. Cette rétroaction chimique déplace l'équilibre global de la composition, augmentant progressivement la concentration du catalyseur, et tendant à assécher les réactifs consommés, qui ne peuvent alors plus alimenter d'autres types de synthèses.
Ainsi, avant même qu'un code génétique puisse se mettre en place, une pression de sélection a pu s'exercer sur la formation des molécules plus complexes, ou sur la formation de groupe fonctionnel au sein de ces molécules. Les réactions de synthèses organiques conduisant à la formation de catalyseurs favorables à ces mêmes synthèses bénéficient d'un avantage sélectif[31].
Sélection naturelle moléculaire
Pour Richard Dawkins[32], un système chimique « réplicateur » (ce qu'est, fondamentalement, un réseau autocatalytique) présente la propriété importante de ne pas être parfait : les répliques produites ne sont pas toujours strictement identiques au modèle original, et ces différences peuvent influer sur la capacité à se répliquer. Par ailleurs, les espèces chimiques de ce système peuvent avoir des durées de vie limitées et subir des transformations chimiques (des « mutations »), que ce soit par l'agressivité chimique du milieu où elles évoluent, par l'agitation thermique, ou par des rayonnements ionisants. Enfin, ces réplicateurs se « nourrissent » d'espèces chimiques de base, qui forment une ressource limitée dont le renouvellement ne peut qu'être lent en comparaison avec la croissance exponentielle d'une boucle autocatalytique.
La « soupe prébiotique » va donc progressivement se peupler de différentes variantes de réplicateurs, en compétition pour accéder aux ressources de base et réaliser le plus de duplications possibles dans une durée de vie limitée. Dans cet environnement, une variante devient d'autant plus majoritaire qu'elle présente des caractéristiques assurant un avantage comparatif par rapport aux autres variantes[32] :
- toutes choses égales par ailleurs, une espèce de molécule pourra contribuer à un nombre de répliques d'autant plus élevé que la durée de vie de cette molécule dans le milieu considéré est importante. Il y a donc une pression de sélection vers une longévité croissante de l'assemblage chimique formant le réplicateur ;
- toutes choses égales par ailleurs, un cycle chimique produira un nombre de répliques d'autant plus élevé qu'il peut les produire rapidement. Il y a donc une pression de sélection vers une « fécondité » croissante du réplicateur ;
- toutes choses égales par ailleurs, une variante deviendra d'autant plus majoritaire qu'elle assure une reproduction fidèle de son type. Il y a donc une pression de sélection vers une précision croissante de la duplication.
Que ce soit la longévité, la fécondité, ou la fidélité de la reproduction, dans tous les cas la pression de sélection favorise structurellement la meilleure performance dans l'une de ces trois formes de stabilité. À partir du moment où des boucles chimiques autocatalytiques se mettent en place, la « soupe prébiotique » est donc le lieu d'une sorte de sélection de survie conduisant à favoriser à terme toute nouvelle variante qui présente une meilleure stabilité, ou qui réduit la stabilité des variantes de réplicateurs avec lesquelles elle est en compétition[32].
Thermodynamique des systèmes hors équilibre
Entropie et déséquilibre


La vie n'est cependant pas juste une réaction chimique comme les autres, gouvernée en sens unique par le deuxième principe de la thermodynamique.
En tant que système organisé, tout élément vivant doit combattre la croissance interne de l'entropie afin de maintenir son propre état structuré. Cependant, une réduction d'entropie ne peut pas être réalisée par un système isolé. Ce maintien passe donc par la réalisation d'un couplage de ces processus internes à des processus externes augmentant l'entropie, de manière que le bilan d'ensemble aille dans le sens d'une création d'entropie.
Autrement dit, le sous-système qualifié de vivant doit pouvoir s'appuyer sur une source de faible entropie (matérialisée par une certaine quantité de matière ou d'énergie) et doit pouvoir la dégrader vers un puits de plus forte entropie. Fondamentalement, la vie ne peut exister qu'en exploitant un gradient d'entropie, c'est-à-dire un déséquilibre thermodynamique ; et c'est l'augmentation d'entropie que produit la dissipation de ce déséquilibre qui apparaît comme la « force motrice » de la vie[34].
Ce point fondamental, repris par Erwin Schrödinger dans son essai Qu'est-ce que la vie ?, avait déjà été souligné par Ludwig Boltzmann[35] :
« La lutte universelle des êtres animés pour exister n'est donc pas une recherche de matières premières - celles-ci, pour les organismes, sont l'air, l'eau et le sol, tous disponibles en abondance - ni d'énergie, qui existe en abondance dans tout corps sous forme de chaleur (bien que malheureusement pas transformable), mais la recherche d'entropie, qui est rendue disponible grâce au transfert de l'énergie du soleil chaud vers la terre froide. »
Il ressort de cette discussion que la diminution d'entropie qui caractérise une entité vivante ne peut pas correspondre à une simple réaction chimique se déroulant dans la « soupe primitive », mais passe nécessairement par l'action d'un médiateur « sur mesure » spécifique à un couplage particulier, agissant en présence d'un déséquilibre thermodynamique, médiateur dont le fonctionnement n'est pas seulement celui d'un catalyseur enzymatique, et doit en fait « transcender la chimie »[34].
Dispersion de déséquilibre
L'étude des systèmes hors équilibre[36] montre que les processus irréversibles présentent régulièrement deux aspects : près de l'équilibre, les flux détruisent l'information présente et créent de l'entropie ; mais loin de l'équilibre ils entraînent souvent l'apparition de structures dissipatives, localement créatrices d'ordre[37]. C'est par exemple ce qui fait la différence entre la conduction thermique et la convection.
À proximité de l'équilibre, les forces thermodynamiques induites par le flux d'énergie libre donnent lieu à des flux proportionnels à ces forces, qui conservent les symétries spatiale et temporelle de l'état d'équilibre, et se présentent comme des états dynamiques stables, c'est-à-dire que le système répond à des perturbations en les amortissant de manière exponentielle. Mais lorsque le déséquilibre augmente au point de saturer les capacités de dispersion, au-delà d'un certain point de bifurcation, le système répond aux fluctuations par une rupture de symétrie pour tomber dans un nouvel état stable présentant des flux et des structures organisés macroscopiquement. Ces structures dissipatives, états dynamiques organisés, sont intrinsèquement à faible entropie, instables et transitoires ; elles ne peuvent survivre qu'en étant constamment alimentées par le flux d'énergie libre qui les créé[37].
Sur le plan thermodynamique, l'émergence d'une structure qualifiée de vivante correspond à une telle structure dissipative, s'auto-organisant au sein d'un système présentant un déséquilibre thermodynamique élevé (gradient chimique, thermique...)[37].
Couplage et cycle de conversion

Un système vivant dépend d’une multitude de réactions endergoniques, thermodynamiquement « contre la pente », dont chacune doit nécessairement être entraînée en étant couplée à une réaction exergonique, entraînant un flux plus important « dans le sens de la pente ». Cependant, la réaction exergonique doit nécessairement être naturellement frustrée, et ne peut se réaliser qu'indirectement, par l'entremise d'un élément médiateur[38].
Dans cet échange, le processus de couplage est réalisé par un médiateur essentiel, dont l'intervention va au-delà de la simple chimie pour combiner au niveau moléculaire des effets à la fois thermodynamiques et mécaniques[34].
Les cycles de conversion apparaissant en biologie comprennent invariablement au moins six états distincts. On peut décrire un cycle minimaliste entre une réaction exergonique A⇒B et d'une réaction endergonique C⇒D par un médiateur M de la manière suivante[34] (voir figure) :
- Le médiateur M s'associe à un élément cible C, ce qui le met dans une configuration capable d'accepter ensuite un réactif A. La réaction C⇒D est endergonique, ce qui implique que soustraire l'élément C sous l'effet de fluctuations thermiques conduit localement à une diminution d'entropie.
- Le complexe MC s'associe à un réactif A, ce qui le met dans la configuration AMC. Le réactif A étant relativement fréquent, la transition de MC vers AMC peut être suffisamment rapide pour ne pas laisser au complexe MC le temps de relâcher sa cible C. Dans le complexe AMC, la cible C tend généralement à être stabilisée par la présence de A, empêchant la formation d'un complexe AM stérile.
- Le complexe AMC change de configuration stérique sous l'effet des chocs thermiques et des fluctuations énergétiques associées, et passe dans une configuration plus stable pouvant s'analyser comme un complexe BM'D. Contrairement à la forme précédente, c'est ici l'élément D qui tend à stabiliser la complexion de B.
- Toujours sous l'effet des chocs et fluctuations thermiques, l'élément D est accidentellement libéré, bien que l'injection de D conduise au contraire à diminuer localement l'entropie.
- L'élément B n'étant plus stabilisé par la présence de D est libéré du complexe M'B. Sur le complexe M'B, cette libération est en compétition avec la capture à nouveau d'un élément D reformant le complexe BM'D de départ. La libération de D intervient d'autant plus rapidement que la réaction A⇒B est exergonique, elle porte le médiateur dans un état non complexé M’ qui ne peut plus recevoir d'élément D. La dissipation d'énergie libre correspondant à la libération de B verrouille en pratique le cycle et interdit son retour en arrière.
- Sous sa forme décomplexée, le médiateur M’ reprend sa forme M, ce qui le rend capable d'initier un nouveau cycle.
Les points importants d'un tel cycle sont d'une part, que le relâchement de B, qui disperse l'entropie du cycle, est d'une manière ou d'une autre interdite tant que celui de D n'est pas acquis sous l'effet de fluctuations thermiques : la réaction A⇒B ne paye son enthalpie libre qu'après que la réaction C⇒D ait été exécutée. D'autre part, après la libération de D, il est souhaitable que cette réaction ait au contraire lieu le plus rapidement possible pour interdire en pratique tout retour en arrière[39]. Contrairement à ce que suggère une approche intuitive, dans un tel dispositif qui globalement convertit de l'enthalpie libre[38], ce n'est donc pas la réaction A⇒B qui fournit une énergie manquante à C⇒D : l'énergie nécessaire à C⇒D est simplement empruntée aux fluctuations thermiques du milieu, et la dissipation ultérieure d'énergie de A⇒B n'a pour fonction que de rendre cette transformation irréversible (et restituer l'emprunt, si bien que le bilan entropique sur l'ensemble du cycle est conforme au deuxième principe)[34].
La superposition de tels médiateurs en cascade forme en fin de compte un grand réseau dynamique, évolutif, auto-organisé[38]. La question de « l'origine de la vie » doit donc commencer par une exploration de ces médiateurs ; ce qui les a motivés, ce qu'ils ont produit, comment les moteurs de conversion auraient pu surgir et fonctionner de manière abiotique; et comment non seulement l'environnement physique a pu permettre que cela se produise[40], mais aussi a entretenu les fragiles étincelles de ce premier allumage au point qu'il a pu transmettre ce relais à l'autocatalyse[38] puis à la sélection naturelle, pour finalement exploser en cet immense système de conversion des déséquilibres organiques qui anime et impacte à présent la planète[34].
Chimie dans la soupe primitive
Problématique
La chimie prébiotique est une branche de la biochimie qui se propose d'étudier les conditions primitives de la vie sur Terre, et surtout la manière dont elle est apparue à partir de quelques molécules inertes.
La vie repose sur la chimie du carbone en présence d'eau, et met principalement en jeu cinq familles de composés chimiques :
- les lipides sont des molécules grasses comprenant de longues chaînes d'hydrocarbones. Elles jouent notamment un rôle important dans la structure des membranes cellulaires, conditionnant de manière active ou passive le passage d'autres molécules vers l'intérieur ou l'extérieur des cellules ;
- les « hydrates de carbone » sont des sucres. Différents « oses » monomères peuvent s'assembler en polymères appelés polysaccharides, comme la cellulose qui rigidifie les parois de la plupart des cellules végétales ;
- les bases azotées sont des molécules organiques dans lesquelles un groupe azoté amine joue un rôle important. La chlorophylle est par exemple construite sur un noyau de porphyrine constitué de quatre sous-unités amine, et joue un rôle essentiel pour capturer l'énergie solaire nécessaire à la vie végétale ;
- les monomères des acides nucléiques sont formés par un monosaccharide, une base azotée, et un ou plusieurs groupes phosphate ; les polymères correspondants matérialisent les gènes sous forme d'ADN ou d'ARN, et permettent la transcription de protéines ;
- une protéine est un polymère, généralement formé par l'assemblage de monomères pris parmi une vingtaine d'acides aminés, composés organiques comportant à la fois un groupe carboxyle (CO2H) et un groupe amine (NH2), et une chaîne hydrocarbone. Les protéines permettent la catalyse de nombreux processus chimiques indispensables à la vie.
Toute théorie de l'abiogenèse doit pouvoir expliquer l'origine et les interactions de ces cinq classes de molécules[41].
Conditions chimiques initiales

L’atmosphère de l’Archéen ne contient apparemment pas ou très peu d’oxygène libre.
Selon Alexandre Oparine et John Haldane, l'atmosphère terrestre primitive, lors de sa formation, était composée de méthane (CH4), d'ammoniac (NH3), de vapeur d'eau (H2O), de dioxyde de carbone (CO2) et de sulfure d’hydrogène (H2S). Par l'action du rayonnement ultraviolet provoquant la photolyse de ces molécules, l'atmosphère a évolué progressivement, perdant son méthane pour évoluer vers une atmosphère de gaz carbonique et d'azote. Lors de l'apparition de la vie, l'atmosphère est moyennement réductrice : (CO2 ; N2). Des modèles plus anciens considéraient une atmosphère fortement réductrice comme probable : (CH4 ; NH3).
L’eau sous forme liquide est présente, les océans ont probablement fini de se former durant l’Hadéen.
Au début de l'Archéen, l'activité solaire plus faible était compensée par une atmosphère très chargée en gaz à effet de serre. La température était élevée. L'eau et le gaz carbonique étant transférés dans les océans et les carbonates, l'atmosphère restante devient progressivement constituée majoritairement d'azote. L'effet de serre diminuant avec la perte du CO2, la température de surface baisse progressivement, et atteint des valeurs (70 à 100 °C) où les réactions chimiques des thermophiles deviennent possibles. L'analyse isotopique du silicium montre que la température des océans a décru de 70 °C il y a 3 500 Ma, à 20 °C il y a 800 Ma[42]. Si cette atmosphère s'était maintenue, la Terre ne serait plus habitable aujourd'hui.
La vie apparaît à cette époque.
L'expérience Urey-Miller et l'origine des molécules organiques
En 1953, le chimiste américain Stanley Miller réalisa une série d'expériences dans le cadre de sa thèse de doctorat.
Les expériences de Miller-Urey furent décisives dans la compréhension de la chimie prébiotique. En effet, à partir d'eau (H2O), de méthane (CH4), d'ammoniac (NH3) et d'hydrogène (H2), on a recueilli des acides aminés (dont 13 des 22 qui sont utilisés pour fabriquer des protéines dans les cellules des organismes), des sucres, des lipides, et quelques composants des acides nucléiques mais pas d'acides nucléiques entiers (ADN ou ARN). Des racémiques des énantiomères gauche lévogyre et droite dextrogyre se sont formés.
Les travaux révolutionnaires de Miller ont donné lieu à de très nombreuses publications, la composition de l'atmosphère terrestre primitive faisant débat.
En 2009, Sutherland et son équipe[43] sont parvenus à établir une voie de synthèse des nucléotides à base pyrimidique, l'acide uridylique et l'acide cytidylique. Les chercheurs butaient sur cette voie de synthèse prébiotique depuis quarante ans en raison de la difficulté à trouver la bonne façon de lier le ribose à la base azotée. La clé de cette voie est de passer par un précurseur commun au ribose et la base azotée. Ce précurseur, le 2-aminooxazol, est obtenu à partir de molécules organiques élémentaires : le glycéraldéhyde, le cyanamide, le cyanoacétaldéhyde, le cyanoacétylène et le phosphate inorganique. Le mélange réactionnel alimenté par de l'azote gazeux est soumis à un cycle de chauffage-refroidissement afin de simuler le cycle d'évaporation d'une mare par le rayonnement solaire et l'alimentation par la pluie. Après une semaine le 2-aminooxazol s'accumule dans le réacteur. Le précurseur se transforme ensuite en ribose et cytosine liés ensemble. Le phosphate est ensuite ajouté au milieu réactionnel en présence d'UV durant trois jours (l'absence d'ozone dans les conditions prébiotiques engendrait un bombardement intense d'UV). L'acide cytidylique se synthétise sous l'effet du rayonnement UV et quelques nucléotides portant une cytosine se transforment en acide uridylique. Pour les deux autres nucléotides, l'équipe de Sutherland travaillait sur un précurseur commun aux acides nucléiques à base purique.
Les conditions de la formation de molécules organiques se rencontrent également dans l'Atmosphère de Titan.
Des conditions prébiotiques plausibles entraînent la création de molécules organiques simples, qui sont les briques de base du vivant.
En 2016 deux équipes ont réussi à synthétiser les quatre bases de l'ARN (on pense que les premières formes de vie — y compris LUCA — étaient des cellules régulées par ARN) mais deux par deux (l'une les bases A et G, l'autre les bases C et U) et dans des conditions incompatibles l'une de l'autre. En 2019, une nouvelle équipe est parvenue à synthétiser les 4 bases dans des conditions représentatives de celles de la terre il y a 4 milliards d'années. Pour y parvenir, ils ont dû simuler le rythme des saisons en faisant varier l'humidité, les températures et l'acidité[44],[45].
Un autre aspect de la chimie prébiotique est la formation de peptides puis de protéines à partir des acides aminés, dont la synthèse abiotique est relativement aisée dans diverses conditions dont celles de l'expérience historique de Miller. Une voie abiotique possible est démontrée expérimentalement en 2023, dans une atmosphère gazeuse froide en présence de dimères moléculaires protonés de la glycine, sans substrat catalytique solide. Les résultats montrent la formation préférentielle de dimères protonés mixtes constitués d'un dipeptide et d'une molécule de glycine. En simulant l'impact de rayons cosmiques, on observe qu'une seule collision en phase gazeuse induit une polymérisation par déshydratation dans les ions dimères mixtes et purs. La croissance de la chaîne peptidique peut ainsi se produire au cours de réactions en phase gazeuse[46].
Structures cellulaires en émulsion

Les expériences sur la formation spontanée de membranes cellulaires sont bien antérieures aux synthèses de Urey-Miller de 1953.
Après de multiples réflexions et hypothèses, le biochimiste soviétique Alexandre Oparine est parvenu à créer de petites structures sphériques appelées coacervats à partir d'une émulsion contenant des protéines histones et de la gomme arabique, en 1922. Il s'agissait de sphérules colloïdales, de quelques micromètres de diamètre en suspension et liées entre elles. Les savons et les émulsions sont des substances de type colloïde.
Les coacervats d'Oparine ont la propriété d'être séparés du monde par une surface hydrophobe polyhémiacétalique (ils ont un intérieur et un extérieur) tout comme les cellules biologiques (mais dans le cas étudié, il ne s'agit pas de lipides). La similarité ne s'arrête pas là : en présence de catalyseurs, ces coacervats échangeaient des substances avec le monde extérieur, grossissaient et finissaient par se multiplier par scission.
C'est à partir de cette expérience que l'on imagina la synthèse possible d'un système vivant à partir de molécules organiques, d'eau et d'énergie provenant d'un rayonnement électromagnétique (Soleil, éclairs) : la « soupe prébiotique ».
Reprenant ces travaux, l'Anglais John Haldane proposa, en 1929, que ces systèmes « biologiques » ne pouvaient évoluer dans une atmosphère oxydante, en présence d'oxygène. En effet, la survie des molécules organiques (glucides, lipides, protéines, etc.) n'est possible qu'en l'absence de leur oxydation par le dioxygène.
L'apparition de vésicules dont la membrane serait formée de phospholipides pose de sérieux problèmes car ces lipides n'étaient pas présents sur la Terre primitive. Cependant des molécules amphiphiles plus simples ont pu être synthétisées de manière abiotique dans des conditions particulières. Par exemple il est reconnu que les sources hydrothermales ('mont hydrothermal') sont des environnements favorables pour des synthèses abiotiques soutenues[47]. À des températures entre 100 °C et 400 °C, des composés amphiphiles peuvent être produits par des réactions chimiques semblables au procédé Fischer-Tropsch[48]. De fait, la production abiotique de chaines courtes d'hydrocarbures a été récemment découverte au niveau du site hydrothermal Lost City situé au milieu de l'Atlantique[49]. Les concentrations millimolaires de méthane pourraient être à l'origine des réactions chimiques réductrices du carbone au niveau de ce type de site hydrothermal[50].
Autres milieux supports

L'assemblage de petites molécules (comme les acides aminés) en macromolécules (comme les protéines) nécessite l'élimination de molécules d'eau. Or, la thermodynamique indique qu'il est défavorable de réaliser une telle concentration dans l'eau elle-même. Il est possible pour résoudre cette contradiction de faire appel à des surfaces minérales, comme les micas, les argiles ou les pyrites. L'adsorption des petites molécules sur ces surfaces les concentre et les modifie chimiquement, ce qui peut rendre la formation de macromolécules plus favorable.
Le scénario de la « soupe primitive » comportant des micelles ayant des limites, les scientifiques commencèrent à repenser l'évolution de la vie prébiotique sans membrane, et imaginèrent des molécules organiques capables de croître sur une surface minérale. Les chimistes Graham Cairns-Smith et Günter Wächtershäuser (en), montrèrent que les argiles et les cristaux de pyrite permettent l'agglutination des molécules organiques facilement.
L'argile, par exemple, se trouve très abondamment sur Terre et est constituée d'un empilement de couches fines. Entre les différentes couches de l'argile peuvent se glisser certaines petites molécules organiques, ce qui permet une adsorption importante. L'argile est aussi un catalyseur très efficace pour de nombreuses réactions organiques et aurait donc pu permettre la polymérisation des acides aminés et/ou des acides nucléiques[51].
Une surface est plus stable qu'un espace tridimensionnel comme l'eau, mais aussi plus réduit (deux dimensions). Dans un espace tridimensionnel, les molécules sont sujettes à des contraintes selon tous les axes. Ainsi, à la surface des cristaux le milieu devient très réactif et facilite l'agglomération des molécules organiques.
D'autres variantes sont apparues dans les années 1980 et 1990, comme la théorie de Günter Wächtershäuser (en) sur un monde sulfuro-ferreux ou les modèles de Christian de Duve fondés sur la chimie des thioesters. Les thiols, et donc la cystéine, ont pu jouer un rôle majeur[52].
En 2022, la capacité des verres volcaniques de composition basaltique à catalyser la polymérisation des acides nucléiques est démontrée expérimentalement : les nucléotides s'assemblent spontanément en chaînes d'ARN, qui grandissent jusqu'à comporter quelques centaines de « lettres » au bout de quelques mois. La formation des nucléosides elle-même est catalysée par ces verres[53],[54].
Asymétrie des biomolécules

La chimie prébiotique conduit a priori à un racémique de molécules. Or, on sait depuis le milieu du XIXe siècle (notamment avec des travaux de Pasteur en 1847) que les acides aminés et les glucides simples naturels n'existent pratiquement que sous une de leurs deux formes énantiomères : la forme L pour les premiers et la forme D pour les seconds. On parle d'homochiralité du vivant. Si l'on suppose que cette asymétrie dans les composés monomères remonte à la formation de la « soupe originelle », se pose alors la question de l'origine de cette énantiospécificité originelle.
Cette asymétrie des molécules biologiques a été utilisée comme argument en faveur d'un dessein intelligent de l'origine de la vie.
Si l'on admet en revanche que l'énantiospécificité n'intervient qu'avec la formation de polymères, elle peut recevoir une explication plus simple. Les polypeptides sont plus stables quand ils sont formés par des acides aminés de même forme, qu'ils soient tous L ou tous D. Une chaîne composée d'un mélange des deux sortes d'isomères ne serait pas stable, parce que quand un acide aminé L est associé à un D, les deux chaînes latérales dépassent du même côté, conduisant à une tension structurelle qui affaiblit la liaison chimique[55].
Du moment que l'avantage sélectif appartient aux cycles qui produisent leurs propres catalyseurs, il suffit que le premier polypeptide impliqué dans un cycle favorable ait été composé de L pour que sa synthèse elle-même ait par la suite favorisé ce type de composé.
Machinerie pré-biotique
Site pré-biotique

Il y a des différences majeures entre les archées et les bactéries, en particulier pour ce qui est de la structure et de la formation des membranes cellulaires, et dans l'organisation et l'origine du métabolisme relatif à la réplication de l'ADN[56]. Ces différences suggèrent que à l'origine commune de ces types de cellules, ces deux métabolismes ne faisaient pas partie du répertoire figé, et donc que cette origine commune n'était pas encore une cellule au sens moderne du terme.
Dans tous les modèles modernes, la vie émergente ne peut prendre forme qu'initialement confinée dans des compartiments préexistants, proto-cellules ou surfaces favorables, qui servent de réacteurs et où les réactions et leurs produits peuvent être conservés concentrés[57],[58].
Sur la base de considérations énergétiques, la vie est assez probablement apparue autour d'une cheminée hydrothermale diffusant de l'hydrogène réducteur dans un milieu chargé en gaz carbonique[59],[60]. De tels sites présentent deux déséquilibres thermodynamiques importants[40] : d'une part, le gradient de potentiel rédox (< 1 V) entre H2 porté par les fluides internes et le CH4 de l'océan extérieur ; d'autre part, le gradient de protons correspondant à la différence de pH entre les fluides internes basiques (pH de l'ordre de 10,5) et un océan acide (pH de l'ordre de 5,5), entraînant une différence de potentiel électrique de l'ordre de 300 mV.
« D'un point de vue proto-biologique, nous pouvons voir tout le système convectif alcalin et le monticule hydrothermal initialement comme une seule entité proto-métabolisante complexe mesurant des kilomètres cubes ; où les conduits guidant l'océan carbonique vers le bas et à travers la croûte ont agi comme catalyseurs des importations redox, où l'eau était chauffée à environ 150°C et partiellement réduite en hydrogène, où le dioxyde de carbone était à son tour partiellement réduit en méthane et minoritairement en formiate, où les protons libéraient le calcium des silicates pour laisser une solution alcaline avec un pH ~ 11, où les ions bisulfure ont été libérés des sulfures métalliques, où les solutions exhalées ont réagi avec l'océan carbonique ambiant pour générer un monticule catalytique compartimenté, où les protons de cet océan se sont infiltrés dans le monticule pour stimuler la production de pyrophosphate et le dioxyde de carbone qui s'échappait pour être hydrogéné en formiate supplémentaire, où les oxydants à l'extérieur pourraient oxyder le méthane à l'intérieur en divers intermédiaires, et où tous les produits non coopératifs pourraient être épuisés avec l'effluent. Au fur et à mesure que le monticule évoluait, certains des chemins d'importation pourraient être entraînés vers sa base, commençant la tendance à la miniaturisation aux mètres cubes, et finalement impliquant les compartiments millimétriques à micrométriques comprenant les marges extérieures du monticule, un prélude au développement du premier biofilm quand les molécules organiques ont repris les principaux rôles structurels de la cellule tout en laissant les molécules inorganiques à leurs rôles catalytiques - phosphate pour le stockage et le transfert d'énergie libre, métaux de transition, en particulier sous forme de sulfures, dans la catalyse redox, et le magnésium comme influence stabilisatrice sur les biomolécules chargées négativement. »[40].
Membranes et protons
Quel que soit le réseau de réactions chimiques en présence, le mouvement d'ensemble doit être alimenté par une source d'énergie primaire[61].
L'énergie chimique potentielle résulte d'un gradient chimique, mais elle ne peut conduire à un résultat structuré que si elle est elle-même contrainte par une structure, en l'occurrence la membrane qui isole et localise les réactions initiales d'oxydoréduction. Le premier constituant de la chimie pré-biotique a donc été la formation d'une membrane entre quelque chose et autre chose, mais le lieu où se sont forgées les premières réactions auto-catalytiques n'a pas nécessairement pris la forme d'une membrane de lipides telle qu'on la connaît aujourd'hui.
À l'origine de la chimie pré-biotique, des fluides plutôt alcalins sortent d'un mont hydrothermal pour diffuser dans un océan rendu plutôt acide par la teneur en CO2 de l'atmosphère primitive. Ces fluides diffusaient à travers un labyrinthe de micro pores, éponge minérale sur laquelle des composés amphiphiles variés ont pu s'être assemblés en membranes. Quand ces membranes se plaçaient en travers du flux, la différence de pH entre l'intérieur du complexe hydrothermal et l'extérieur océanique était de l'ordre de 4, correspondant donc à une concentration naturelle de protons dix mille fois plus forte dans l'océan que dans le fluide hydrothermal[62]. C'est cette différence de polarité (extérieur positif) et de potentiel électrochimique (de l'ordre de 200 mV, que l'on constate dans les cellules modernes[62]), qui a pu constituer la « pile » primitive.
Le maintien d'un gradient de protons à travers une membrane est le mécanisme universel alimentant le métabolisme. Dans un premier temps, cette énergie a été fournie gratuitement par la géologie. Par la suite, le réseau métabolique a pu s'étendre à la réduction du gaz carbonique marin par l'hydrogène produit par la circulation hydrothermale, qui fonctionne ici comme une pile à combustible, et peut utiliser des sources d'énergie secondaires avec le catabolisme, qui est l'équivalent fonctionnel d'une cellule électrochimique. Plus tard, avec l'invention de la photosynthèse, l'énergie sera fournie par l'équivalent d'une cellule photo-électrique.
Source d'énergie
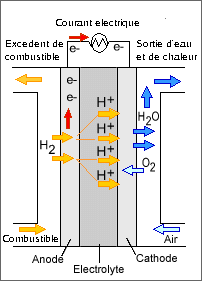
La source d'énergie primaire des cellules primitives aura été la méthanogenèse[63],[64], réaction d'oxydoréduction exothermique produisant du méthane à partir d'hydrogène et de gaz carbonique :
- CO2 + 4 H2 → CH4 + 2 H2O + 135 kJ
La réaction entre hydrogène et gaz carbonique est une réaction d'oxydoréduction entre le carbone et l'hydrogène, dont les demi-réactions correspondent à :
- Civ + 8 e− → C-iv (réduction du carbone)
- 8 H0 - 8 e− → 8 H+ (oxydation de l'hydrogène)
On peut noter la ressemblance entre cette réaction et le fonctionnement d'une pile à combustible à membrane d'échange de protons, le rôle de l'oxydant étant ici tenu par l'oxyde de carbone.
De fait, le métabolisme d'une cellule peut être vu d'une manière très générale comme la superposition de très nombreuses réaction d'oxydoréduction et d'échanges inter-régulées, séparés par des membranes ou par la localisation de différents sites de réactions. Les réactions sont alimentées par le flux créé par les gradients de concentration qui organisent à la fois les transferts des produits de réaction, et les transferts de donneurs ou d'accepteurs d'électron, qui peuvent eux-mêmes être des produits complexes (comme le nicotinamide adénine dinucléotide phosphate) ou simples (comme le proton H+). Ces mêmes éléments étaient nécessairement présents dès la chimie pré-biotique : un gradient de degré d'oxydation correspond à un système hors équilibre, source d'énergie, autour duquel se met en place un système dissipatif de plus en plus complexe[65].
La production de méthane à partir de gaz carbonique est exothermique, mais le CO2 est une molécule particulièrement stable, à laquelle il faut fournir de l'énergie par un intermédiaire pour la dissocier et initier la réaction. Une fois la réaction achevée, l'énergie récupérée représente de l'ordre de 150% de cette énergie d'activation[62]. Ce faible rendement interdit de réaliser la réaction par un cycle chimique direct, parce que l'énergie récupérée ne fournit que l'énergie nécessaire pour recharger un intermédiaire unique (de type ATP), pas deux, et en cas de cycle fermé il n'y a plus d'énergie excédentaire permettant d'alimenter ensuite d'autres réactions et de créer un métabolisme.
Le passage par un gradient de proton à travers une membrane permet de séparer l'activation et la récupération d'énergie. D'un côté, le gradient de proton présent sur la membrane permet d'entretenir la concentration d'intermédiaire chargés, capables de transporter de l'énergie dans le cytosol. De l'autre, ces intermédiaires permettent de fournir une énergie d'activation suffisante pour déclencher la dissociation du CO2 et entretenir le gradient de proton. L'énergie prélevée sur le gradient étant inférieure à celle fournie en retour, le gradient peut fournir des intermédiaires chargés en excès pour alimenter d'autres réactions[62].
Évolution chimique
Sélections de produits et voies chimiques

Sur un site pré-biotique, d'une manière ou d'une autre, les réactions chimiques du milieu ont conduit à la production régulière de monomères. Les chaînes polymérisées formées de monomères de même famille ont pu être synthétisées dans ce milieu. Les conditions d'émergence d'un tel site, et les voies chimiques qu'il pourrait abriter, ne sont pas identifiées[66].
Certaines des chaînes formées peuvent à leur tour intervenir comme catalyseur sur telle ou telle réaction de premier niveau. Ceci conduit à un second niveau de rétroaction, où des collections de courtes chaînes polymérisées servent de catalyseurs aux différentes réactions nécessaires pour la synthèse et l'assemblage de leurs constituants. Cette « pré-vie » est capable de sélection et de mutation[31], conduisant à une pression de sélection vers plus de pérennité, plus d'effet catalytique, et plus de rendement de réaction[32], mais n'est pas encore capable à ce niveau de stabiliser ni un métabolisme défini, ni une réplication fidèle.
À ce niveau, la sélection naturelle porte sur les voies des process chimiques concurrents sur un site pré-biotique donné. C'est à ce stade, en raison des voies réactives qui les forment et qu'ils permettent en retour, qu'a pu être sélectionnée la nature des monomères préférentiellement utilisés par les circuits métaboliques ultérieurs.
Compte tenu de leur apparition dans le métabolisme ultérieur, les composés favorisés ont compris les assemblages composés de base nucléique (ou base azotée), d'ose et de groupes phosphate, permettant la formation de nucléotides que l'on retrouve de manière universelle dans le système de transfert d'énergie (avec l'Adénosine triphosphate) et dans le code génétique (avec l'ARN).
Réplication de polymères
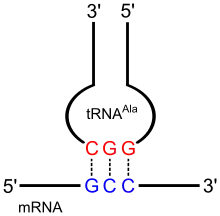
Après accumulation et concentration des monomères par la chimie pré-biotique, la « machine » la plus simple qui peut apparaître spontanément, et dont on peut supposer l’existence, est un mécanisme capable de polymériser ces composés pour former des polymères, homologues à des formes modernes : polypeptides, polysaccharides ou surtout polynucléotides[67].
Il s'agit initialement d'une polymérisation aléatoire, dans le sens où la voie de synthèse apparaissant peut assembler entre eux n'importe quels monomères ou polymère de la famille, indépendamment de leur nature. Les conditions du réacteur sont telles que des monomères sont produits et qu'ils subissent des polymérisations. Les polymères sont les produits d'accumulation dans ce milieu, où ils forment une population perpétuellement mouvante, en équilibre dynamique entre polymérisation et dégradation.
La chimie peut produire des gènes ou des protéines aléatoires, mais ne peut pas conduire directement à un métabolisme ; il faut une étape d’évolution intermédiaire[67]. Dans la population perpétuellement mouvante de chaînes polymères, certaines de ces chaînes polymérisées peuvent servir de modèles pour leur propre réplication[31], devenant ainsi leur propre catalyseur.
La polymérisation pertinente a probablement été celle de nucléotides d'ARN, initiant les premiers stades d'un monde à ARN. Si les conditions du site pré-biotique sont favorables à la polymérisation de l'ARN, celui-ci présente deux avantages clefs pour favoriser ensuite sa propre réplication : des brins d'ARN peuvent avoir une certaine activité catalytique, et plusieurs segments d'ARN peuvent s'apparier à un brin long, synthétisant le brin complémentaire d'un ARN double brin lorsqu'ils subissent leur polymérisation[68]. Dans ce contexte, la polymérisation n'est pas aléatoire, mais tend globalement à produire des chaînes reproduisant les séquences présentes dans la population existante, aux erreurs de retranscription et d'assemblage près.
Si l'on suppose que le milieu chimique est favorable et contient les constituants nécessaires, non seulement à la synthèse de ces polymères, mais également à leur réplication, les réactions chimiques de premier niveau, présentes dans la « soupe primordiale » initiale, conduisent alors, à un second niveau, à alimenter non seulement la formation, mais également la réplication de chaînes polymérisées.
Progénotes

Un point crucial pour discuter de la chimie pré-biotique est de voir que dans les cellules, les gènes et les protéines dans leur forme actuelle ne sont jamais produits par des réactions chimiques aveugles. Ils sont le résultat de voies métaboliques où des enzymes assemblent leurs composants dans un ordre imposé par un modèle : ce sont des artéfacts fruits de processus mis en œuvre par des machines, pas comme des molécules créées par des réactions chimiques[67].
De toute évidence, il est nécessaire de passer par une forme de sélection naturelle pour expliquer l'émergence progressive des métabolismes et des cellules modernes, mais tant que la complexité des métabolismes ne permet pas de créer des « cellules » vivantes, cette compétition ne peut pas prendre place entre des cellules — puisqu’elles n'existent pas. La sélection naturelle, à ce stade, a nécessairement porté sur des macro-molécules[56] portées par des sites pré-biotiques, et a été gouvernée par les principes généraux de cette sélection conduisant à une pression de sélection vers plus de longévité, plus de vitesse de reproduction, et plus de précision dans la réplication[32].
On peut visualiser cette évolution prébiotique sous la forme de transferts de « soupe prébiotique concentrée » encapsulée dans des vésicules amphiphiles et essaimant d'un site pré-biotique à l'autre[56]. En effet, un liposome se détachant du site pré-biotique emporte avec lui un échantillonnage suffisant des composés chimiques de son site pour reproduire les voies chimiques que ces composés supportent sur un site d'arrivée où il échoue, et orienter l'énergie et les matériaux de sa machinerie pré-biotique sur sa propre réplication, colonisant ainsi un nouveau site.
Carl Woese avait proposé en 1977 le terme « progénote » pour désigner un organisme vivant primitif cherchant à acquérir une maîtrise de son phénotype par son génome et qui serait l'ancêtre de l'ensemble du vivant[69],[70].
Hypothèse « lipidique »
En 2001, Doron Lancet publie l'hypothèse d'un « monde lipidique »[71]. Des molécules amphiphiles (pas nécessairement des phospholipides) présentent l'avantage de s'auto-assembler en micelles qui peuvent croître par capture de nouvelles molécules, et fissionner en micelles séparées quand elles atteignent une taille trop importante[72].
Les composés lipidiques (ou plus généralement amphiphiles[71]) dans une telle proto-membrane peuvent être extrêmement variés. Ces différents composants insérés dans la membrane sont libres de se déplacer sur sa surface, dans un monde bidimentionnel[72]. Leurs interactions entre eux et avec les autres molécules présentes dans le milieu peuvent ainsi forcer certaines configurations et avoir un rôle catalytique, plus facilement qu'en trois dimensions : certains composants (ou assemblages de composants) catalysent des réactions de synthèse et de fission. De tels catalyseurs peuvent être qualifiés de « lipozymes »[71]. Un tel milieu peut alors gagner en complexité et, à un certain point, voir émerger un réseau autocatalytique sur lequel il se condense : à partir du moment où un certain type de composition tend à se catalyser pour se reproduire à l'identique, les membranes de ce type croissent avec une composition sensiblement constante, produisant des micelles de compositions sensiblement identiques, et présentant les mêmes propriétés globales d'autocatalyse. Le tableau d'ensemble est celui d'un proto-métabolisme : des micelles individuelles croissent, fissionnent et se reproduisent à l'identique.
Ces proto-métabolismes ont pu avoir comme sous-produits parasite des ARN primitifs, récupérés ensuite pour former les bases d'une future information génétique.
Invention du code génétique
L'acide ribonucléique
Dans la biochimie actuelle, l'acide ribonucléique (ARN) joue un double rôle d'intermédiaire entre l'ADN support d'information et les polypeptides catalyseurs : l'ARN polymérase guide la polymérisation d'un brin ARN messager en suivant la composition d'une séquence de l'ADN, puis un ribosome guide la polymérisation d'un polypeptide en suivant la composition de cet ARN messager. Cette seconde traduction s'appuie sur une famille d'ARN de transfert, qui apportent l'acide aminé compatible avec la séquence de l'ARN messager en cours de traitement. En amont de la formation des protéines, les cellules vivantes contiennent une vingtaine d'ARN de transfert, chacune d'elles étant spécifique de l'un des acides aminés ; le ribosome assurant le décodage d'un ARN en assemblant les acides aminés dans l'ordre spécifié par l'ARN messager.
L'ARN est un polymère linéaire constitué d'un enchaînement de nucléotides. On trouve quatre bases nucléiques dans l'ARN : l'adénine, la guanine, la cytosine et l'uracile. L'ARN a de nombreuses similitudes avec l'ADN, avec cependant quelques différences importantes : d'un point de vue structurel, l'ARN est chimiquement moins stable ; sur le plan fonctionnel, l'ARN se trouve le plus souvent dans les cellules sous forme monocaténaire, c'est-à-dire de simple brin ; enfin, les molécules d'ARN présentes dans les cellules sont plus courtes, leur taille variant de quelques dizaines à quelques milliers de nucléotides.
La plupart des ARN naturels sont présents sous forme monocaténaire (simple brin) dans la cellule[73].
De même que les protéines, les brins d'ARN se replient le plus souvent sur eux-mêmes, formant une structure intramoléculaire qui peut être très stable et très compacte. La description des appariements internes entre les bases d'un ARN s'appelle la structure secondaire. La base de cette structure est la formation d'appariements internes, entre bases complémentaires : A avec U, G avec C (et, parfois, G avec U).
Un monde à ARN
L'hypothèse d'un monde à ARN, qui a aujourd'hui la faveur des scientifiques[74],[75], est fondée sur la découverte que l'Acide ribonucléique (ARN) peut à la fois être le support d'une information génétique, comme l'ADN, et assurer des tâches métaboliques, comme les protéines.
Le prix Nobel de chimie Thomas Cech indiqua que l'ARN pouvait être la première molécule répliquante du fait de ses propriétés catalytiques et auto-catalytiques :
- la structure des ARN est à la base de la richesse de leurs fonctions, et en particulier de leur capacité à catalyser des réactions chimiques (ribozymes) ;
- inversement, les règles d'appariement relativement rigides entre les bases de l'ARN permettent de transcrire un brin en son négatif, et par une nouvelle transcription permettent de dupliquer l'original.
Du point de vue de la reproduction, cette molécule possède donc deux fonctions primordiales : le stockage de l'information et la catalyse nécessaire à l'autoréplication. Il est donc en théorie possible, sur ce modèle, que l'ARN seul suffise à établir un métabolisme primitif.
L'image se dégageant de ces études est celle d'une population de brins d'ARN en interdépendance mutuelle, se reproduisant au sein d'une sorte d'écosystème chimique, et où chaque brin est en compétition sélective par rapport à ses propres allèles. C'est par exemple sa capacité optimale à se dupliquer rapidement dans un environnement peuplé d’enzymes ARN polymérase ARN-dépendante et de bases d'ADN, y compris sans amorce d'ARN initiale[76], qui caractérise le monstre de Spiegelman (en) : cette chaîne de 218 nucléotides d'ARN est un parasite prospérant dans un milieu contenant l'ARN polymérase et les bases d'ARN.
En 1990, Larry Gold et Jack Szostak ont mis au point une méthode visant à diriger l'évolution d'ARN, afin de sélectionner ceux montrant une activité catalytique. Ils ont depuis réussi à obtenir des ribozymes capables de lier des nucléotides entre eux, de lier des acides aminés à des ARN, d'effectuer des réactions d'oxydoréduction, de se lier à des composants de la membrane, etc.
C'est Walter Gilbert qui a utilisé pour la première fois le terme « monde à ARN » (en anglais : RNA world) en 1986. L'hypothèse du monde à acide ribonucléique (ARN) est que l'ARN était la principale — et sans doute la seule — forme de polymère supportant un métabolisme, avant l'émergence de la première cellule à ADN.
Toutefois, il reste encore à découvrir un ARN capable de se répliquer lui-même.
Hypothèse d'un ribosome autorépliquant
L'étape cruciale pour synthétiser des protéines et permettre l'apparition d'une première cellule semble être la formation d'un ribosome primitif, étant entendu qu'il a pu être par la suite soumis à la sélection naturelle, et évoluer en se complexifiant pour augmenter sa performance.
L'activité enzymatique de l'ARN ribosomique démontre que l'ARN peut avoir une activité enzymatique essentielle. L'acide ribonucléique ribosomique (ARNr) en est le principal constituant (60% en masse). Ainsi, le ribosome est un « ribozyme », dans le sens où le responsable de la synthèse des protéines n'est pas une protéine (comme c'est le cas dans la grande majorité des catalyses d'une cellule vivante) mais l'ARN ribosomique lui-même.
Le consensus scientifique est que l'ARN du ribosome est purement structurel et n'a pas vocation à être traduit[28] ; et de fait, son absence de traduction dans le monde à ADN lui permet de subir des dérives génétiques qui modifient le détail de son code sans avoir d'impact sur sa structure fonctionnelle. Cependant, l'analyse des séquences présentes dans la transcription des ARNr des procaryotes (23S, 16S et 5S) montre qu'elles présentent (encore?) de très fortes similarités avec celles des différents ARN de transfert[28]. De plus, la lecture des différents ARNr permet de synthétiser des protéines similaires à celles réalisant des fonctions importantes dans la transcription des ARN.
L'ensemble suggère que le ribosome a pu apparaître avant la première cellule, et a pu avoir eu à cette époque à la fois la capacité enzymatique de catalyser la polymérisation des protéines, et la capacité d'encoder les éléments (ARN-t et protéines) nécessaires à la duplication de ce métabolisme primitif, ce dont il conserve encore des traces sous la forme d'une grande densité d'informations redondantes[28].
À partir du moment où cette voie généraliste de synthèse des protéines est ouverte, la sélection naturelle ne porte plus uniquement sur la capacité enzymatique des brins d'ARN, mais également sur les protéines qu'ils sont capables de synthétiser.
Évolution de la traduction génétique
La traduction génétique est un mécanisme largement gouverné par l'ARN, qui a pu prendre naissance dans un monde à ARN[77]. Initialement, en l'absence de protéines sophistiquées, la traduction n'a pu être qu'imprécise, avec par exemple de mauvais appariements entre codons et anticodons, ou une mauvaise stabilisation du cadre de lecture. De ce fait, il n'était possible de synthétiser que des petites protéines, ou des protéines à séquence variable[77]. Mais inversement, l'absence de grande protéine permettait au génome d'être relativement petit.
Une sélection naturelle peut alors porter sur la performance de la traduction génétique associée à des macro-molécules d'ARN, et comme précédemment, induire une pression de sélection vers plus de longévité, plus de vitesse de reproduction, et plus de précision dans la réplication[32]. Une meilleure performance de la traduction génétique signifie que les protéines peuvent de leur côté évoluer vers plus de complexité et plus de spécificité[77], ouvrant la voie à de nouveaux mécanismes métaboliques.
Métabolisme élémentaire

Métabolismes élémentaires
La cellule a besoin d’énergie sous deux formes très différentes : celle de l'ATP, rapide et instable ; et celle restituée par la décomposition de composés (catabolisme) tels que les protéines, sucres et lipides, où l'énergie a été emmagasinée et stable, et pour lequel un accepteur d'électron est nécessaire[78]. D'autre part, les réactions d'oxydoréduction supposent la présence d'un donneur ou d'un accepteur d'électron. Enfin, sur le plan des composés organiques, la matière même de la cellule résulte de l'équilibre entre l'anabolisme, qui la construit en utilisant de l'énergie, et le catabolisme, qui en recycle les composés organiques et l'énergie.
- Pour produire de l'ATP, les premiers micro-organismes apparus vers −3,8 Ga consomment du gaz carbonique et de l'hydrogène, disponibles dans leur environnement, et rejettent du méthane[59],[60], dans une réaction redox fournissant directement de l'énergie utilisable sous forme d'ATP. Cette réaction utilise le dihydrogène comme donneur d'électrons principal :
- CO2 + 4 H2 → CH4 + 2 H2O + énergie.
- En aval de la production d'ATP, ces réactions de méthanogenèse produisent de l'énergie utilisable par la cellule, permettant en aval de produire des « hydrates de carbone » de formule générique CH2O, toujours à partir de gaz carbonique et d'eau, et initiant un cycle du carbone biologique :
- CO2 + 2 H2 → (CH2O) + H2O
- La fermentation présente l'avantage d'utiliser pratiquement les mêmes processus que ceux du métabolisme méthanogène précédent, et était donc à la portée de ces cellules primitives[60]. Pour la fermentation, le moins efficace des catabolismes, ce sont des composés organiques eux-mêmes qui jouent le rôle d'accepteur d'électron[78]. La fermentation de matière organique notée (CH2O) conduit à rejeter du méthane (et du gaz carbonique) dans une réaction qui peut se décrire schématiquement par[64] :
- 2 (CH2O) → CO2 + CH4
Transferts d'énergie
On constate aujourd'hui que l'adénosine triphosphate (ATP) sert de donneur d'énergie dans toutes les réactions chimiques nécessaires au métabolisme de tous les êtres vivants connus, se réduisant en adénosine diphosphate (ADP). De ce fait, il est légitime de considérer que ce « moteur » était également celui du dernier ancêtre commun universel. Plus en amont, il est probable que ce même moteur était déjà « inventé » avec la première cellule, et a peut-être même participé aux réactions autocatalytiques de la chimie prébiotique.
Par la suite, la variété du vivant s'est développée suivant deux directions :
- l'une, très prolifique en aval, a été de multiplier les composés organiques, les structures et les mouvements physiques qu'il est possible de produire dans une cellule à partir d'ATP — comment utiliser l'ATP ;
- l'autre direction, assez restreinte en amont, a été d'identifier des cycles biologiques capables de fournir cette énergie, et régénérer l'ADP consommé en ATP utilisable comme source d'énergie — comment régénérer l'ATP. L'histoire de la biosphère est jalonnée par l'ouverture de cycles énergétiques de plus en plus performants.
Premiers cycles écologiques
Transfert horizontal de gènes
Les transferts horizontaux modernes sont le fait de mécanismes très élaborés, mis en œuvre par des virus, ou réalisant des conjugaisons bactériennes échangeant des plasmides.
Les premiers transferts pré-biotiques ont pu prendre une forme beaucoup plus informelle de transferts mécaniques de « soupe pré-biotique concentrée » encapsulée dans des vésicules amphiphiles et essaimant d'un site pré-biotique à l'autre[56].
L'intégralité du métabolisme d'un site pré-biotique peut être ainsi dupliqué d'un site à l'autre. Cet essaimage permet de diversifier les métabolismes d'un site, et de coloniser de nouveaux sites prébiotiques, conduisant à une compétition entre variantes, et à une forme de sélection naturelle entre métabolismes de sites[77].
La prédominance des transferts horizontaux implique que l'évolution à ce stade ne fonctionne pas par apparition de nouvelles espèces dont on peut retracer l'arbre généalogique, mais fait évoluer en phase l'ensemble des sites concernés par ces échanges, en une évolution essentiellement monophylétique[79],[77]. Il faut que le métabolisme atteigne un seuil de complexité minimal, le « seuil darwinien », pour qu'une entité puisse « se » reproduire du seul fait de son métabolisme, et ainsi franchir l'étape critique à partir de laquelle le transfert de gènes vertical devient le mécanisme prépondérant[77].
Cependant cette forme non contrôlée laisse les sites pré-biotiques vulnérables à l'apparition de molécules prédatrices ou parasites, qui épuisent le métabolisme du site et sont susceptibles de se répandre par le même moyen.
Origine et évolution des capsides
Il a été suggéré que de nombreuses protéines de capside virales ont pu évoluer à plusieurs reprises, à partir de protéines cellulaires fonctionnellement diverses[80].
Le recrutement de protéines cellulaires semble s'être produit à différents stades d'évolution ; certaines protéines cellulaires ont été capturées et fonctionnalisées avant la divergence des organismes cellulaires dans les trois domaines de la vie contemporains, tandis que d'autres n'ont été détournées que relativement récemment. En conséquence, certaines protéines de capside sont répandues dans les virus infectant des organismes apparentés éloignés (par exemple, les protéines de capside avec le repliement jelly roll), tandis que d'autres sont limitées à un groupe particulier de virus (par exemple, les protéines de capside des alphavirus)[80]
Cette ubiquité montre que les capsides ayant conduit aux virus sont antérieurs à LUCA, et peut-être même antérieurs à l'apparition des premières cellules autonomes.
Il est possible que ces capsides aient été un moyen pour les systèmes répliquant primitifs de réaliser des transferts de gènes horizontaux, permettant la survie de ces gènes hors de sites contaminés par des réplicateurs parasites ou menacés par un stress quelconque[57]. Ces échanges auraient alors été l'équivalent pré-cellulaire de la conjugaison bactérienne.
Maîtrise de la diffusion
La modélisation montre que dans une communauté de sites pré-biotiques contenant des réplicateurs, le métabolisme d'un site particulier isolé finit par s’effondrer à la suite de l'émergence de « parasites de réplication », qui consomment et assèchent les ressources disponibles du site sans contribuer au renouvellement de ces ressources[57], sciant la branche dont ils sont issus. Cependant, ce résultat n'est plus vrai quand les sites peuvent essaimer d'un site à l'autre. Dans ce cas, si la fréquence d'essaimage est comprise dans une certaine fourchette, le métabolisme d'un site a statistiquement le temps d'essaimer avant d'être submergé par l'émergence de parasites[57].
Cette fourchette temporelle dépend de deux temps moyens caractérisant l'implosion du métabolisme par parasitage : il faut en effet que le temps moyen entre deux essaimages soit suffisamment long, par rapport au temps d'implosion, pour que la probabilité d'un essaimage contaminé soit faible ; inversement ce temps moyen entre deux essaimages doit être suffisamment court, par rapport au délai moyen d'apparition d'un parasite, pour que le site pré-biotique ait eu le temps d'émettre un nombre suffisant d'essaims.
L'émergence de « parasites de réplication » dans des sites pré-biotiques étant probable, l'émergence d'un mécanisme de type capside, permettant de réaliser et réguler des transferts de gènes horizontaux, procure au site qui en dispose un avantage sélectif certain, et peut avoir été une condition nécessaire à la survie de la vie. De ce mécanisme d'échange primitif ont pu dériver d'une part les capsides virales, et d'autre part les mécanismes de conjugaison bactériennes[57].
De l'ARN à l'ADN
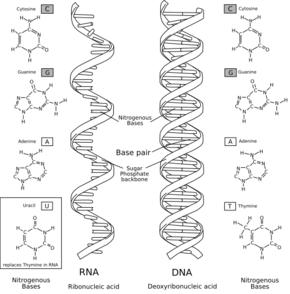
Ribozymes ou protéines ?
Bien que l'ARN soit donc à l'origine de l'ADN dans le métabolisme cellulaire, cette réaction est très difficile à réaliser. De fait, dans les trois lignées, elle est catalysée par des protéines spécialisées : les ribonucléotides-réductases. De plus, cette réaction est très coûteuse en énergie, du fait de la réduction du ribose, et elle produit des radicaux libres, très réactifs, sur la protéine. L'ARN étant une molécule fragile, il paraît improbable qu'elle puisse supporter des radicaux libres sans l'intervention de protéines.
Ainsi, l'origine de l'ADN trouve vraisemblablement sa source après l'apparition des protéines, indispensables à chaque étape de sa synthèse à partir de précurseurs de type ARN, au sein de la cellule.
Intérêt de l'ADN

L'ADN présente un certain nombre d'avantages sur l'ARN, en termes de conservation de l'intégrité de l'information génétique.
Tout d'abord il se casse moins facilement, car le désoxyribose de l'ADN contient un atome d'oxygène de moins que le ribose de l'ARN. Or l'oxygène peut facilement interagir sur les liaisons entre nucléotides, posant alors un problème de stabilité.
Certains auteurs avancent l'hypothèse que la désamination spontanée de la cytosine en uracile, aisément détectable dans l'ADN (où U n'est pas normalement présent) par la machinerie cellulaire de réparation des mutations, expliquerait l'utilisation de la base T dans l'ADN. L'ARN (notamment l'ARNm), molécule régulièrement renouvelée dans la cellule, ne voit pas sa séquence contrôlée par des systèmes de réparation, d'où une conservation de la base U dans cette molécule. Dans cette hypothèse, la base U est donc ancestrale, la base T dérivée.
Avantage sélectif de l'ADN : l'hypothèse du virus
Les avantages en termes de stabilité de l'ADN pourraient ne pas suffire à expliquer son adoption. Ainsi, Patrick Forterre avance l'hypothèse qu'un avantage sélectif supplémentaire peut être dû aux conflits entre virus et cellules vivantes.
Dans ce modèle, le premier organisme à ADN serait un virus. L'ADN conférerait au virus le pouvoir de résister à des enzymes dégradant les génomes à ARN, arme de défense probable des cellules. On retrouve le même principe chez des virus actuels, qui altèrent leur ADN pour résister à des enzymes produites par des bactéries infectées.
Actuellement, on peut observer que les enzymes nécessaires à la rétrotranscription de l'ARN vers l'ADN sont très présentes chez les rétrovirus, dont le génome est porté par de l'ARN. De la même façon, de nombreux virus codent leurs propres enzymes de synthèse de l'ADN.
Cette hypothèse est également corroborée par la découverte de virus à ADN, dont celui-ci contient, non pas des groupements thymine, mais des groupements uracile. Du point de vue évolutif, il y aurait donc eu d'abord apparition des désoxyribonucléotides, puis de l'ADN à uracile (ADN-U), puis d'ADN à thymine (ADN-T), qui se serait progressivement imposé. D'après Patrick Forterre, il est même probable que l'ADN-T ait été « inventé deux fois », chez des virus différents.
Les virus à ARN seraient ici des reliques du monde à ARN, les virus à ADN-U seraient alors des reliques du monde ayant précédé celui à ADN-T.
Les virus, premiers organismes à ADN

Les virus à ADN pourraient être plus anciens que la première cellule à ADN : la première cellule à ADN l'aurait donc emprunté à un ou plusieurs virus, sous la pression d'une course aux armements (théorie de la reine rouge).
Didier Raoult et Jean-Michel Claverie ont ainsi découvert le mimivirus : un virus géant à ADN (son génome étant deux fois plus long que le plus petit génome bactérien connu). La particularité de ce virus est qu'il peut produire des protéines impliquées dans la traduction de l'ARN en protéines (comme des enzymes chargeant des acides aminés sur des ARNt), il pourrait donc avoir pour ancêtres des virus plus anciens que la première cellule à ADN.
Eugène Koonin et ses collègues ont mis en avant, en comparant des génomes séquencés, que la plupart des enzymes impliqués dans la réplication de l'ADN sont différents entre les eubactéries et les eucaryotes (accompagnés des archées). Ils en concluent que l'ADN aurait été inventé indépendamment dans la lignée des eubactéries et celle conduisant aux eucaryotes et aux archées.
De même, les enzymes de réplications des virus à ADN sont très différentes d'un virus à l'autre, ainsi que par rapport aux enzymes cellulaires jouant le même rôle.
Ces indices laissent penser que les enzymes liées à l'ADN sont apparues au cours d'un « premier âge » du monde à ADN, où existaient cellules à ARN et virus à ARN et à ADN.
Passage de l'ADN dans les cellules
La nature du génome du plus ancien ancêtre commun à tous les êtres vivants (que les scientifiques prénomment LUCA) reste inconnue : faisait-il encore partie du monde à ARN, ou avait-il déjà un génome à ADN ? Quoi qu'il en soit, LUCA est le fruit d'une longue évolution. Le génome des premières cellules, qui ont précédé LUCA, était sans doute constitué par des molécules d'ARN et non pas d'ADN. Contrairement à l'ADN, l'ARN peut en effet jouer à la fois le rôle d'enzyme et de matériel génétique. Stanley Miller et Christian de Duve pensent que l'apparition de l'ARN a été elle-même un évènement tardif, en effet, cette molécule ne semble pas pouvoir être synthétisée par les méthodes simples de la chimie prébiotique. L'ARN aurait donc été précédé par des molécules dont nous ne connaîtrons sans doute jamais la nature exacte.
Ce qui amène les scientifiques à penser que l'ARN a précédé l'ADN résulte de ce constat : les trois grandes lignées du vivant ne partagent que le système de biosynthèse des protéines, alors qu'elles diffèrent sur le système de réplication de l'ADN[81].
En ce qui concerne la réplication de l'ADN, les cellules vivantes présentent deux jeux de protéines non homologues pour assurer cette réplication[82], l'un présent dans les bactéries, et l'autre trouvé dans les archées et les eucaryotes. En revanche, les protéines impliquées dans le métabolisme de l'ADN sont homologues dans les trois domaines. Ces éléments suggèrent que l'ADN a été inventé avant LUCA, mais n'était pas répliqué à cette époque ; les mécanismes de réplication n'ayant été inventés que postérieurement à la séparation archées / bactéries[83].
Il existe alors plusieurs hypothèses pour le passage de l'ADN (d'origine viral) dans les cellules vivantes : soit ce passage s'est produit une seule fois, soit il a pu avoir lieu plusieurs fois, indépendamment dans les différentes lignées. Dans le premier cas, les premiers gènes des enzymes de réplications auraient donc été remplacés par la suite par ceux d'un autre virus, formant ainsi les trois lignées.
L'hypothèse de Patrick Forterre est que les trois lignées du vivant trouvent leurs sources dans le remplacement du génome à ARN par le génome à ADN de trois virus différents. On retrouve notamment ici l'hypothèse d'une origine virale du noyau des eucaryotes.
Les travaux de Carl Woese semblent appuyer cette hypothèse, en démontrant que la vitesse d'évolution des protéines semble avoir chuté au moment de l'apparition des trois lignées. Cette diminution serait due au passage de l'ARN à l'ADN, les génomes à ADN étant plus stables, et donc moins sensibles aux mutations.
Dans ce scénario, l'apparition de trois lignées uniquement s'explique par le fait que les cellules à ADN ont peu à peu supplanté les cellules à ARN, empêchant ainsi l'apparition de nouvelles lignées par passage ARN→ADN.
Évolutions cellulaires
Racine de l'arbre de la vie
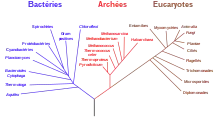
La comparaison de l'ARN des ribosomes montre l'émergence de trois formes de cellules : les bactéries, les archées et les eucaryotes[56],[79]. Cependant, il y a de bonnes raisons de croire que cette racine commune aux trois formes de cellules n'était pas une cellule au sens moderne du terme ; bien le code génétique se soit probablement formé à l'époque de cette racine, les mécanismes de la transcription devaient être rudimentaires et ont fait l'objet d'évolutions majeures par la suite[77].
Eucaryotes modernes
Les eucaryotes modernes présentent de nombreuses propriétés intermédiaires entre archées et bactéries.
Le génome des eucaryotes modernes est un hybride comprenant pour un tiers des gènes provenant des archées, concernant majoritairement le code génétique, et pour deux-tiers des gènes d'origine bactérienne[56].
L'explication généralement admise est que la cellule eucaryote moderne correspond à une symbiose entre un hôte archée et une protéobactérie alpha, qui sous forme symbiotique a évolué pour finalement constituer les mitochondries[56].
Autres modèles
Controverse sur la véracité de la théorie
On ne peut pas remonter le temps pour aller voir comment était l’océan primitif et comment il a évolué. Mais on peut trouver des témoignages indirects. Le passé laisse des traces dans le présent. Si on a les bons outils, théoriques et observationnels, on peut déduire le passé à partir du présent. Par exemple, les techniques autocatalytiques d’aujourd’hui se sont en quelque sorte fossilisées depuis des milliards d’années, puisqu’elles n’ont pas ou peu évolué. Elles nous renseignent donc sur un passé très lointain. En combinant ces informations avec d’autres, on peut espérer remonter encore plus loin dans le temps. Des expériences en laboratoire de chimie prébiotique peuvent apporter des renseignements précieux.
La théorie de la génération spontanée peut être combinée avec celle de la panspermie afin d'élargir le volume de matières mis en jeu à d'autres planètes viables. Néanmoins, elle rajoute la question de la probabilité que des organismes vivants puissent être véhiculés par des astéroïdes d'une planète viable à une autre au hasard des collisions intersidérales.
Milieu de collision entre planètes et noyaux de comètes
Selon des travaux de modélisation présentés en 2010[84], la compression brutale des matériaux constituant le noyau de glace d'une comète, au moment du choc avec une planète, pourrait donner lieu à la production d'acides aminés. Cette modélisation a pris comme base 210 molécules diluées dans un mélange d'eau, de méthanol, d'ammoniac, de dioxyde de carbone et de monoxyde de carbone, des composés qu'on estime être présents couramment dans le noyau des comètes. Avec une vitesse d'impact de 29 km/s, la pression atteint 10 gigapascals et une température de 700 K. Selon les auteurs, d'autres simulations, à pressions et températures plus élevées encore, conduisent à des réactions chimiques encore plus complexes. Selon le modèle, une simulation de 47 gigapascals et une température de 3 141 K pour les 20 premières picosecondes de l'impact fait naître des molécules complexes, dont de grosses molécules à liaisons carbone-azote qui pourraient être les premières briques de la vie. Le calcul a nécessité quelques milliers d'heures sur le superordinateur du cluster Atlas du Lawrence Livermore.
Sources hydrothermales : le monde du soufre et de l'azote
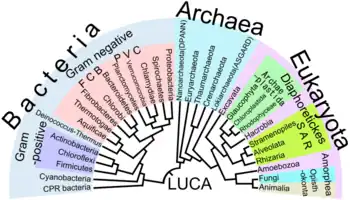
Des sources hydrothermales ont été découvertes en 1977 à 2 600 mètres de profondeur, là où deux plaques tectoniques se séparent.
Les monts hydrothermaux sont situés sur la couche sédimentaire. Leur diamètre à la base varie de 25 à 100 mètres et leur hauteur varie de 70 à 100 mètres. Les cheminées de ces fumeurs sont parfois recouvertes d'une croûte d'oxyde de manganèse. Les fumeurs situés sur ces monts sont composés d'un solide friable dont la couleur varie du gris noir à l'ocre, ce sont des sulfures de fer, de cuivre et de zinc.
Ces sources sont particulièrement intéressantes car on y a trouvé la vie où on la croyait impossible : milieu privé d'oxygène, à haute température, chargé de métaux et de soufre, dans l'obscurité la plus totale. Cependant les gradients de température importants autour de ces zones et le fait que les ultraviolets destructeurs ne parviennent pas si profondément (alors qu'ils détruisent toute molécule formée à la surface) sont de bonnes conditions pour l'apparition de la vie.
Ces organismes au mode de vie extrêmophile (forte température, forte acidité, niveaux de radiation extrême) ont les mêmes formes que ceux que l'on connaît plus près de la surface (ADN, protéines, sucres, etc.) mais puisent leur énergie de l'oxydation du sulfure d'hydrogène (H2S) pour transformer le carbone minéral en matière organique. Il s'agit d'archéobactéries hyperthermophiles (90-110 °C). Ce mode de vie correspondrait alors aux conditions de milieu qui régnaient sur Terre il y a 3,5 à 3,8 milliards d'années[86].
D'autre part, des expériences ont été menées[87], au laboratoire de géophysique de la Carnegie Institution de Washington, et ont montré que dans les conditions qui existent autour des évents, il y a formation d'ammoniac (NH3), forme réduite de l'azote qui est tant nécessaire à la formation des molécules organiques de la première partie et qui n'existait pas dans l'atmosphère oxydante. Les sources hydrothermales sont donc de bonnes sources de NH3.
Dans le cadre de l'International Ocean Discovery Program (en), les géomicrobiologistes et géochimistes ont pu observer des acides aminés aromatiques, molécules complexes indispensables au vivant, synthétisés abiotiquement par des réactions de Friedel-Crafts catalysées par une stéatite (argile riche en fer) durant l'altération de serpentinites du massif Atlantis au niveau de la dorsale médio-atlantique (serpentinisation des péridotites et stéatisation qui ne sont qu'une des facettes d'un phénomène plus général, l'hydrothermalisme du site de Lost City (en) associé aux roches du manteau)[88]. « C’est l’interaction entre l’eau de mer et les minéraux de ces roches, issues du manteau terrestre, ainsi que la structure en feuillet de l’argile résultant de leur altération, qui ont sans doute apporté les conditions idéales pour la formation de ces constituants primaires des premières briques du vivant, tel un miroir géologique aux expériences atmosphériques de Miller[89] ».
Une origine extraterrestre primitive (exogenèse)
Une hypothèse alternative est que la vie se soit d'abord formée hors de la Terre.
Les composés organiques sont relativement fréquents dans l'espace, notamment dans les zones lointaines du Système solaire où l'évaporation des composés volatils est très réduite. Certaines comètes présentent des couches externes de matière sombre, qu'on pense être une sorte de bitume formé par une combinaison de composés carbonés simples exposés aux rayons ultraviolets. La pluie de matériaux cométaires sur la Terre primitive pourrait avoir apporté des quantités de molécules organiques complexes, ce qui aurait favorisé l'apparition de la vie sur Terre.
Ce serait plutôt là un apport de matériaux organiques, plus ou moins complexes, que l'apport de la vie elle-même.
Panspermie
Une hypothèse plus large est la panspermie, une théorie selon laquelle l'origine de la vie sur Terre serait due à une contamination extraterrestre. Des transferts d'organismes vivants microscopiques, à travers l'espace jusqu'à la Terre, naturels ou délibérés, grâce aux comètes ou aux météorites par exemple[90]. Certains pensent aussi que la vie est originaire de l'espace, les théories sont nombreuses.
En cas de contamination volontaire, on parle de panspermie dirigée.
Ces théories d'une origine extraterrestre n'expliquent pas directement comment la vie est apparue, car elles ne font a priori que reporter le problème. Cependant, elles élargissent les conditions dans lesquelles la vie a pu apparaître dans l'Univers. Les futurs échantillons de sols ramenés de Mars et de comètes permettront peut-être d'obtenir de nouveaux éléments de réponse.
Origine de l’évolution darwinienne plutôt qu’origine de la vie
Des scientifiques préfèrent étudier la question de l'origine de la vie à partir d’un paradigme ou d’un point de vue différent. Il s’agit de traiter la question de comment et quand l’évolution a commencé au lieu d’essayer de définir la vie et de délimiter une frontière quant à son origine[91],[92].
Aujourd’hui il semble y avoir un consensus parmi les chercheurs sur l’origine de la vie (chimistes, géochimistes, biochimistes, exo/astrobiologistes, informaticiens, philosophes et historiens des sciences) qu’il est absolument nécessaire de trouver une définition universelle de la vie[93]. Cependant le nombre de définitions ne cesse d’augmenter[94]. De fait, la frontière entre systèmes vivants et systèmes non vivants de même que le moment à partir duquel le non vivant serait devenu vivant sont arbitraires.
On constate depuis quelques années un changement profond car le problème de la vie n’est plus de rechercher les principes qui la sous-tendent mais est devenu un problème de nature historique. La question n’est plus : « Qu’est-ce qui caractérise les organismes par rapport aux objets inanimés ? » ; mais plutôt : « comment ces caractéristiques se sont progressivement installées dans les systèmes que nous appelons des organismes ? »[95]. Finalement, la question deviendrait : « Comment l'évolution darwinienne a-t-elle émergé sur la Terre, il y a quatre milliards d'années environ, dans un monde qui ne la contenait pas encore ? »[96].
Notes et références
- 1 2 3 (en) Elizabeth A. Bella, Patrick Boehnkea, T. Mark Harrisona et Wendy L. Maob, « Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon », Proceedings of the National Academy of Sciences, (DOI 10.1073/pnas.1517557112).
- 1 2 3 4 (en) Matthew S. Dodd, Dominic Papineau, Tor Grenne, John F. Slack, Martin Rittner, Franco Pirajno, Jonathan O’Neil et Crispin T. S. (2017), Little Evidence for early life in Earth’s oldest hydrothermal vent precipitates, Nature, 543, p. 60–64, 2 mars 2017, DOI 10.1038/nature21377, résumé.
- ↑ J. William Schopf, Anatoliy B. Kudryavtsev, David G. Agresti, Thomas J. Wdowiak et Andrew D. Czaja, « Laser–Raman imagery of Earth's earliest fossils », Springer Science and Business Media LLC, vol. 416, no 6876, , p. 73–76 (ISSN 0028-0836, DOI 10.1038/416073a)
- ↑ J. William Schopf, Anatoliy B. Kudryavtsev, Andrew D. Czaja et Abhishek B. Tripathi, « Evidence of Archean life: Stromatolites and microfossils », Elsevier BV, vol. 158, nos 3-4, , p. 141–155 (ISSN 0301-9268, DOI 10.1016/j.precamres.2007.04.009)
- ↑ (en) John Charles Priscu, « Origin and Evolution of Life on a Frozen Earth » [archive du ], Arlington County, VA, National Science Foundation (consulté le ).
- ↑ (en) Norman H. Sleep, The Hadean-Archaean Environment, Cold Spring Harbor perspectives in biology, juin 2010.
- ↑ (en) Van Kranendonk, M. J., Philippot, P., Lepot, K., Bodorkos, S. et Pirajno, F. (2008), Geological setting of Earth’s oldest fossils in the c. 3.5 Ga Dresser Formation, Pilbara craton, Western Australia. Precambr. Res., 167, 93–124.
- ↑ Abderrazak El Albani et Alain Meunier, Aux origines de la vie, Dunod, 2016.
- ↑ (en) J. Alleon, S. Bernard, C. Le Guillou, O. Beyssac, K. Sugitani et F. Robert, « Chemical nature of the 3.4 Ga Strelley Pool microfossils », Geochemical Perspectives Letters, vol. 7, , p. 37-42 (DOI 10.7185/geochemlet.1817).
- 1 2 (en) Allen P. Nutman, Vickie C. Bennett, Clark R. L. Friend, Martin J. Van Kranendonk et Allan R. Chivas, « Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures », Nature, vol. 537, , pages 535-538 (DOI 10.1038/nature19355).
- 1 2 (en) Mark A. van Zuilen, « Proposed early signs of life not set in stone », Nature, vol. 563, , p. 190-191 (DOI 10.1038/d41586-018-06994-x).
- 1 2 (en) Abigail C. Allwood, Minik T. Rosing, David T. Flannery, Joel A. Hurowitz et Christopher M. Heirwegh, « Reassessing evidence of life in 3,700-million-year-old rocks of Greenland », Nature, vol. 563, , p. 241-244 (DOI 10.1038/s41586-018-0610-4).
- ↑ Des microfossiles vieux de 3,77 milliards d’années découverts au Canada, sur lemonde.fr.
- ↑ Rosing, M. T. (1999), « 13C-Depleted carbon microparticles in >3700-Ma sea-floor sedimentary rocks from west greenland », Science, 283, 674–676 CAS.
- 1 2 (en) Yoko Ohtomo, Takeshi Kakegawa, Akizumi Ishida, Toshiro Nagase et Minik T. Rosing, « Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks », Nature Geoscience, vol. 7, , p. 25-28.
- ↑ Bell, E. A., Boehnke, P., Harrison, T. M. et Mao, W. L. (2015), Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon, Proc. Natl. Acad. Sci. USA, 112, 14518–14521.
- ↑ Pinti, D. L., Hashizume, K. et Matsuda, J. (2001), Nitrogen and argon signatures in 3.8 to 2.8 Ga metasediments: clues on the chemical state of the Archean ocean and the deep biosphere, Geochim. Cosmochim. Acta, 65, 2301–2315.
- ↑ van Zuilen, M.A. et al. (2005), Nitrogen and argon isotopic signatures in graphite from the 3.8-Ga-old Isua Supracrustal Belt, Southern West Greenland, Geochim. Cosmochim. Acta, 69, 1241–1252.
- ↑ Hassenkam T., Andersson M.P., Dalby K.N., Mackenzie D.M.A. et Rosing M.T. (2017), Elements of Eoarchean life trapped in mineral inclusions, Nature, DOI 10.1038/nature23261, 24 juillet 2017, résumé.
- ↑ (en) Robert Shapiro, Origins : A Skeptic's Guide to the Creation of Life on Earth, Bantam Books, , 332 p. (ISBN 0-671-45939-2), p. 110.
- ↑ Oparin A, Fesenkov V., Life in the Universe, Moscou : USSR Academy of Sciences publisher, 3rd edition, 1956 (in Russian). English translation: Oparin, A. et V. Fesenkov. Life in the Universe. New York: Twayne Publishers (1961).
- ↑ Bernal, J.D. (1959) The problem of stages in biopoiesis In The Origin of Life on the Earth, edited by A.I. Oparin, Pergamon Press, London, p. 38–43.
- ↑ Varela FG, Maturanaa HR, Uribea R. (May 1974) Autopoiesis: The organization of living systems, its characterization and a model, Biosystems, 5 (4):187-196.
- ↑ Grégoire Nicolis et Ilya Prigogine, Exploring complexity: An introduction, New York, NY: W. H. Freeman, 1989. (ISBN 0716718596).
- ↑ Tirard S, Morange M et Lazcano A. (2010), The Definition of Life: A Brief History of an Elusive Scientific Endeavor, Astrobiology, 10, 1003-1009.
- ↑ Orgel, L.E. (2000) Self-organizing biochemical cycles, Proc Natl Acad Sci USA, 97, 12503–12507.
- ↑ Benner et al., dans Gesteland RF, Cech TR, Atkins JF. (2006), The RNA World, 3e éd., Cold Spring Harbor Laboratory Press, New York, NY, États-Unis, 2006.
- 1 2 3 4 The ribosome as a missing link in the evolution of life. Meredith Root-Bernstein, Robert Root-Bernstein. Journal of Theoretical Biology, Volume 367, 21 February 2015, Pages 130-158.
- 1 2 3 4 5 Richard Dawking, The ancestor's tale, Chap. Canterbury.
- ↑ (en) J. W. Szostak, « An optimal degree of physical and chemical heterogeneity for the origin of life? », Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 366, no 1580, , p. 2894–2901 (DOI 10.1098/rstb.2011.0140).
- 1 2 3 Voir Prevolutionary dynamics and the origin of evolution, Martin Nowak, Hisashi Ohtsuki, Proceedings of the National Academy of Science of the USA, 2008.
- 1 2 3 4 5 6 The Selfish Gene, Richard Dawkins, 1976.
- ↑ La profusion des aléas et des redondances au sein du vivant est une cause majeure d'inefficience locale qui est un facteur d'adaptabilité et de résilience. Voir Olivier Hamant, La Troisième Voie du vivant, Odile Jacob, , p. 119.
- 1 2 3 4 5 6 Escapement mechanisms and the conversion of disequilibria; the engines of creation. E. Branscomb, T. Biancalani, N. Goldenfeld, M. Russell, Physics Reports, volume 677, 2017, p. 1-60, ISSN 0370-1573.
- ↑ L. Boltzmann, « La deuxième loi de la thermodynamique » (discours à une réunion formelle de l'Académie impériale des sciences, 29 mai 1886, réimprimé dans « Physique théorique et problème philosophique ».
- ↑ Modern Thermodynamics from Heat Engines to Dissipative Structures. Kondepudi & Prigogine. John Wiley & Sons, 1998. (ISBN 9780471973942).
- 1 2 3 The inevitable journey to being.Russell MJ, Nitschke W, Branscomb E. 2013. Phil Trans R Soc B 368: 20120254.
- 1 2 3 4 The inevitable journey to being. Michael J. Russell, Wolfgang Nitschke, Elbert Branscomb. Phil Trans R Soc B 368: 20120254.
- ↑ On peut remarquer que le cycle est fonctionnellement invariant à une inversion de sens de parcours : dans un milieu où au contraire la réaction D⇒C serait motrice et la réaction B⇒A entraînée, le même médiateur M pourrait coupler ces deux réactions en parcourant le cycle en sens inverse.
- 1 2 3 Turnstiles and bifurcators: The disequilibrium converting engines that put metabolism on the road. Elbert Branscomb, Michael J.Russell. Biochimica et Biophysica Acta (BBA) - Bioenergetics. Volume 1827, Issue 2, February 2013, Pages 62-78.
- ↑ (en) Ward, Peter et Kirschvink, Joe, A New History of Life : the radical discoveries about the origins and evolution of life on earth, Bloomsbury Press, , 39–40 p.
- ↑ François Robert et Marc Chaussidon, A palaeotemperature curve for the Precambrian oceans based on silicon isotopes in cherts, Nature, 443, 969-972, 26 octobre 2006.
- ↑ Matthew W. Powner, Béatrice Gerland et John D. Sutherland, Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions, Nature, 2009, 459, 239-242.
- ↑ Pour la Science, décembre 2019, p. 12
- ↑ (en) S. Becker et al., Unified prebiotically plausible synthesis of pyrimidine and purine RNA ribonucleotides, Science, 2019 DOI 10.1126/science.aax2747
- ↑ (en) Denis Comte, Léo Lavy, Paul Bertier, Florent Calvo, Isabelle Daniel et al., « Glycine Peptide Chain Formation in the Gas Phase via Unimolecular Reactions », Journal of Physical Chemistry A, vol. 127, no 3, , p. 775-780 (DOI 10.1021/acs.jpca.2c08248).
- ↑ Baross J, Hoffman SE, Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life, Orig. Life Evol. Biosph., 1985, 15, 327-345.
- ↑ Rushdi AI, Simoneit BR. Lipid formation by aqueous Fischer-Tropsch-type synthesis over a temperature range of 100 to 400 degrees C. Orig.Life Evol.Biosph. 2001, 31, 103-118.
- ↑ (en) G. Proskurowski, « Abiogenic Hydrocarbon Production at Lost City Hydrothermal Field », Science, vol. 319, no 5863, , p. 604–607 (DOI 10.1126/science.1151194).
- ↑ (en) William Martin, « Hydrothermal vents and the origin of life », Nature Reviews Microbiology, (DOI 10.1038/nrmicro1991).
- ↑ (en) Graham Cairns-Smith, Seven clues to the origin of life, 1985 ; édition française : L'énigme de la vie, 1990.
- ↑ (en) Yannick Vallee, Ibrahim Shalayel, Kieu-Dung Ly et K. V. Raghavendra Rao, « At the very beginning of life on Earth: the thiol-rich peptide (TRP) world hypothesis », International Journal of Developmental Biology, vol. 61, nos 8-9, (ISSN 0214-6282 et 1696-3547, lire en ligne, consulté le ).
- ↑ (en) Robert F. Service, « Did volcanic ‘glasses’ help spark early life? », Science, (DOI 10.1126/science.add3257, lire en ligne
 , consulté le ).
, consulté le ). - ↑ (en) Craig A. Jerome, Hyo-Joong Kim, Stephen J. Mojzsis, Steven A. Benner et Elisa Biondi, « Catalytic Synthesis of Polyribonucleic Acid on Prebiotic Rock Glasses », Astrobiology (en), (DOI 10.1089/ast.2022.0027, lire en ligne , consulté le ).
- ↑ Cité d'après Isaac Asimov, Asimov's new guide to science (1984), chap. 12.
- 1 2 3 4 5 6 7 (en) Eugene V Koonin, Carl Woese’s vision of cellular evolution and the domains of life, RNA Biology, 11:3, p. 197–204, mars 2014.
- 1 2 3 4 5 Jalasvuori M, Mattila S et Hoikkala V, Chasing the Origin of Viruses: Capsid-Forming Genes as a Life-Saving Preadaptation within a Community of Early Replicators, PLOS One, 10(5), e0126094, DOI 10.1371/journal.pone.0126094, 8 mai 2015
- ↑ Jack W. Szostak, David P. Bartel et P. Luigi Luisi, Synthesizing life, Nature, vol. 409, p. 387–390 (2001).
- 1 2 Anne-Sophie Boutaud, Quand la Terre était une boule de neige, Le journal du CNRS, 5 décembre 2017
- 1 2 3 Peter Schönheit, Wolfgang Buckel et William F. Martin, On the Origin of Heterotrophy, Trends in Microbiology, janvier 2016, vol. 24, n° 1.
- ↑ Wolfgang Nitschke, The evolutionary aspects of bioenergetic systems, Biochimica et Biophysica Acta (BBA), Bioenergetics, vol. 1827, n° 2, février 2013, p. 61.
- 1 2 3 4 Why Are Cells Powered by Proton Gradients? Nick Lane, Nature Education 3(9):18 2010.
- ↑ Petsch, S. T., The Global Oxygen Cycle, Treatise on Geochemistry, vol. 8, éditeur William H. Schlesinger, éditeurs exécutifs Heinrich D. Holland et Karl K. Turekian, p. 682 (ISBN 0-08-043751-6), Elsevier, 2003, p. 515-555.
- 1 2 A Coupled Ecosystem-Climate Model for Predicting the Methane Concentration in the Archean Atmosphere. James F. Kasting, Alexander A. Pavlov, Janet L. Siefert. Origins of life and evolution of the biosphere, juin 2001, vol. 31, n° 3, p. 271–285.
- ↑ François Roddier, Vie et dissipation d’énergie, « point de vue d'un astronome », 2 janvier 2007.
- ↑ Review : Re-creating an RNA world, U. F. Müller, Cellular and Molecular Life Sciences, 63 (2006) 1278–1293.
- 1 2 3 (en) Marcello Barbieri Code Biology – A New Science of Life, Biosemiotics, volume 5, numéro 3, décembre 2012.
- ↑ Self replicating systems. Patzke V, von Kiedrowski G, ARKIVOC 2007, 8:293–310.
- ↑ (en) C. Woese, G. Fox, « The concept of cellular evolution », J. Mol. Evol., (PMID 903983).
- ↑ (en) C. Woese, G. Fox, « Phylogenetic structure of the prokaryotic domain: the primary kingdoms », Proc. Natl. Acad. Sci. USA, vol. 74, no 11, , p. 5088–90 (PMID 270744, DOI 10.1073/pnas.74.11.5088).
- 1 2 3 Doron Lancet, Daniel Segrè et Amit Kahana, Twenty Years of « Lipid World »: A Fertile Partnership with David Deamer, Life, 9(4), 2019, p. 77, DOI 10.3390/life9040077.
- 1 2 Pré-vivants : les composomes vivent la vie d'avant la vie, Science et Vie, no 1218, « Les nouvelles frontières de la vie ».
- ↑ Harvey Lodish, Arnold Berk, Paul Matsudaira, Chris-A Kaiser, M. Krieger, M.P. Scott, Lodish Zipursky et James Darnell, Biologie moléculaire de la cellule, Bruxelles, de Boeck, , 3e éd., 1096 p. (ISBN 978-2-8041-4802-7).
- ↑ (en) Nicholas V. Hud et David M. Fialho, « RNA nucleosides built in one prebiotic pot », Science, vol. 366, no 6461, , p. 32-33 (DOI 10.1126/science.aaz1130).
- ↑ (en) Sidney Becker, Jonas Feldmann, Stefan Wiedemann, Hidenori Okamura, Christina Schneider et al., « Unified prebiotically plausible synthesis of pyrimidine and purine RNA ribonucleotides », Science, vol. 366, no 6461, , p. 76-82 (DOI 10.1126/science.aax2747).
- ↑ (en) M Sumper et R. Luce, « Evidence for de novo production of self-replicating and environmentally adapted RNA structures by bacteriophage Qbeta replicase », Proceedings of the National Academy of Sciences, vol. 72, no 1, , p. 162–166 (PMID 1054493, PMCID 432262, DOI 10.1073/pnas.72.1.162).
- 1 2 3 4 5 6 7 Carl R. Woese, On the evolution of cells, Proc. Natl. Acad. Sci. USA, 25 juin 2002, 99 (13), 8742–8747.
- 1 2 L’origine de la vie et les métalloenzymes à fer-soufre. Juan C. Fontecilla-Camps. Exposé au Collège de France, 26 mai 2010.
- 1 2 Carl Woese, The universal ancestor, Proc. Natl. Acad. Sci. USA, vol. 95, p. 6854–6859, juin 1998.
- 1 2 (en) M Krupovic et EV Koonin, « Multiple origins of viral capsid proteins from cellular ancestors », Proc Natl Acad Sci U S A, vol. 114, no 12, , E2401–E2410 (PMID 28265094, PMCID 5373398, DOI 10.1073/pnas.1621061114)
- ↑ Marie-Christine Maurel, « À la frontière du vivant : les viroïdes », sur The Conversation (consulté le ).
- ↑ Patrick Forterre, The two ages of the RNA world, and the transition to the DNA world: a story of viruses and cells, Biochimie, vol. 87, n° 9–10, septembre–octobre 2005, p. 793-803.
- ↑ Patrick Forterre, The origin of viruses and their possible roles in major evolutionary transitions, Virus Research, vol. 117, n° 1, avril 2006, p. 5-16.
- ↑ Nir Goldman et al. (Lawrence Livermore National Laboratory), American Chemical Society, , San Francisco, Californie ; puis Comet crash creates potential for life, Nature, , DOI 10.1038/news.2010.152.
- ↑ Luciano Paolozzi et Jean-Claude Liébart, Microbiologie, Éditions Dunod, , p. 50.
- ↑ (en) Gregory Bock, Evolution of hydrothermal ecosystems on Earth (and Mars ?), Wiley, , p. 11.
- ↑ (en) Brandes et al., Abiotic nitrogen reduction on the early Earth, Nature, 1998.
- ↑ (en) Bénédicte Ménez, Céline Pisapia, Muriel Andreani, Frédéric Jamme, Quentin P. Vanbellingen, Alain Brunelle, Laurent Richard, Paul Dumas & Matthieu Réfrégiers, « Abiotic synthesis of amino acids in the recesses of the oceanic lithosphere », Nature, (DOI 10.1038/s41586-018-0684-z).
- ↑ « Les premières briques à l’origine de la vie sur Terre observées dans des roches océaniques profondes », sur ipgp.fr, .
- ↑ (en-US) Peter Reuell, « Harvard study suggests asteroids might play key role in spreading life », sur Harvard Gazette, (consulté le ).
- ↑ Cyrille Jeancolas, « Modéliser la transition de l’inerte au vivant. Ethnographie d’un laboratoire de recherche microfluidique », Techniques & Culture. Revue semestrielle d’anthropologie des techniques, (ISSN 0248-6016, lire en ligne, consulté le )
- ↑ Thomas Heams, Infravies - Le vivant sans frontières, Editions du Seuil, (ISBN 978-2-02-142447-8, lire en ligne)
- ↑ (en) Tsokolov SA., « Why is the definition of life so elusive? Epistemological considerations », Astrobiology 2009, 9, p. 401-412.
- ↑ Trifonov EN., Vocabulary of Definitions of Life Suggests a Definition, J. Biomol. Struct. Dyn., 2011, 29 (2), p. 259-266 (DOI 10.1080/073911011010524992).
- ↑ (en) S. Tirard S et al., The Definition of Life: A Brief History of an Elusive Scientific Endeavor, Astrobiology, 2010, 10, p. 1003-1009.
- ↑ (en) Tessera M., « Is A n+1 Definition of Life Useful? », J. Biomol. Struct. Dyn., 2012, 29, (4):635-636 (https://doi.org/10.1080/073911012010525012)
Voir aussi
Sources
- (en) John Desmond Bernal, The origin of life, éd. Weidenfeld & Nicolson, 1967.
- Guillemette Lauters, Olivier Lenaerts, Daniel Rousselet et Eric Depiereux, Bioscope : L’origine de la vie.
- John Maynard Smith et Eörs Szathmáry, Les Origines de la vie, éd. Dunod, 2000 (ISBN 2100048600)
- (en) J. William Schopf, Anatoliy B. Kudryavtsev, David G. Agresti, Thomas J. Wdowiak et Andrew D. Czaja, « Laser–Raman imagery of Earth's earliest fossils », Springer Science, vol. 416, no 6876, , p. 73–76 (ISSN 0028-0836, DOI 10.1038/416073a)
- (en) J. William Schopf, Anatoliy B. Kudryavtsev, Andrew D. Czaja et Abhishek B. Tripathi, « Evidence of Archean life: Stromatolites and microfossils », Elsevier BV, vol. 158, nos 3-4, , p. 141–155 (ISSN 0301-9268, DOI 10.1016/j.precamres.2007.04.009)
- Marie-Christine Maurel, La Naissance de la vie : de l'évolution prébiotique à l'évolution biologique, éd. Dunod, 2003, 3e éd. (ISBN 2100068822)
- (en) N. Fujii et T. Saito, Homochirality and life, The Chemical Records, vol. 4, no 5, p. 267–78, 2004 [(en) lire en ligne]
- Uwe Meierhenrich, « Un pas vers l’origine de l’homochiralité… »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ), .
- Patrick Forterre, « L'origine du génome », dans Les dossiers de La Recherche : l'histoire de la vie, les grandes étapes de l'évolution, no 19, p. 34–40, .
- Marc-André Selosse, « Quelle parenté entre les trois grandes lignées du vivant ? », dans Les dossiers de La Recherche : l'histoire de la vie, les grandes étapes de l'évolution, no 19, p. 42–45, .
- « Laurent Nahon, « L'asymétrie des biomolécules vient de l'espace » », propos recueillis par Franck Daninos, dans La Recherche, no 390, .
- (en) U. Meierhenrich, L. Nahon, C. Alcaraz, J. Bredehöft, S. Hoffmann, B. Barbier et A. Brack, « Asymmetric VUV photodecomposition of the amino acid D,L-Leucine in the solid state », dans Angewandte Chemie International Edition, vol. 44, p. 5630–5634, [(en) lire en ligne].
Bibliographie
- Éric Karsenti, Aux sources de la vie, Flammarion, , 306 p. (lire en ligne).
- Abderrazak El Albani et Alain Meunier, Aux origines de la vie, Dunod, 2016.
- (en) Uwe Meierhenrich, Amino acids and the asymmetry of life, Springer-Verlag, 2008 (ISBN 978-3-540-76885-2).
- (en) Site consacré au mimivirus.
- (en) Anthony M. Poole, « What is the Last Universal Common Ancestor ? »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ), .
- (en) Antonio Lazcano « The Origins of Life. Have too many cooks spoiled the prebiotic soup? » (consulté le ), Natural History magazine, .
- Review : (en) The origin of modern terrestrial life. Patrick Forterre, Simonetta Gribaldo. HFSP J., septembre 2007, 1(3): 156–168.
- (en) Erwin Schrödinger, What Is Life?, 1944 (traduction française : Qu'est-ce que la vie ?, éd. du Seuil).
- Joël de Rosnay, Les Origines de la vie, éd. Seuil, 1966 (ISBN 2020002167).
- Jean C. Baudet, La vie expliquée par la chimie, Vuibert, Paris, 2006.
- André Brack et Bénédicte Leclercq, La vie est-elle universelle ? – Des premiers êtres vivants à l'exploration spatiale, éd. EDP Sciences, 2003 (ISBN 2-86883-674-7).
- Sous la direction de Muriel Gargaud, Didier Despois, Jean-Paul Parisot, L'Environnement de la Terre Primitive, coll. « L'origine de la Vie sur Terre et dans l'Univers », Presses universitaires de Bordeaux, 2001 (ISBN 2-86781-267-4).
- (en) Jeffrey L. Bada, « How life began on Earth: a status report », Earth and Planetary Science Letters, vol. 226, nos 1-2, , p. 1-15 (DOI 10.1016/j.epsl.2004.07.036).
- Charles Darwin, L'Origine des espèces, édition du Bicentenaire, trad. A. Berra sous la dir. de P. Tort, coord. par M. Prum. Précédé de Patrick Tort, « Naître à vingt ans. Genèse et jeunesse de L’Origine », Paris, Champion Classiques, 2009.
- Guillaume Suing, Origine de la vie - Un siècle après Oparine, Delga, 2020.
Articles connexes
- Auto-organisation
- Biologie, Biologie cellulaire, Vie
- Biochimies hypothétiques
- Chauvinisme du carbone
- Exobiologie
- Fossiles les plus anciens
- Génération spontanée
- Habitabilité d'une planète
- Histoire évolutive du vivant
- Lyfe (astrobiologie)
- Protéinoïde
- Bombardement par rayonnement ionisant
- Origines de la vie multicellulaire
- Évolution de la multicellularité
- Vie sur d'autres planètes
- Hypothèse de la Terre pourpre
Liens externes
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Site de la Société française d'Exobiologie (SFE).
- Conférence de Patrick Forterre, Regards croisés sur les origines de la vie (2009) à l'« ENS Cachan »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ).
 Écouter La leçon consacrée aux origines chimiques de la vie et dispensée par Jacques Reisse au Collège Belgique (2009).
Écouter La leçon consacrée aux origines chimiques de la vie et dispensée par Jacques Reisse au Collège Belgique (2009).- Institut Jacques-Monod, « Les origines du vivant », sur Université Pierre-et-Marie-Curie - Paris VI (consulté en )
- Les premières traces de la vie sur Terre – L'univers de la géologie